



Publicado
Estabilidad fenotípica y densidades de planta de genotipos de estevia (Stevia rebaudiana Bert.) en la región Caribe de Colombia
Phenotypic stability and plant densities of stevia (Stevia rebaudiana Bert.) genotypes in the Caribbean Region of Colombia
Palabras clave:
Densidad de siembra, glucósidos, interacción genotipo-ambiente, relación hoja/tallo, rendimiento de biomasa (es)Biomass yield, genotype-environment interaction, glycosides, leaf/stem ratio, plant density. (en)
1Profesores Investigadores, Universidad de Córdoba, Montería, Colombia. 2Profesor Investigador, Universidad Pedagógica y Tecnológica de Colombia, Duitama, Colombia. *Autor para correspondencia: ajarma@sinu.unicordoba.edu.co; †haramendiz@hotmail.com; ‡clevesalejandro@yahoo.com
Los estudios de estabilidad fenotípica son importantes para la siembra de genotipos que muestran un adecuado comportamiento en una o varias zonas. Estevia (Stevia rebaudiana Bert.) es una especie recientemente introducida en Colombia, por tanto, los estudios sobre su adaptación en áreas productoras del país son escasos. Esta planta se caracteriza por sintetizar glucósidos de diterpeno altamente edulcorantes con bajo aporte calórico. El presente estudio se realizó en Montería (8° 52' N y 76° 48' O), Colombia, con el objeto de determinar la estabilidad fenotípica de los genotipos Morita 1 y Morita 2 de estevia, sembrados en tres densidades de plantas (25, 12.5 y 6.25 plantas/m2) y dispuestos en un diseño de bloques completos al azar, en tres épocas de siembra en el Caribe húmedo de Colombia. Los resultados más importantes señalan que Morita 2, en densidades de 25 plantas/m2, registró los mejores índices de estabilidad para el rendimiento de hoja seca/ha (Pig = 0; Pid = 0). Por su parte, éste mismo genotipo presentó una mayor relación hoja/tallo (1.17 ± 0.3) y mayor contenido de glucósidos totales (0.1586 ± 0.024) cuando la radiación solar fue mayor y las densidades eran entre 12.5 y 25 plantas/m2.
Palabra clave: Densidad de siembra, glucósidos, interacción genotipo-ambiente, relación hoja/tallo, rendimiento de biomasa.
Phenotypic stability studies are important to recommend objectively the planting of genotypes showing appropriate behavior in one or more areas. Stevia (Stevia rebaudiana Bert.), a species of recent cropping in Colombia, is characterized by the capacity of synthesize highly sweeteners diterpene glycosides with low calorie. However, while defining the environmental conditions for production, leaf /stem ratio in the canopy, and the amount of total glycosides, few studies of adaptation have been made in the country. This study was carried out in Montería-Colombia (8°52' N and 76°48' W) to determine the phenotypic stability of two genotypes of stevia (Morita 1 and Morita 2) and three plant densities (25; 12.5 and 6.25 plants/ m2) during three sowing seasons in the Humid Colombian Caribbean Region. The experimental design consisted of a randomized complete block. The results showed that Morita 2, established in densities of 25 plants/m2, recorded the highest levels of stability for the dry leaf yield per hectare (Pig =0; Pid =0). On the other hand, the same genotype showed a higher leaf/stem ratio (1.17±0.3) and higher content of total glycosides (0.1586±0.024) when radiation is higher and plant densities are between 12.5 and 25 plants/m2.
Key words: Biomass yield, genotype-environment interaction, glycosides, leaf/stem ratio, plant density.
Stevia rebaudiana Bert. es una planta herbácea perenne que pertenece a la familia de las Asteráceas. Crece como arbusto salvaje en el suroeste de Brasil y Paraguay y tiene un alto valor entre los vegetales nativos de ambos países debido a su contenido de glucósidos bajos en calorías, llamados comúnmente esteviósidos, cuyo poder edulcorante en estado puro y cristalino puede ser 300 veces mayor que el del azúcar de caña (Dacome et al., 2005).
Los edulcorantes en esta planta se encuentran concentrados en las hojas, son glucósidos de diterpeno sintetizados, al menos en los estados iniciales, usando la misma ruta del ácido giberélico a partir del mevalonato (Hsieh y Goodman, 2005; Guevara-García et al., 2005; Kasahara et al., 2002). Varios investigadores señalan que la diferencia radica que en estevia, el kaureno, precursor de dichas hormonas, se convierte a esteviol en el retículo endoplasmático (Geuns, 2003; Totté et al., 2000; Totté et al., 2003; Brandle et al., 2002; Brandle, 2005).
Con la variación en la época de siembra y la densidad de plantas, los factores ambientales como radiación solar, precipitación y temperatura pueden afectar las expresiones fenotípicas y fisiológicas de las plantas, ya que se afectan los procesos de fotosíntesis, transpiración, alargamiento celular, crecimiento y competencia entre especies (Jarma, 2008; Montemayor et al., 2006). En diez ambientes contrastantes de la India, Jaitak et al. (2008) encontraron que los edulcorantes de estevia presentaban valores entre 3.3% y 6.7% de esteviósidos (St) y entre 1.2% y 2.3% de rebaudiósido A (RebA), lo que confirmó el efecto del ambiente sobre la síntesis de glucósidos de esteviol. La diferencia entre estos glucósidos radica en que el RebA posee una molécula adicional de glucosa en su estructura.
Los cultivos de estevia en Colombia se han establecido sin criterios técnicos o científicos sobre la base genética de los cultivares (cv.), Morita 1 y Morita 2 (Jarma et al., 2005) de los cuales no se conoce su estabilidad fenotípica, lo que implica un alto riesgo derivado de su variabilidad genética estrecha que, asociada a la alta variabilidad ambiental del país, los hace sensibles a factores tanto bióticos como abióticos. En la actualidad no existen registros oficiales del área cultivada en estevia, aunque se ha reportado su siembra en los departamentos de Antioquia, Córdoba, Tolima, Huila, Valle del Cauca, Caquetá y Meta en ambientes que presentan condiciones agroecológicas variables, siendo el cv. Morita 2 el más cultivado, aunque autores como Maya (2000) estiman que el cv. Morita 1 es igualmente cultivado. El manejo agronómico de este cultivo, considerando labores como siembras, podas, controles fitosanitarios y beneficio, lo convierten en un sistema altamente generador de empleos rurales, que giran alrededor de 245 jornales/ha por año (Jarma, 2008). En Colombia, trabajos adelantados en la región Caribe han sugerido su posible adaptación a la oferta ambiental de esta parte del país, lo que podría convertirlo en una nueva alternativa de producción para el sector agroindustrial (Jarma et al., 2005; 2006). Es una especie que se reproduce sexual y asexualmente, pero este último método es el más utilizado en la producción de plántulas para el establecimiento de cultivos, siendo una práctica frecuente en diversos países como Colombia (Jarma, 2009), Brasil (Dacome et al., 2005; Yoda et al., 2003; Campos, 2007), Paraguay (Casaccia y álvarez, 2006), India (Jaitak et al., 2008) y Australia (Hearn y Subedi, 2008).
La estabilidad fenotípica de especies vegetales tiene varias interpretaciones. Becker (1981) propuso dos conceptos: el biológico, cuyo ideal es el genotipo de mínima variación a través de ambientes, que muestra un rendimiento constante en cualquier condición de producción (varianza estadística mínima), y el agronómico, que representa una mínima interacción genotipo x ambiente, lo cual está asociado con la pretensión de obtener un incremento del rendimiento en respuesta a mejoras ambientales.
La metodología de Lin y Binns (1988) es una buena alternativa para la evaluación de la interacción x genotipo de cultivares. Esta metodología se sustenta en las variaciones pequeñas de un genotipo en una misma localidad pero en diferentes años. Sobre este método, Atroch et al. (2000) y Scapim et al. (2000) señalan que no tiene las limitaciones de la regresión cuya interpretación es más difícil; es simple, fácil de aplicar y caracteriza los genotipos en un solo parámetro Pi (estimativo estadístico de adaptabilidad y estabilidad del comportamiento), que asocia la estabilidad con el rendimiento y, que a menor valor, mayor será su estabilidad. Además, la inclusión o retiro de uno o pocos cultivares no causa grandes variaciones en la estimación de los parámetros y permite la clasificación de los genotipos para su selección en procesos de mejoramiento genético.
Considerando que es necesario desarrollar avances tecnológicos en el cultivo de la estevia, el objetivo del presente trabajo fue determinar la estabilidad fenotípica de dos genotipos (cvs. Morita 1 y Morita 2) en función de tres densidades de planta y tres épocas de siembra en el Caribe colombiano.
Localización y material experimental
El estudio se realizó en el área experimental de la Universidad de Córdoba (13 m.s.n.m., 8° 52' N 76° 48' O), Montería, Colombia, en el ecosistema bosque seco tropical (bs-T) (Holdridge, 1967, citado por Jiménez-Saa, 2003), con una precipitación anual de 1200 mm, temperatura promedio de 28 °C, humedad relativa de 84% y brillo solar anual de 2108.2 horas (Palencia et al., 2006).
Se establecieron tres experimentos (épocas de siembra) durante el primer semestre de 2006 (abril 2 - junio 13) y los dos semestres agrícolas de 2007 (febrero 20 - abril 17 y noviembre 20 - enero 30/08, respectivamente). Las principales variables ambientales en cada época aparecen en el Cuadro 1. En cada experimento se utilizó un arreglo factorial de 2 (genotipos) x 3 (poblaciones) bajo un diseño experimental de bloques completamente al azar con 4 repeticiones. Los genotipos estudiados fueron cvs. Morita 1 (M1) y Morita 2 (M2) multiplicados en forma asexual, en densidades de planta de 6.25, 12.5 y 25 plantas/m2 establecidas bajo arreglos de 0.4 x 0.4, 0.2 x 0.4 y 0.2 x 0.2 m entre plantas y surcos respectivamente, con unidades experimentales de 210 plantas cada una.
Las variables dependientes medidas fueron relación hoja/tallo en base seca (RHT), rendimiento de hoja seca/ha (RHS) en kg/ha al momento de la floración, y contenido de glucósidos totales (GT) (St y RebA) medidos en porcentaje de peso seco de la hoja por el método de cromatografía líquida de alta eficacia (HPLC), utilizando en la fase móvil una mezcla de Acetonitrilo y agua (80:20), con una columna Kromasil 100-5HN2 (método isocrático) y un patrón de un 90% de pureza. Estos análisis fueron realizados por el Laboratorio de Control de Calidad de la empresa Prodalysa Ltda. en Valparaíso (Chile).
Análisis de resultados
Para identificar la importancia de los efectos de los componentes experimentales del modelo mixto se hicieron análisis de varianza individuales y combinados, considerando los genotipos y las poblaciones como efectos fijos y como aleatorio las épocas; las interacciones se descompusieron con tablas de doble entrada y comparaciones por la prueba de F (5%), utilizando el programa SAS (versión 9.1).
En el método de Lin y Binns (1988) el desempeño general de los genotipos se define como el cuadrado medio de la distancia entre la media del cultivar y la respuesta media máxima para todas las épocas, de tal modo que los genotipos y las poblaciones con menores valores corresponden a los de mayor estabilidad. De acuerdo con esta definición, el estimador Pig se determina por la ecuación siguiente:
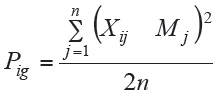
donde,
Pig = estadístico del parámetro de estabilidad del cultivar i,
Xij = respuesta de la variable dependiente del i ésimo cultivar o población en la j ésima época,
Mj = respuesta máxima observada entre todos los cultivares y poblaciones en la época j,
n = número de épocas.
Carneiro (1998) descompuso el estadístico Pig, con el fin de identificar los genotipos superiores en ambientes favorables y desfavorables, utilizando la misma metodología de clasificación de ambientes de Eberhart y Russell (1966). En esta descomposición se estiman los estadísticos siguientes:
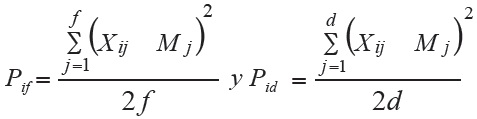
donde:
Pif = estadístico de Pig para cultivares y poblaciones favorables,
Pid = estadístico de Pig para cultivares y poblaciones desfavorables,
f = número de épocas favorables,
d = número de épocas desfavorables,
Xij y Mj = como fueron definidos anteriormente.
De esta manera, la selección de los cultivares y poblaciones puede ser realizada para las diferentes épocas (general y específica), siendo mucho más completa que la propuesta original.
El análisis de varianza combinado para las variables consideradas aparece en el Cuadro 2. Los resultados muestran que tanto la RHT como la RHS fueron afectados (P < 0.01) por los efectos individuales de la época, el genotipo y la población, mientras que los GT sólo mostraron diferencias por el efecto de la época y el genotipo, lo cual confirma que las condiciones ambientales en los semestres del estudio fueron diferentes, los genotipos son genéticamente diferentes y las poblaciones difieren de manera importante en número. Resultados similares encontraron Carbonell y Pompeu (2000) y Prado et al. (2001) cuando estudiaron el efecto de cinco y tres épocas de siembra en soya y fríjol, respectivamente.

Con excepción de las interacciones genotipo x época y población x época, que registraron variaciones significativas para la RHS (P < 0.01), las demás combinaciones no fueron significativas. Esto indica que para esta variable, los genotipos y poblaciones presentaron un comportamiento diferente en función de las épocas de siembra, posiblemente por los cambios ambientales ocurridos en los períodos del estudio, y demuestra la dificultad para recomendar de manera general cultivares y poblaciones, ya que sus expresiones fenotípicas se alteran bajo los ambientes del Caribe colombiano. Esto resalta la importancia de recurrir a estudios detallados de la interacción genotipo x ambiente, con el fin de realizar recomendaciones mucho más precisas y confiables para cultivares de esta especie.
La presencia de una interacción significativa con el ambiente obedece, en parte, a la ocurrencia de algún tipo de estrés (Carbonell y Pompeu, 2000), el cual puede ser ocasionado, entre otros factores, por cambios de temperatura, radiación solar, humedad relativa y precipitación (Calviño y Sadras, 1999; Montemayor et al., (2006; Jaitak et al., 2008). La interacción de segundo orden no fue significativa (P > 0.05), lo que significa que una parte de los efectos de ésta pueden ser explicados por las épocas, como lo demostraron en caña de azúcar Rea y de Sousa-Vieira (2001).
Los coeficientes de variación experimental oscilaron entre 13.5% y 21.5%, lo que evidencia una buena precisión de los datos experimentales (Pimentel-Gomes, 1985); los coeficientes de determinación se consideran altos para RHS (0.92) y aceptables para RHT (0.60) y GT (0.60), por tanto en este estudio el modelo puede ser utilizado para interpretar la estabilidad (Prado et al., 2001).
Relación hoja/tallo (RHT)
La RHT indica la eficiencia del genotipo en la traslocación de asimilados a la lámina foliar respecto a la formación de tejidos de sostén (tallo), probablemente como una respuesta a condiciones favorables, especialmente la temperatura en el suelo, como lo destacan Stone et al. (1999). Este fenómeno le permite a la planta alcanzar de manera rápida mayor área foliar, interceptar la radiación solar en forma eficiente, mayor producción de biomasa y consecuentemente, aumentar las tasas de fotosíntesis. Los efectos individuales altamente significantes (P < 0.01) de la época, el genotipo y la población sobre la RHT indican que en estevia esta relación respondió de manera diferencial al variar cada uno de estos factores (Cuadro 3).
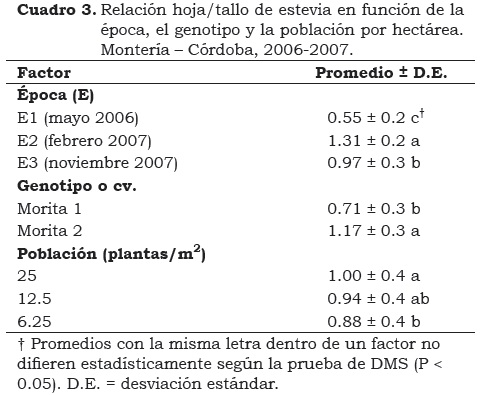
Para el efecto de épocas, los resultados indicaron que la RHT fue favorecida (P < 0.01) cuando el cultivo se sembró en febrero de 2007 (E2), alcanzando valores 1.3 y 2.3 veces el valor de la RHT en las siembra de noviembre del mismo año (E3) y mayo de 2006 (E1), respectivamente.
Los valores de radiación fotosintéticamente activa (RFA) que aparecen en el Cuadro 1, indican que en las tres épocas de siembra ocurrieron, en promedio, 15.05; 21.61 y 18.28 MJ/m2 por día para E1, E2 y E3, respectivamente. A pesar de que existen otros factores que ayudan a revelar el comportamiento de la RHT, tales como las diferencias en la temperatura promedio (27.7, 29.1 y 28.1 °C para la E1, E2 y E3 respectivamente), la oferta de RFA podría ser determinante para explicar las ventajas de la siembra en febrero de 2007 (E2), lo cual indica que la planta de estevia favorece la distribución de asimilados hacia la hoja en ambientes de mayor radiación, como lo encontraron Midmore y Rank, 2002; Jarma et al., 2006 y Jarma, 2008.
La eficiencia fisiológica en función de la radiación indica que el cv. Morita 2 tiene mayor índice de área foliar (IAF) que el cv. Morita 1 (datos no presentados) cuando los niveles de radiación aumentan (Jarma et al., 2005) lo que a su vez, explica las diferencias observadas entre genotipos en este trabajo y confirma que presentan sensibilidad diferente a los cambios de luz y la fotosíntesis.
Los resultados revelan que la RHT es directamente proporcional al número de plantas/m2, al menos en las densidades evaluadas en el estudio. En cultivos de estevia en el Caribe húmedo colombiano, en poblaciones entre 12.5 y 25 plantas/m2, se han encontrado consistentemente aumentos importantes en el IAF (Jarma, 2008; Espitia et al., 2009) lo que puede significar que estas poblaciones favorecen un balance de biomasa hacia el tejido foliar respecto al tallo, tal como lo indican Bruggink y Heuvelink (1987). Probablemente la RHT observada en altas poblaciones en estevia, es una respuesta a la competencia intra-específica que promueve una variación de la arquitectura del dosel, el cual se adapta a altas poblaciones y/o ambientes (Barot y Gignoux, 2004; Vance y Nevai, 2007; Combatt et al., 2008).
Rendimiento de hoja seca por hectárea (RHS)
La región Caribe colombiana es un área geográfica extensa y contrastante, la presencia de interacción de genotipo x épocas de siembra para el RHS (kg/ha), amerita minimizar los efectos de las épocas, lo que es posible a través de la identificación de cultivares con estabilidad fenotípica (Cuadro 4). El análisis de estabilidad fenotípica, de acuerdo con la metodología de Lin y Binns (1988), la cual define como genotipo superior aquel que registre el menor valor Pig indica que el cv. Morita 2 (Pig = 0) es el de mayor interés para los productores, debido a que dicho valor se atribuye a la poca desviación genética a través de los ambientes. Por su parte, el cv. Morita 1 presentó un valor de Pig = 238533, probablemente debido a su alta desviación genética, como ocurre con otras especies (Scapim et al., 2000).
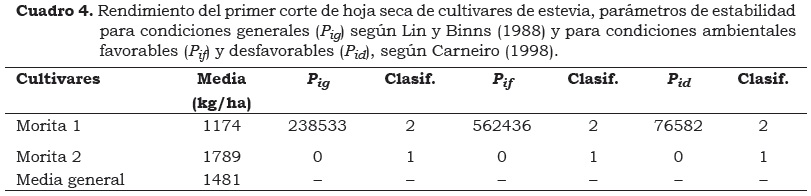
El comportamiento del cv. Morita 2 como genotipo estable es corroborado con la aplicación de la metodología de Carneiro (1998), la cual al descomponer el comportamiento de los genotipos en ambientes favorables (Pif) y desfavorables (Pid), indica que fue estable y de superior comportamiento al cv. Morita 1 en ambos ambientes (Cuadro 4). Por tanto, en la producción de hoja seca/ha, el cv. Morita 2 representa la mejor alternativa para los agricultores de la región Caribe colombiana.
En el Cuadro 5 se observa la respuesta diferencial del rendimiento de hoja seca (kg/ ha) de las poblaciones, en función de las condiciones edafoclimáticas de los períodos de estudio. De acuerdo con la metodología de Lin y Binns (1988) la densidad de 25 plantas/ m2 fue la de mayor estabilidad (Pig = 0), seguida de la densidad de 12.5 plantas/m2 (Pig = 83783) y la densidad de 6.25 plantas/ m2 que resultó ser la más inestable en las condiciones ambientales de la región.

De acuerdo con los criterios sugeridos por Carneiro (1998) la densidad de 25 plantas/m2 sería la más estable para ambientes tanto favorables como desfavorables, debido a los valores de Pif = 0 y Pid = 0, seguido de las densidades de 12.5 y 6.25 plantas/m2. Según lo anterior, los resultados de estabilidad encontrados señalan que el cv. Morita 2 en una densidad de 25 plantas/m2 es una buena alternativa para siembras en diferentes épocas en el Caribe húmedo de Colombia. No obstante, es necesario evaluar si las diferencias en rendimiento y calidad de la hoja entre 12.5 y 25 plantas/ m2, son lo suficientemente importantes en el momento de estimar los costos de semilla, si se tiene en cuenta que se encontraron diferencias entre poblaciones para los GT.
Los resultados de estabilidad observados en este estudio no difieren del comportamiento fisiológico de los genotipos y las poblaciones en otros trabajos, donde se ha reportado mayor producción de hoja por hectárea con el cv. Morita 2 (Jarma et al., 2006) y/o con altas poblaciones (Espitia et al., 2009).
Glucósidos totales
Tanto el genotipo (P < 0.05) como la época de siembra (P < 0.01) tuvieron efecto significativo sobre el contenido total de glucósidos en la hoja (Cuadro 2). Las pruebas de comparación de medias revelaron mayor contenido de glucósidos totales (GT) en las épocas E2 y E3 y en el cv. Morita 2 (Figura 1). Por épocas, el mayor valor de glucósidos totales se observó cuando los genotipos se cultivaron en la E3 (0.173 g/g de masa seca de hoja) el cual no difirió en la E2 (0.148 g/g) pero sí en la E1 (0.134 g/g) (Figura 1). Las variables del ambiente, principalmente la luz, tienen un efecto directo y positivo sobre la síntesis del isopentenil pirofosfato -IPP (Guenther et al., 1994), una molécula de cinco carbonos que es conocida como un isopreno activo de importancia en el alargamiento de la cadena terpénica. Esta ruta es la misma del ácido giberélico que, a partir del ácido mevalónico, conduce a la síntesis de las giberelinas y de los glucósidos de esteviol (Guevara-García et al., 2005). Con base en ello, es posible deducir que los altos niveles de luz observados en la región en E2 y E3 favorecieron esta ruta y por consiguiente la concentración de glucósidos totales.
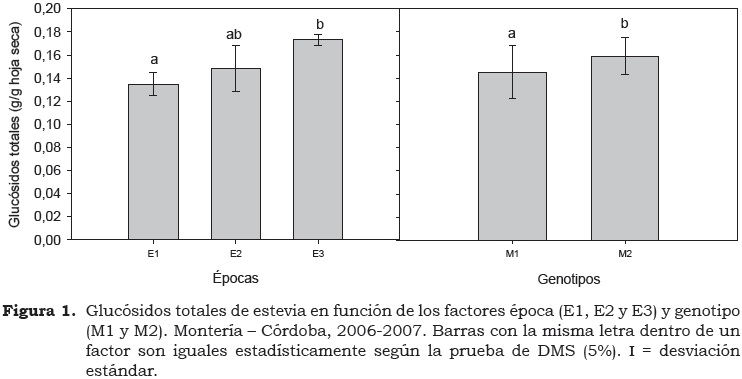
Aunque los valores de días a floración no fueron objeto de esta investigación, en las tres épocas de siembra se encontró que cuanto mayor RFA, los genotipos fueron más precoces, coincidiendo con otras especies hortícolas donde se ha reportado una correlación lineal positiva entre temperatura y precocidad (Real-Rosas et al., 2002; Ortegón et al., 2006). En este sentido, estos resultados coinciden con los reportados por Sumida (1980) quien indicó que genotipos de floración temprana tienden a un contenido de esteviósido superior y un menor rendimiento de biomasa. Contrariamente, otros autores han indicado que períodos de floración más tardíos permiten mayor tiempo de acumulación de glucósidos (Ramesh et al., 2006); sin embargo, para considerar ambas teorías y definir el período de floración de un cultivar, se deben considerar otros factores determinantes como el genotipo, la luminosidad y la competencia intraespecífica, entre otros.
Con base en las diferencias de GT entre el cv. Morita 1 (0.1446 ± 0.019 g/g hoja seca) y cv. Morita 2 (0.1586 ± 0.024 g/g hoja seca), los resultados de la presente investigación confirman que el proceso de mejoramiento genético del cv. Morita 2 fue conducido para obtener un recurso con mayor contenido de GT. Esta condición se mantiene estable con el genotipo, ya que la propagación de estevia se hizo a partir de esquejes (asexual), y se conserva, por tanto, las características genéticas del genotipo donante (Lee, 1982).
Es importante recordar que esta especie tiene un grado de mejoramiento genético relativamente incipiente y que, de acuerdo con varios estudios, se han encontrado por su autoincompatibilidad, poblaciones heterogéneas heterocigotas (Handro et al., 1993), con valores de GT que varían entre 4% y 16% (Nakamura y Tamura, 1985), los cuales están por debajo de los reportados en este estudio.
Estudios realizados por Brandle (1999) muestran que el rendimiento de hoja, RHT y contenido de glucósidos, poseen alta heredabilidad (75% - 83%). Con base en ello, la investigación en estevia también ha mostrado algunas correlaciones interesantes que han contribuido con los programas de mejoramiento genético, entre ellas: rendimiento de hoja/planta proporcional al número de brácteas, número de hojas y aunque no siempre, a la altura de planta (Buana, 1989; Shu y Wang, 1988); el contenido de GT muestra una correlación positiva con la RHT (Tateo et al., 1998), lo cual es consistente con lo encontrado en el presente trabajo; por otra parte, el grosor de la hoja, área foliar, contenido de clorofilas y proteínas y la tasa de fotosíntesis neta correlacionan positivamente con la relación RebA/ St (Shyu, 1994; Weng et al., 1996).
- Los análisis de estabilidad empleados permitieron identificar al cv. Morita 2, establecido en una densidad de 25 plantas/ m2, como una opción tecnológica estable para el uso de los productores en un rango de condiciones ambientales diferentes en la región Caribe de Colombia. No obstante, es importante considerar el costo de las semillas y el retorno de la inversión en este insumo, los cuales se aumentan significativamente al establecer esta población respecto a 12.5 plantas/m2.
- La relación hoja/tallo fue mayor para el cv. Morita 2, principalmente en ambientes con mayor oferta de RFA y cultivado en altas poblaciones.
- La calidad de la hoja, medida como el contenido total de glucósidos, indicó que esta variable es afectada positivamente por la oferta de RFA y que el cv. Morita 2 es consistentemente el genotipo con mayor contenido de glucósidos totales respecto al cv. Morita 1.
Los autores expresan sus agradecimientos a la Universidad de Córdoba por la financiación de la investigación; así mismo, a la empresa Prodalysa Ltda. en Valparaíso (Chile) a través de los doctores John Alarcón y Andrés Llanos del Laboratorio de Control de Calidad en Chile y Colombia, respectivamente, por su colaboración decidida en el análisis de las muestras de contenido de glucósidos totales.
Atroch, A.; Soares, A.; y Patto, M. 2000. Adaptabilidade e estabilidades de linhagens de arroz de sequeiro testadas no estado de Minas Gerais. Ciência e Agrotecnologia 24(3):541 - 548.
Barot, S. y Gignoux, J. 2004. Mechanisms promoting plant coexistence: can all the proposed processes be reconciled? Oikos 106(1):185 - 192.
Becker, H. 1981. Correlations among some statistical measures of phenothypic stability. Euphytica 30(3):835 - 840.
Brandle, J. E. 1999. Genetic control of rebaudioside A and C concentration in leaves of the sweet herb, Stevia rebaudiana. Canadian Journal of Plant Science 79(1):85 - 92.
Brandle, J. E.; Richman, A.; Swanson, A.; y Chapman; B. 2002. Leaf ESTs from Stevia rebaudiana: a resource for gene discovery in diterpene synthesis. Plant Mol. Biol. 50(4-5):613 - 622.
Brandle, J. E. 2005. Stevia, Natures Natural Low Calorie Sweetener. Disponible en: http://www.certinedorganicfood Stevia FAQ. (Fecha de revision: junio 12 de 2005).
Bruggink, G. T.; y Heuvelink, E. 1987. Influence of light on the growth of young tomato, cucumber and sweet pepper plants in the greenhouse: effects on relative growth rate, net assimilation rate and leaf area ratio. Scientia Horti. 31:161 - 174.
Buana, L. 1989. Determination of the required growth variables in an agronomic experiment with Stevia [Indonesian]. Menara Perkebunan 57(2):29 - 31.
Calviño, P. A. y Sadras, V. O. 1999. Interanual variation in soybean yield: interaction among rainfall soil depth and crop management. Field Crops Research 63(3):237 - 246.
Campos, R. 2007. Experiencias de la producción de Stevia en Brasil. En: III Simposio Internacional de la Stevia. Asunción Paraguay. 15 y 16 de noviembre de 2007. 35 p. Ministerio de Industria y Comercio de Paraguay Cámara Paraguaya de la Stevia Red de inversiones y exportaciones de Paraguay. Asunción, Paraguay.
Carbonell, S. y Pompeu, A. 2000. Estabilidade fenotípica de linhagens de feijoeiro en três ópocas de plantio no estado de Sao Paulo. Pesquisa Agropecuria Brasileira 35(2):321 - 329.
Carneiro, P. 1998. Novas metodologías de análisis da adaptabilidade e estabilidade de comportamiento. Tese Doutorado. Universidade Federal de Viçosa, Viçosa-MG Brasil. 168p.
Casaccia, J. y álvarez, E. 2006. Recomendaciones técnicas para la producción sustentable del Kaa Hee (Stevia rebaudiana) Bertoni en el Paraguay. Manual No. 8. p. 43. Caacupe Ministerio de Agricultura y Ganadería, Dirección de Investigación Instituto Agronómico Nacional Agrícola. Asunción-Paraguay.
Combatt, E.; Jarma, A.; y Maza, L. 2008. Crecimiento de Brachiaria decumbens Stapf y Cynodon nlemfuensis Vanderyst, en suelos sulfatados ácidos de Córdoba. MVZ Córdoba 13(2):1380 - 1392.
Dacome, A.; da Silva, C. C.; da Costa, C. E. M.; Fontana, J. D.; Adelmann, J.; y da Costa, S. C. 2005. Sweet diterpenic glycosides balance of a new cultivar of Stevia rebaudiana (Bert.) Bertoni: Isolation and quantitative distribution by chromatographic, spectroscopic, and electrophoretic methods. Process Biochemistry 40:3587 - 3594.
Eberhart, S. y Russell, W. 1966. Stability parameters for comparing varietes. Crop Science 6(1):36 - 40.
Espitia, M.; Montoya, R.; y Atencio, L. 2009. Rendimiento de Stevia rebaudiana Bert., bajo tres arreglos poblacionales en el Sinú Medio. U.D.C.A. Actualidad y divulgación científica 12(1):151 - 161.
Geuns, J. M. 2003. Stevioside. Phytochemistry 64(5):913 - 921.
Guenther, A.; Zimmerman, P.; y Wildermuth, M. 1994. Natural volatile organic compound emission rate estimates for U.S. woodland landscapes. Atmospheric Environment 28(6):1197 - 1210.
Guevara-García, A.; San Román, C.; Arroyo, A.; Cortes, M.; Gutiérrez-Nava, de la Luz, M.; y León, P. 2005. Characterization of the Arabidopsis clb6 mutant illustrates the importance of posttranscriptional regulation of the methyl-d-erythritol 4-phosphate pathway. Plant Cell 17(2):628 - 643.
Handro, W.; Ferreira, C. M.; y Floh, E. I. S. 1993. Chromosomal variability and growth rate in cell suspension cultures of Stevia rebaudiana. Plant Science 93(1):1 - 2.
Hearn, L. K. y Subedi, P. P. 2008. Determining levels of steviol glycosides in the leaves of Stevia rebaudiana by near infrared reflectance spectroscopy. Journal of Food Composition and Analysis 22(2):165 - 168.
Hsieh, M. y Goodman, H. 2005. The Arabidopsis IspH homolog is involved in the plastid nonmevalonate pathway of isoprenoid biosynthesis. Plant Physiology 138(2):641 - 653.
Jaitak, V.; Gupta, A.; Kaul, A. V.; y Ahuja, P. 2008. Validated high-performance thin-layer chromatography method for steviol glycosides in Stevia rebaudiana. Journal of Pharmaceutical and Biomedical Analysis 47(4-5):790 - 794.
Jarma, A. 2008. Estudios de adaptación y manejo integrado de estevia (Stevia rebaudiana Bert.): nueva alternativa agroindustrial del Caribe colombiano. Una revisión. Rev. Colomb. Cienc. Hort. 2(1):109 - 120.
Jarma, A. 2009. Avances de la investigación en Stevia en Colombia. En: Memorias IV Simposio internacional de la Stevia Ka´a Heê. 12 y 13 de Marzo de 2009. 38 p. Ministerio de Industria y Comercio de Paraguay Cámara Paraguaya de la Stevia Red de inversiones y exportaciones de Paraguay. Asunción, Paraguay.
Jarma, A.; Rengifo, T.; y Araméndiz, H. 2005. Aspectos fisiológicos de estevia (Stevia rebaudiana Bertoni) en el Caribe colombiano. I. Efecto de la radiación incidente sobre el área foliar y la distribución de biomasa. Agronomía Colombiana 23(2):207 - 216.
Jarma, A.; Rengifo, T.; y Araméndiz, H. 2006. Aspectos fisiológicos de estevia (Stevia rebaudiana Bertoni) en el Caribe colombiano. II. Efecto de la radiación incidente sobre los índices de crecimiento. Agronomía Colombiana 24(1):38 - 47.
Jiménez-Saa, H. 2003. Anatomía del sistema de ecología basada en zonas de vida de L. R. Holdridge. 32 p. San José, CR, Centro Científico Tropical.
Kasahara, H.; Hanada, A.; Kuzuyama, T.; Takagi, M.; Kamiya, Y.; y Yamaguchi, S. 2002. Contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthesis of gibberellins in Arabidopsis. J. Biol. Chem. 277(47):45188 - 45194.
Lee, J. I. 1982. New high Rebaudioside-A Stevia variety Suweon 11. p. 4. Research Reports, Office of Rural Development. S. Korea.
Lin, C. y Binns, M. 1988. A method for analyzing cultivar x location x year experiments: A new stability parameter. Theoretical Aplied Genetics 76(3):425 - 430.
Maya, D. 2000. Stevia rebaudiana Bertoni. Primera edición. Corpoica. Bogotá, D.C. Colombia. 16 p.
Midmore, D. y Rank, A. 2002. A new rural industry - Stevia - to replace imported chemical sweeteners. p. 55. A report for the Rural Industries Research and Development Corporation RIRDC. Kingston, AU.
Montemayor, J.; Zermeño, J. A.; Olague, J.; Aldaco, R.; Fortis, M.; Salazar, E.; Cruz, J.; y Vázquez- Vázquez, C. 2006. Efecto de la densidad y estructura del dosel de maíz en la penetración de la radiación solar. FYTON. 75:47 - 53.
Nakamura, S. y Tamura, Y. 1985. Variation in the main glycosides of stevia. Japanese Journal of Tropical Agriculture 29(2):109 - 115.
Ortegón, A.; Díaz, F.; González, J.; y Garza, I. 2006. La temperatura en la etapa reproductiva del cultivo de canola (Brassica napus L.). Agric. Téc. Méx. 32(3):259 - 265.
Palencia, G.; Mercado, T.; y Combatt, E. 2006. Estudio agro-meteorológico del Departamento de Córdoba. 126 p. Facultad de Ciencias Agrícolas, Universidad de Córdoba. Gráficas del Caribe. Montería Córdoba.
Pimentel-Gomes, F. 1985. Curso de estatística experimental. 8a ed. 450p. Nobel. Sao Paulo Brasil.
Prado, E.; Hiromoto, D.; Campos, V.; Utumi, M.; y Rostand, A. 2001. Adaptabilidade e estabilidade de cultivares de soja em cinco épocas de plantio no cerrado de Rondônia. Pesq. Agropec. Bras. 36(4):625 - 635.
Ramesh, K.; Singh, V.; y Megeji, N. W. 2006. Cultivation of stevia [Stevia rebaudiana (Bert.) Bertoni]: A comprehensive review. En: Advances in Agronomy, v. 89. 360 p. Academic Press. San Diego, California - USA.
Rea, R. y Sousa-Vieira, O. de 2001. Interacción genotipo x ambiente y análisis de estabilidad en ensayos regionales de caña de azúcar en Venezuela. Caña de azúcar 19:3 - 15.
Real-Rosas, M.; Lucero-Arce, A.; Toyota, M.; López- Aguilar, R.; y Murillo-Amador, B. 2002. Variedades precoces de brécol para la diversificación agrícola en zonas áridas. INCI. 27(5):247 - 251.
Scapim, C.; Rodrigues, V.; Braccini, A.; Cruz, C.; Bastos-Andrade, C.; y Conçalves, M. 2000. Yield stability in maize (Zea mays L.) and correlations among the parameters of the Eberhart and Russell, Lin and Binns, and Huehn models. Genetics and Molecular Biology 23(2):387 - 393.
Shu, S. Z. y Wang, W. Z. 1988. Variation in quantitative characters in Stevia. Acta Agronomica Sinica 14(2):167 - 173.
Shyu, Y. T. 1994. Effects of harvesting dates on the characteristics, yield, and sweet of Stevia rebaudiana Bertoni. Journal of Agricultural Research of China 43(1):29 - 39.
Stone, P. J.; Sorense, I. B.; y Jamieson, P. D. 1999. Effect of soil temperature on phenology, canopy development, biomass and yield of maize in a cool-temperature climate. Field Crops Research 63(2):169 - 178.
Sumida, T. 1980. Studies on Stevia rebaudiana Bertoni as a possible new crop for sweetening resource in Japan. J. Central Agricultural Exp. Stn. 31:1 - 71.
Tateo, F.; Mariotti, M.; Bononi, M.; Lubian, E.; Martello, S.; y Cornara, L. 1998. Stevioside content and morphological variability in a population of Stevia rebaudiana (Bertoni) Bertoni from Paraguay. Italian Journal of Food Science 10(3):261 - 267.
Totté, N; Charon, L.; Rohmer, M.; Compernolle, F.; Baboeuf, I.; y Geuns, J. 2000. Biosynthesis of the diterpenoid steviol, an entkaurene derivative from Stevia rebaudiana Bertoni, via the methylerythritol phosphate pathway. Tetrahedron Letters 41(33):6407 - 6410.
Totté, N.; Van den Ende, W.; Van Damme, E.; Compernolle, F.; Baboeuf, I.; y Geuns, J. 2003. Cloning and heterologous expression of early genes in gibberellin and steviol biosynthesis via the methylerythritol phosphate pathway in Stevia rebaudiana Bertoni. Can. J. Bot. 81(5):517 - 522.
Vance, R. R. y Nevai, A. L. 2007. Plant population growth and competition in a light gradient: A mathematical model of canopy partitioning. Journal of Theoretical Biology 245(2):210 - 219.
Weng, X. Y.; Sun, J. Y.; y Zang; R. C. 1996. Study on the growth and physiological characteristics of Stevia rebaudiana SM4 [Chinese]. Journal of Zhejiang Agricultural University 22(5):538 - 540.
Yoda, S. K.; Marques, M. O.; Petenate, A. J.; y Meireles, A. A. 2003. Supercritical fluid extraction from Stevia rebaudiana Bertoni using CO2 and CO2 + water: extraction kinetics and identification of extracted components. Journal of Food Engineering 57(2):125 - 134.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2011 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.





















