



Published
Evaluation of uva camarona (Macleania rupestris Kunth A.C. Smith) propagation with air layering
Keywords:
Ericaceae, indole-butyric acid, vegetative propagation, roots, callus. (es)Downloads
Uva camarona (Macleania rupestris Kunth A.C. Smith) belongs to the Ericaceae family and grows in páramo and subpáramo areas in Colombia, between 2,200 and 3,500 m a.s.l. This plant presents edible berries that serve as a source of food and small income for local communities. The absence of a propagation protocol for this species limits its use. This study aimed to evaluate the effectiveness of asexual propagation of M. rupestris with air layering, using indole-butyric acid (IBA) as a rooting hormone at different concentrations: 0, 500, 1,000, and 2,000 mg L-1. The results showed that an exogenous application of IBA accelerated the rooting process in the layered zone, with a notable emission of first adventitious roots at 60 days from the start of the experiment. The treatments of 500 and 1,000 mg L-1 IBA had the highest number of roots per layer, being two to three times higher than those presented in the control. No significant differences were seen in root length between treat- ments. The treatments of 500 and 1,000 mg L-1 IBA showed a high production for the dry weights of the roots and callus, with a higher weight of callus compared to root weight. Air layering negatively affected the longitudinal growth of the branches, since their average growth rate was 1.49 cm per month, while the growth of intact branches was 2.78 cm per month. The re- sults suggest that the best concentration for rooting was 1.200 mg L-1 IBA because it had the largest number and dry weight of roots in air-layered M. rupestris.
PROPAGATION & TISSUE CULTURE
Evaluation of uva camarona (Macleania rupestris Kunth A.C. Smith) propagation with air layering
Evaluación de la propagación de uva camarona (Macleania rupestris Kunth A.C. Smith) por medio de acodos aéreos
Santiago Durán-Casas1, Clara Veloza Suan1 ,Stanislav Magnitskiy 1 and Héctor Orlando Lancheros2
1Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia).
sduranc@unal.edu.co
2Research Division, Jardín Botanico José Celestino Mutis. Bogota (Colombia).
Received for publication: 28 August, 2012. Accepted for publication: 29 March, 2013.
ABSTRACT
Uva camarona (Macleania rupestris Kunth A.C. Smith) belongs to the Ericaceae family and grows in páramo and subpáramo areas in Colombia, between 2,200 and 3,500 m a.s.l. This plant presents edible berries that serve as a source of food and small income for local communities. The absence of a propagation protocol for this species limits its use. This study aimed to evaluate the effectiveness of asexual propagation of M. rupestris with air layering, using indole-butyric acid (IBA) as a rooting hormone at different concentrations: 0, 500, 1,000, and 2,000 mg L-1. The results showed that an exogenous application of IBA accelerated the rooting process in the layered zone, with a notable emission of first adventitious roots at 60 days from the start of the experiment. The treatments of 500 and 1,000 mg L-1 IBA had the highest number of roots per layer, being two to three times higher than those presented in the control. No significant differences were seen in root length between treatments. The treatments of 500 and 1,000 mg L-1 IBA showed a high production for the dry weights of the roots and callus, with a higher weight of callus compared to root weight. Air layering negatively affected the longitudinal growth of the branches, since their average growth rate was 1.49 cm per month, while the growth of intact branches was 2.78 cm per month. The results suggest that the best concentration for rooting was 1.200 mg L-1 IBA because it had the largest number and dry weight of roots in air-layered M. rupestris.
Key words: Ericaceae, indole-butyric acid, vegetative propagation, roots, callus.
RESUMEN
La uva camarona (Macleania rupestris Kunth A.C. Smith) es una planta de la familia Ericaceae que crece en zonas de páramo y subpáramo en Colombia, entre los 2.200 y 3.500 msnm. Presenta bayas comestibles, las cuales sirven como alimento o fuente de pequeños ingresos para las comunidades locales. La ausencia de un protocolo de propagación de esta especie limita su aprovechamiento. El presente trabajo tuvo el objetivo de evaluar la eficacia de la propagación asexual mediante acodos aéreos, utilizando como hormona de enraizamiento ácido indolbutírico (AI B) a diferentes concentraciones: 0, 500, 1.000 y 2.000 mg L-1. Los resultados mostraron que la aplicación exógena de AI B aceleró el proceso de enraizamiento en la zona de acodamiento, observando emisión de las primeras raíces adventicias a los 60 días de iniciado el ensayo. Los tratamientos de 500 y 1.000 mg L-1 de AI B presentaron el mayor número de raíces por acodo, siendo dos a tres veces mayor que las presentes en el control. No hubo diferencias significativas en cuanto a longitud de raíces entre los tratamientos. La mayor producción de peso seco de raíces y de callo se presentó en los tratamientos de 500 y 1.000 mg L-1 de AI B, mostrando un mayor peso de callo comparado con el de raíces. El acodamiento afectó negativamente el crecimiento longitudinal de las ramas, ya que la tasa de crecimiento promedio de éstas fue de 1,49 cm por mes, mientras que las que no se acodaron presentaron un crecimiento de 2,78 cm por mes. Los resultados sugieren que con la concentración aproximada de 1.200 mg L-1 de AI B se obtienen el mayor número y peso seco de raíces en los acodos de M. rupestris.
Palabras clave: Ericaceae, ácido indolbutírico, propagación vegetativa, raíces, callo.
Introduction
Within the great diversity of plant species of high use potential in Colombia, Andean fruit trees are found, with only a few domesticated species and the majority grown in the wild. These plants account for an important source of food and/or income for local communities (National Research Council, 1989; Lagos-Burbano et al., 2010).
Many of these fruit species are gaining value, given their high commercial potential due to the excellent fruit quality for direct consumption (Vásquez and Coimbra, 2002; Sanjinés et al., 2006) and increasing demand in North America, Europe, and Japan, where they are known as exotic Andean fruits (Lagos-Burbano et al., 2010).
The Ericaceae family is one of the most representative in the Andean forests, mainly in mountain cloud forests, as well as páramo and subpáramo regions, where it plays an important ecological role (Luteyn and Pedraza;-Peñalosa, 2008; Lagos-Burbano et al., 2010) as a water regulator, food source and, in some cases, source of income for the inhabitants of the region (Lagos-Burbano et al., 2010).
Macleania rupestris Kunth A.C. Smith is one of the undomesticated fruit species of the Ericaceae family. This plant is found in wild areas of the Andean region at altitudes between 1,500 and 4,100 m (Acero and Bernal, 2003; Luteyn and Vidal, in press). In Colombia, M. rupestris is commonly called uva camarona or camaronera and is located in wild mountain habitats as a constituent of native vegetation (Sturm and Rangel, 1985; Lagos-Burbano et al., 2010). This species is found in the provinces of Antioquia, Boyacá, Caldas, Cundinamarca, Huila, Meta, Nariño, Norte de Santander, Putumayo, Risaralda, Santander, and Magdalena in the Sierra Nevada de Santa Marta, growing between 2,200 and 3,500 m (Cardozo et al., 2009; Lagos- Burbano et al., 2010).
Macleania rupestris is an epithytic shrub measuring from 0.6 to 2.5 m tall with branching originating almost at the base of the stems (Acero and Bernal, 2003; Cardozo et al., 2009; Lagos-Burbano et al., 2010). Sometimes, this plant grows near tall trees and ascends through their branches imitating a vine species. The stems of M. rupestris are wavy and have a clear and scaly bark. The young leaves are red and light green when mature (Fig.1A), and are alternate, simple, shiny, and fleshy (Cardozo et al., 2009). The plant has axillary inflorescences located at the apex of the branches, without apical flowers; the inflorescences are racemose with four to 20 flowers. The flowers have a bell-shaped calyx limb typical of the Ericaceae family; the perianth of the flower and its pedicel are pink to red (Fig.1B), with cylindrical, succulent sepals (De Valencia and De Carrillo, 1991; Cardozo et al., 2009; Lagos-Burbano et al., 2010). This species grows in rocky areas, forests, and bushes as a part of native vegetation. It is resistant to strong winds and frost, and requires good sun exposure (Rodríguez and Peña, 1984).

The fruit is edible and originates from an inferior ovary (Font Quer, 1965); it is stalked, and a berry type. Its color during the ripening process changes from light pink, pale green, and whitish to dark purple and almost black in senescence (Fig.1: C and D), so the skin of the fruit is purple in maturity and the pulp has a light color similar to a grape. The fruit is fleshy, having a diameter of 1.0-2.0 cm and an average weight of 2.6 g (De Valencia and De Carrillo, 1991; Acero and Bernal, 2003; Cardozo et al., 2009). The fruits are arranged on a racemose axis with acropetal maturation similar to that observed in the flowers. The flowers are pendulous, but the fruits are not, which indicates that the twisting of petioles occurs during fruit development (De Valencia and De Carrillo, 1991).
Bromatological characterization of M. rupestris fruits by Amaya and Rincon (1989) showed that they have a total acidity of 0.49% citric acid, 12.5°Brix soluble solids, 1.8% crude fiber, 1.3% crude protein, and 0.08 g tannin in 100 g of pulp. The contents of mineral elements per 100 g of pulp were 136.9 mg calcium, 0.97 mg iron, 5.24 mg potassium, 8.69 mg sodium, and 10.3 mg phosphorus; and the sugars (per 100 g of pulp): 15.9 g fructose, 1.62 g of sucrose, 13.5 g of α-glucose, and 16.1 of Β-glucose (Amaya and Rincon, 1989).
Macleania rupestris plants have on average of 14 fruits per inflorescence, with two peaks of production per year in Colombia (De Valencia and De Carrillo, 1991), the first one from mid-January to April, and the second one from August to mid-November. These periods fluctuate with rainfall distribution; and with additional rainfall in dry weather, the periods of flowering and fruiting overlap (Cardozo et al., 2009).
Human activities have led to about 75% of forestland located in the northern Andes being transformed to the point of total deforestation (Henderson et al., 1991). These ecosystems are home to about 80% of the different neotropical Ericaceae species and some of them could be a source of income for local communities. For this reason, reforestation programs of these environments are needed, employing these species, taking into account that many Ericaceae plants are successful colonizers of natural or anthropogenically disturbed areas (Luteyn, 2002). In this sense, the potential use of M. rupestris is not only limited to its use as a commercial crop, but also to land restoration.
The absence of an adequate M. rupestris propagation protocol, that is affordable and accessible to small growers is one of the obstacles to the introduction of this plant into commercial cultivation and its use for the recovery of páramo and subpáramo ecosystems. Therefore, it is necessary to undertake research that seeks to procure consistent and reliable plant material of M. rupestris through the evaluation of various methods of plant propagation.
Air layering is an ancient method used for vegetative propagation of woody plants that can ensure the asexual propagation of a species when sexual reproduction is very slow or ineffective (Adriance and Brison, 1955), as is the case of M. rupestris (Acero and Bernal, 2003; Cardozo et al., 2009), or when there is no detailed protocol for propagation through cuttings (Adriance and Brison, 1955).
The methodology of air layers can obtain 100% rooting because the branch is rooted to the mother plant; in addition, homogeneous plants are obtained within a relatively short period of time (Adriance and Brison, 1955; Lambe et al., 1990). In the literature, no data are found on the possible propagation of M. rupestris with air layering, which would be a new method for this species. Therefore, this study aimed to evaluate the possibility of asexual propagation of M. rupestris by air layering, in order to provide the tools and methodologies that would facilitate the commercial propagation and use of this species.
Materials and methods
The study was carried out in the period from August to December 2011, in the collection of páramo plants of the José Celestino Mutis Botanical Garden, Bogotá (Colombia), located at the coordinates 04°40'01.2'' N and 74°05'57.4'' W, and altitude 2,552 m a.s.l. During this period, the following were registered: 635.5 mm of cumulative rainfall, average temperature of 18°C, average photosynthetically active radiation of 456.6 µmol m-2s-1 and wind speed of 0.14 km h-1 (Sutron® weather station, José Celestino Mutis Botanical Garden). The vegetation in association with Macleania AC rupestris Kunth Smith plants was typical of the páramo and subpáramo ecosystems of Colombia, with species belonging to the families: Ericaceae, Melastomataceae, Asteraceae, among others. The sandy loam soil where the M. rupestris plants were established had an average pH of 5.43.
Adult individuals of M. rupestris were selected with active vegetative growth, located under similar full light exposure, and without visual symptoms of nutritional deficiencies or diseases. The selected plants had approximately 10 to 15 stems and an average height of 2.5 m. The layering was performed on six individual plants, making four air layers per plant, with each layer corresponding to a specific treatment; the branches selected had one layer per branch. Therefore, in total, six replicates were established for each treatment, so that every plant had a full set of treatments.
Lignified orthotropic stems at the vegetative growth stage (with presence of few flowers, if any, and without the presence of fruits) were chosen on every selected plant, with homogeneous characteristics, presenting turgid leaves and vigorous growth. Selected branches underwent a complete ring cut in the middle of the branch, removing a 4.0 cm portion of the bark with a grafting knife previously disinfected with 0.5% sodium hypochlorite. The application of the rooting hormone was performed immediately after removing the bark. With the aid of a brush, the ring was homogeneously covered with the powder mixture, which contained indolebutyric acid (IBA) (99% purity, Biopack®- Buenos Aires, Argentina) and technical talc as an inert material in proportions that yielded four treatments: 0, 500, 1,000, and 2,000 mg L-1 IBA.
After application of the hormone, 200 g of previously moistened peat moss without nutrients (Klasmann-Deilmann GmbH®, Geeste, Germany) were arranged around the cut that was further covered with transparent and black plastic, thus, avoiding light and moisture loss.
Biweekly irrigation was performed with 5 mL of deionized water on each layering, using a syringe. Monthly, the length of the branches was measured with a measuring tape (from the top of the girdling zone to the apex of the branch) and the diameter of each branch was recorded with a limb caliper 30 cm from the top of the layer (accuracy ±0.01 cm). The layered branches had an average length of 67 cm and width of 1.1 cm when the experiment was established. Furthermore, monitoring of root emission on each layer was performed after 60 d from the start of the experiment.
The measurement of quantitative variables was performed at the Laboratory of Plant Physiology, Faculty of Agronomy of the Universidad Nacional de Colombia, Bogota. This took place 120 d after the start of the experiment, performing destructive sampling of all layered branches. The callus and adventitious roots were removed using a scalpel, weighed immediately using a high precision analytical balance (±0.001 g, Ohaus®, Ohaus Corp., Pine Brook, NJ ) and the dry weight was determined after placing the samples in oven at 65°C for 48 h. The variables measured were: number and length of adventitious roots, fresh and dry weights of roots and callus. A photographic record of the results was also performed.
The statystical analysis of the results was carried out with the help of SAS ® version 9.0 and the differences among the treatments were evaluated by AN OVA with a significance level of 95% (P=0.05). The polynomial regressions were additionally determined among the variables.
Results and discussion
At 120 d after the start of the experiment, 100% rooting for all treatments was found, without any dead layered branches. This confirms the data described by Hartmann et al. (2010), who indicated that the layering technique is successful in propagating plants because the layered branch is not separated from the mother plant and, therefore, continues to receive water and nutrients. These data agree with the results found in air-layers of agraz (Vaccinium meridionale Swartz) that presented root emission after 90 to 120 d of the experiment (Ligarreto, 2011).
The results obtained in the control treatment might be due to the influence of buds and leaves present in the layered branch, which stimulate rooting through the production of auxins in these organs (Hartmann et al., 2010). In turn, branch girdling causes blockage in the acropetal and basipetal flow of photoassimilates in the phloem, causing an accumulation of several metabolites (organic compounds, carbohydrates, auxins, etc.) above this region (Alves-de Oliveira et al., 1999) that favors the formation of callus and adventitious root primordia.
Furthermore, the absence of light in the area of root formation, use of substrates to provide continuous moisture, and moderate temperatures are factors that favor rooting in the girdling zone (Ramirez-Villalobos and Urdanetα- Fernández, 2004; Hartmann et al., 2010). In this sense, the results observed in the control treatment might be due to the endogenous content of phytohormones in the branches and provision of beneficial conditions for rooting.
However, the exogenous application of auxin-type growth regulators can speed up the rooting process (Hackett, 1988; Bacarín et al., 1994; Rahman et al., 2000; Naz and Aslam, 2003), which was reflected in the emission of first roots at 60 d of the experiment for the treatments with IBA, with a maximum length of 3.5 cm. Contrary to this, the control treatments presented only a development of callus for the same period of observation.
Number and length of adventitious roots
The results show that the treatments of 500 and 1,000 mg L-1 IBA produced the highest number of adventitious roots with layering (Figs.2 and 3), with average values of 27 and 32 roots, respectively, which represent about two to three times higher root emission than that observed in the control, which had an average of 12 roots. Similar results were found by Naz and Aslam (2003) in mango (Mangifera indica L.) air layers, where the treatments with 2,000 and 3,000 mg L-1 IBA produced an average of 39 and 31 roots, respectively, while the control treatment had only 25 roots.
As shown in Fig.2, there was a reduction of root emission in the 2,000 mg L-1 IBA treatment as compared with the other hormone treatments. However, statistically significant differences existed between the IBA treatments and the control. Likewise, according to the polynomial regression model, the highest number of roots in the M. rupestris layerings was at approximately 1,200 mg L-1 IBA, so this IBA concentration could be the most suitable for this form of propagation.
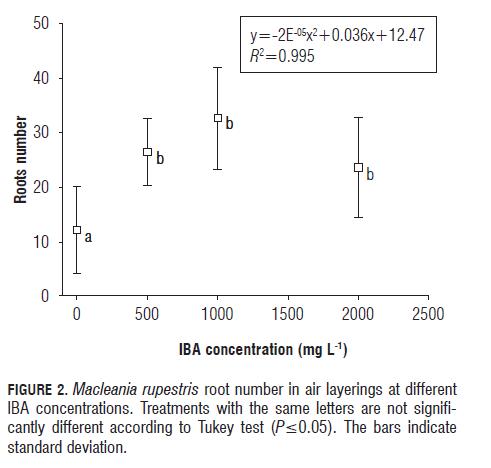
In Fig.4, one can see that an exogenous application of auxin had an effect on root formation for the treatments of 0 mg L-1 (control) and 1,000 mg L-1 IBA. This behavior is consistent with the findings of Paul and Aditi (2009), where the maximum number of roots in air layers made on Syzygium javanica was obtained with the concentration of 1,000 mg L-1 IBA, while the concentrations of 500 and 2,500 mg L-1 showed no significant differences compared to the control.

In studies of Jatropha curcas L., the root number increased with the increasing concentration of napthtaleneacetic acid (NAA ) and IBA to yield the maximum number of roots in layered branches treated with 1,500 mg L-1 NAA followed by IBA at the same concentration (Dhillon et al., 2011). Dhillon et al. (2011) showed that the effect of growth regulators (indoleacetic acid IAA , NAA , IBA) and their concentrations significantly influenced the rooting percentage and number of roots, but did not significantly affect the length of the roots of Jatropha curcas L. in air layers. This agrees with the findings of the length of adventitious roots in layers of M. rupestris, where no significant differences between the treatments were found; all roots had an average length of 5.0 cm. According to Tchoundjeu et al. (2002), auxins promote meristematic differentiation, and thus, contribute to the formation of root primordia in the girdling zone of the branch. For this reason, exogenous IBA might affect the number of adventitious roots more, and to a lesser extent the length of the roots.
Weight of adventitious roots and callus
Auxins promote the formation of callus and adventitious root primordia, because they stimulate cell division; for this reason the accumulation of auxins above and below the girdling zone favors the formation of roots and callus in this specific area (Taiz and Zeiger, 2006). The callus in the girdling zone of the branches was observed as a roundish mass of a yellowish-white color, composed of presumably undifferentiated cells (Fig.3). According to Hartmann et al. (2010) and Colodi et al. (2008), the callus cells could further differentiate and form root primordia or serve as a protective layer for the formation of adventitious roots from other tissues of the layered branch.

In most plants, callus formation is independent on adventitious root formation, although the simultaneous appearance of these tissues may happen because both are influenced by the same environmental factors (Salisbury and Ross, 1994; Burch and McGraw, 1996; Acosta et al., 2000; Colodi et al., 2008). The plants of the Ericales order differ in this feature and can form adventitious roots directly from callus, such as tea (Camellia sinensis L.) plants (Koyuncu and Balta, 2004), or from cambium and phloem, such as Vaccinium sp. plants (Mahlstede and Watson, 1952). However, in order to know whether the formation of callus is independent on adventitious root formation in M. rupestris, a histological study and analysis of the anatomical changes that occur during the rooting of this species are needed.
During the period of evaluation, the adventitious roots were translucent in appearance, but in one specific case, reddish roots were observed growing out of the black plastic; this might possibly be due to the production of anthocyanins in the adventitious roots exposed to light, which are the pigments that play an important role in photoprotection (Taiz and Zeiger, 2006).
Variables of both fresh and dry weight of the roots and callus behaved similarly to the variable of number of roots, presenting the lowest values at 0 mg L-1, the highest values at 500 and 1,000 mg L-1, and decreased values at 2,000 mg L-1 IBA. For the treatments of 500 and 1,000 mg L-1 IBA, a dry weight of 0.14 and 0.19 g for the roots and 0.30 and 0.28 g for the callus, respectively, and a fresh weight of 1.48 and 2.15 g for the roots and 2.62 and 2.70 g for the callus, respectively, were recorded (Fig.5).
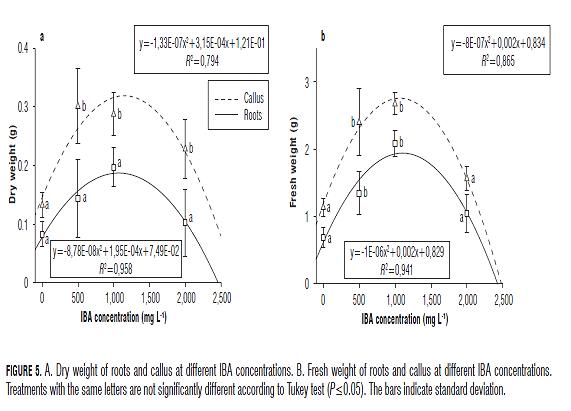
In general, applications of 500 and 1,000 mg L-1 IBA resulted in massive cell growth (callus + roots) in the cut area. According to Hartmann et al. (2010), callus formation is an event that precedes rhizogenesis and, therefore, it is assumed that an increase in callus weight should be reflected in an increase of root weight (Fig.5). However, the histological origin of adventitious root primordia in M. rupestris is unknown, and there is a lack of knowledge on whether callus formation is a prerequisite for the formation of the roots. Comparing these variables, the dry and fresh weights of the callus were high in all IBA treatments, presenting statistically significant differences between the IBA treatments and control (Fig.5).
As for the root dry weight, no statistically significant differences between the treatments were found (Fig.5A), however, when comparing the fresh weight of roots, the differences were statistically significant between the treatments of 500 and 1,000 mg L-1 IBA and the control.
According to Sánchez et al. (2009), in air layers performed on guava (Psidium guajava) plants, with increasing concentrations of IBA (2,000, 4,000, and 6,000 mg L-1), callus formation was reduced, and its formation was completely inhibited at the concentration of 6,000 mg L-1 IBA. A similar trend was obtained in our study for M. rupestris, where concentrations higher than 1,500 mg L-1 IBA resulted in a reduction of dry and fresh weights of the roots and callus. This might be explained by the findings of Moreira et al. (2009), which show that IBA in air layers during periods of active growth (high endogenous auxin levels) could raise hormone levels above the optimal concentrations, leading to a decrease in rooting. Likewise, although IBA increases the elasticity of cell walls, accelerating division, excessive concentrations of the hormone may inhibit this process (Rahman et al., 2000). In the case of propagation by layering in the avocado (Persea sp.), the application of IBA increased the rooting rate, with a concentration of 10,000 mg L-1 being the most efficient, obtaining 74% rooting and presenting the highest number of roots by layer compared to other treatments (Alves-de Oliveira et al., 1999).
According to our findings, one should properly choose the dose of growth regulators because the optimal concentrations vary within the studied species (Kozlowski et al., 1991); for propagation of M. rupestris with air layering, the concentration of 1,200 mg L-1 IBA produced the highest dry and fresh root weights.
Growth of layered branches
The average growth rate of the layered branches was 1.49 cm per month, while the branches without layering presented a growth of 2.78 cm per month (Fig.6). Therefore, the layering adversely affected the longitudinal growth of the branches due to phloem interruption caused by girdling, and the application of exogenous auxins did not completely counteract this effect.

According to Das et al. (1997), during the formation of root primordia in air layers, a gradual consumption of monosaccharides is generated. In turn, the stage of root development decreases the level of nitrogen in the branches (Das et al., 1997). The cicatrization of the wound caused by cutting, followed by consumption of photoassimilates and nutrients for the formation of callus and adventitious roots, might be the cause of the decrease in growth of layered branches (Fig.6), which could affect future flowering and fruiting in the same.
In comparing the longitudinal growth of branches treated with IBA, it may be observed that at concentrations of 1,000 and 2,000 mg L-1 IBA, the branches had the highest growth rates, with values of 1.9 and 2.1 cm per month, respectively. These two treatments had statistically significant differences when compared with the treatments of 0 and 500 mg L-1 IBA which accounted for 1.1 and 1.0 cm per month growth rates, respectively (Fig.6).
The constant contribution of endogenous auxins to subapical regions of the stem is required for the elongation of their cells; additionally the exogenous auxins in cut sections of the stem stimulate their growth (Taiz and Zeiger, 2006). This could help to explain why the treatments with higher IBA concentrations had the highest longitudinal growth rates in layered branches of M. rupestris. Presumably, a branch with a larger number of nodes and leaves in active growth would bring more endogenous auxins to the area of girdling; however, this variable was not evaluated in our study because all branches had a similar size at the time of layering.
For the diameter growth rate (thickness) in the branches, no significant differences were found between the layered and intact branches; also, no effect of the IBA application on this variable was found. Both the thickness and degree of lignification of the branch as well as the rate of photosynthesis in the branch could be indirect indicators of the amount of carbohydrates available for the formation of callus and adventitious roots, therefore it would be important to assess the effect of these variables on the layering of M. rupestris in future studies.
Conclusions and recommendations
The application of exogenous auxins, such as indolebutyric acid, to stimulate the formation of adventitious roots on the layered branches of M. rupestris is feasible due to the fact that the treatments with auxins promoted rooting as compared with the control. A study on the effect of the treatments on post-transplant survival of plants, vegetative growth rate and production is recommended. The results show that a concentration of approximately 1,200 mg L-1 IBA resulted in the largest number of roots and the highest dry weight of roots, and could be considered as the optimal concentration for propagation of M. rupestris with air layering. The propagation of M. rupestris with air layers could be an effective method for consistently obtaining plant material, either for the purposes of production or ecological restoration of páramo and subpáramo areas in Colombia.
Acknowledgements
The authors are grateful to the staff of the Jose Celestino Mutis Botanical Garden, Bogotá, for allowing this study on Macleania rupestris plants belonging to the páramo collection, particularly to Mr. Ricardo Pacheco, Coordinator of the Sustainable Use Project, Scientific Office.
Literature cited
Acero, L. and H. Bernal. 2003. Guía para el cultivo, aprovechamiento y conservación de la uva camarona: Macleania rupestris (H.B.K) A.C. Convenio Andrés Bello y Área de Ciencia y Tecnología No. 127. Bogota.
Acosta, E., M. Sánchez, and M. Bañón. 2000. Auxinas. pp. 305- 323. In: Azcón-Bieto, J. and M. Talón (eds.). Fundamentos de fisiología vegetal. 2nd ed. McGraw-Hill. Interamericana de España, Barcelona, Spain.
Adriance, G. and F. Brison. 1955. Propagation of horticultural plants. 2nd ed. McGraw-Hill, New York, NY .
Aguilar, M. and S. Torres. 2010. Protocolo de uso y aprovechamiento de la uva de anís, Cavendishia bracteata (Ruiz y Pavón ex Jaume Saint. Hillaire) Hoerold, en matorrales andinos del Altiplano Cundiboyacense. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt; MADR; Cámara de Comercio de Bogotá, Bogota.
Alves-de Oliveira, A., O. Carlos-Koller, and A. Villegas-Monter. 1999. Propagación vegetativa de aguacate selección 153 (Persea sp.) por acodo en contenedor. Rev. Chapingo, Serie Hort. 5, 221-225.
Amaya, N. and H. Rincón. 1989. Contribución al estudio de la uva camarona (Macleania rupestris H.B.K.) y su posible utilización en la industria alimenticia. Undergradute thesis. Department of Chemistry, Universidad Nacional de Colombia, Bogota.
Bacarín, M., M. Benincasa, V. Andrade, and F. Fereira. 1994. Enraizamento de estacas aéreas de goiabeira (Psidium guajava L.): efeito do ácido indolbutírico sobre a iniciação radicular. Rev. Cient. 22, 71-79.
Burch, L. and B. McGraw. 1996. Citoquininas. pp. 319-326. In: Azcón-Bieto, J. and M. Talón (eds.). Fundamentos de fisiología vegetal. McGraw-Hill Interamericana de España. Barcelona, Spain.
Cardozo, R., S. Córdoba, J. González, J. Guzmán, H. Lancheros, L. Mesa, R. Pacheco, B. Pérez, F. Ramos, M. Torres, and P. Zúñiga. 2009. Especies útiles en la Región Andina de Colombia. Tomo I. Jardín Botánico de Bogotá José Celestino Mutis, Bogota.
Colodi, F., N. Ramos, K. Zuffellato-Ribas, L. Ribas, and H. Koehler. 2008. Propagação vegetativa de guaco com adição de ácido naftalenoacético. Scient. Agrar. 9(1), 95-98.
Das, P., U. Basak, and A. Das. 1997. Metabolic changes during rooting in pre-girdled stem cuttings and air-layers of Heritiera. Bot. Bull. Acad. Sin. 38, 91-95.
De Valencia, M. and N. De Carrillo. 1991. Anatomía del fruto de Macleania rupestris (H.B.K.) A. C. Smith (uva camarona). Agron. Colomb. 8(1), 286-305.
Dhillon, R., M. Hooda, J. Pundeer, K. Ahlawat, and I. Chopra. 2011. Effects of auxins and thiamine on the efficacy of techniques of clonal propagation in Jatropha curcas L. Biomass Bioenerg. 35, 1502-1510.
Font Quer, A. 1965. Especes fruitieres D' Amérique Tropicale. Fruits 29(6), 474-478.
Hackett, W.P. 1988. Donor plant maturation and adventitious root formation. pp. 11-28. In: Davis, T.D., B.E. Hassing, and N. Sankhla (eds.). Adventitious root formation in cuttings. Advances in Plant Sciences Series. Dioscorides Press, Portland, OR.
Hartmann, H., D. Kester, F. Davies, and R. Geneve. 2010. Plant propagation: principles and Practices. 8th ed. Prentice Hall, Upper Saddle River, NJ .
Henderson, A., S. Clhurchill, and J. Luteyn. 1991. Neotropical plant diversity: are the Northern Andes richer than the Amazon Basin. Nature 351, 21-22.
Koyuncu, F. and F. Balta. 2004. Adventitious root formation in leaf-bud cuttings of tea (Camellia sinensis L.). Pakist. J. Bot. 36(4), 763-768.
Kozlowski, T., P. Kramer, and S. Pallardy. 1991. The physiological ecology of woody plants. Academic. Press, San Diego, CA.
Lagos-Burbano, T.C., H. Ordoñez-Jurado, H. Criollo-Escobar, S. Burbano, and Y. Martínez. 2010. Descripción de frutales nativos de la familia Ericaceae en el altiplano de Pasto, Colombia. Rev. Colomb. Cienc. Hortic. 4, 9-18.
Lambe, A., R. Nuñez-Elisea, and T. Davenport. 1990. New developments in mango marcotting. Tropic. Fruit World. 1, 80-82.
Ligarreto, G. 2011. Agraz (Vaccinium meridionale Swartz.): algunas prácticas de cultivo y poscosecha. Centro Editorial, Facultad de Agronomía, Universidad Nacional de Colombia, Bogota.
Luteyn, J. 2002. Diversity, adaptation and endemism in neotropical ericaceae: biogeographical patterns in the Vaccinieae. Bot. Rev. 68(1), 55-87.
Luteyn, J. and P. Pedraza-Peñalosa. 2008. Neotropical blueberries: the plant family Ericaceae. In: The New York Botanical Garden, http://www.nybg.org/bsci/res/lut2/; consulted: May, 2013.
Luteyn, J. and M. Vidal. (En prensa). Flora de la Real Expedición Botánica del Nuevo Reino de Granada (1783-1816). Tomo XX - Ericaceae. Ediciones Cultura Hispánica, Madrid.
Mahlstede, J. and D. Watson. 1952. An anatomical study of adventitious root development in stems of Vaccinium corymbosum. Bot. Gazette 113(3), 279-285.
Moreira, O., J. Martins, L. Silva, and M. Moura. 2009. Propagation of the endangered Azorean cherry Prunus azorica using stem cuttings and air layering. Life Mar. Sci. 26, 9-14.
National Research Council. 1989. Lost crops of the incas: littleknown plants of the Andes with promise for worldwide cultivation. 2nd ed. Natl. Acad. Press, Washington.
Naz, S. and M. Aslam. 2003. Effect of different concentrations of indole butyric acid (IBA) and age of shoot on air layering of mango (Mangifera indica Linn.). J. Res. Sci. 14(2), 193-198.
Paul, R. and C. Aditi. 2009. IBA and NAA of 1000 ppm induce more improved rooting characters in air-layers of water apple (Syzygium javanica l.). Bulg. J. Agric. Sci. 15(2), 123-128.
Rahman, N., I. Hussain, M. Imran, T. Jan, and A. Awan. 2000. Effect of different concentrations of IBA on rooting of litchi (Litchi chinensis) in air layering. Pakist. J. Biol. Sci. 3(2), 330-331.
Ramírez-Villalobos, M. and A. Urdaneta-Fernández. 2004. Efecto del ácido naftalenacético y de diferentes sustratos sobre el enraizamiento de acodos aéreos del guayabo (Psidium guajava L.). Rev. Fac. Nac. Agron. Medellin 21(1), 28-34.
Rodríguez, J. and J. Peña. 1984. Flora de los Andes. Impresión Escala, Bogota.
Salisbury, F. and C. Ross. 1994. Fisiología vegetal. 4th ed. Grupo Editorial Iberoamérica, Mexico DF.
Sánchez, A., E. Suárez, M. González, Y. Amaya, C. Colmenares, and J. Ortega. 2009. Efecto del ácido indolbutírico sobre el enraizamiento de acodos aéreos de guayabo (Psidium guajava L.) en el municipio Baralt, Venezuela. Evaluación preliminar. Rev. Cient. UDO Agríc. 9(1), 113-120.
Sanjinés, A., B. Øllgaard, and H. Balslev. 2006. Frutos comestibles. pp. 329-346. In: Moraes, M., B. Øllgaard, L. Kvist, F. Borchsenius, and H. Balslev (eds.). Botánica económica de los Andes Centrales. Universidad Mayor de San Andrés, La Paz.
Snow, D. 1981. Tropical frugivorous birds and their food plants: a world survey. Biotropica 12(1), 1-14.
Sturm, H. and J. Rangel. 1985. Ecología de los páramos Andinos. Editora Guadalupe. Bogota.
Taiz, L. and E. Zeiger. 2006. Fisiología vegetal. Volumen II . 3rd ed. Publicacions de la Universitat Jaume I, Castelló de la Plana, Spain.
Tchoundjeu, Z., M. Avana, R. Leakey, A. Simons, E. Asaah, B. Duguma, and J. Bell. 2002. Vegetative propagation of Prunus africana: effects of rooting medium, auxin concentrations and leaf area. Agrofor. Systems. 54, 183-192.
Umba, A., M. Parada, C. Ortiz, I. Alarcón, and L. Rosero. 2007. Dispersión de semillas por aves frugívoras en dos formaciones vegetales del Parque Natural Municipal Ranchería "PN MR". Faculty of Sciences, Universidad Pedagógica y Tecnológica de Colombia (UP TC), Paipa, Colombia.
Vásquez, R. and G. Coímbra. 2002. Frutas silvestres comestibles de Santa Cruz. 2nd ed. Fundación Amigos de la Naturaleza. Santa Cruz, Bolivia.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
Article abstract page views
Downloads
License
Copyright (c) 2013 Agronomía Colombiana

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
© Centro Editorial de la Facultad de Ciencias Agrarias, Universidad Nacional de Colombia
Reproduction and quotation of material appearing in the journal is authorized provided the following are explicitly indicated: journal name, author(s) name, year, volume, issue and pages of the source. The ideas and observations recorded by the authors are their own and do not necessarily represent the views and policies of the Universidad Nacional de Colombia. Mention of products or commercial firms in the journal does not constitute a recommendation or endorsement on the part of the Universidad Nacional de Colombia; furthermore, the use of such products should comply with the product label recommendations.
The Creative Commons license used by Agronomia Colombiana journal is: Attribution - NonCommercial - ShareAlike (by-nc-sa)

Agronomia Colombiana by Centro Editorial of Facultad de Ciencias Agrarias, Universidad Nacional de Colombia is licensed under a Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional License.
Creado a partir de la obra en http://revistas.unal.edu.co/index.php/agrocol/.
















