



Published
The effect of high iron doses (Fe2+) on the growth of broccoli plants (Brassica oleracea var. Italica)
Efecto de dosis altas de hierro (Fe2+) sobre el crecimiento de plantas de brócoli (Brassica oleracea var. Italica)
DOI:
https://doi.org/10.15446/agron.colomb.v32n1.42060Keywords:
plant nutrition, dry mass partitioning, iron sulfate, toxicity to plants (en)nutrición de los cultivos, distribución de la materia seca, sulfato de hierro, toxicidad de las plantas (es)
Downloads
CROP PHYSIOLOGY
1 Plant Ecophysiology Research Group, Faculty of Agricultural Sciences, Universidad Pedagogica y Tecnologica de Colombia. Tunja (Colombia). jaime.pena@uptc.edu.co
Received for publication: 14 January, 2014. Accepted for publication: 19 March, 2014.
ABSTRACT
Tests were carried out under greenhouse conditions in Tunja (Colombia) in order to evaluate the effect of Fe2+ toxicity on the growth of broccoli plants. 'Legacy' hybrid Brassica oleracea var. Italica plantlets were grown in glass containers with a nutritive solution. Iron sulfate was added to the substrate in order to produce excess iron at concentrations of 100 and 200 mg L-1; a control without iron sulfate applications was used. The following evaluations were made: leaf area, total dry weight of the plants, distribution of dry mass (DM) in the different organs, absolute growth rate, relative growth rate, net assimilation rate and the root:shoot ratio. The total DM decreased drastically in the plants subjected to excess Fe2+, the growth indices progressively decreased with increases in the Fe2+ concentrations in the substrate and the distribution of DM in the organs varied as a function of the needs of the plants, with 15.85 and 11.10% less DM in the roots of the plants subjected to Fe2+ than in the control plants, at 100 and 250 mg L-1, respectively.
Key words: plant nutrition, dry mass partitioning, iron sulfate, toxicity to plants.
RESUMEN
Con el objetivo de evaluar el efecto de la toxicidad por Fe2+ sobre el crecimiento de plantas de brócoli, se llevó a cabo un ensayo bajo condiciones de invernadero en Tunja (Colombia). Se utilizaron plántulas de Brassica oleracea var. Italica híbrido 'Legacy', sembradas en contenedores de vidrio con solución nutritiva. Se adicionó sulfato de hierro al sustrato para inducir el exceso del metal, en concentraciones de 100 y 250 mg L-1 y un control sin aplicación de sulfato de hierro. Se evaluó el área foliar, el peso seco total de la planta y la distribución de la materia seca (MS) en los diferentes órganos, se calculó la tasa de crecimiento absoluto, tasa de crecimiento relativo, tasa de asimilación neta y la relación raíz:vástago. El peso seco total disminuyó drásticamente en las plantas sometidas a exceso de Fe2+, los índices de crecimiento disminuyeron progresivamente a medida que aumentó la concentración de Fe2+ en el sustrato y la distribución de la MS en los órganos varió en función de las necesidades de la planta, siendo 15,85 y 11,10% menor la proporción de MS en las raíces de las plantas sometidas a toxicidad por Fe2+ que en las plantas control, para 100 y 250 mg L-1, respectivamente.
Palabras clave: nutrición de los cultivos, distribución de la materia seca, sulfato de hierro, toxicidad de las plantas.
Introduction
Toxicity due to excess iron is a frequent problem in acid soils (Nenova, 2006) and can be caused by microbial action in flooded soils, which results in the reduction of insoluble Fe3+ in favor of soluble Fe2+, which can be taken up by the roots of plants in excessive quantities (Becker and Asch, 2005). In addition, when the content of this metal is high in the soil, the symtomatology seen in affected plants corresponds to deficiencies of other elements considered as antagonistic to iron, such as in the case of Mn (Hanke, 2008), P and ZN, in addition to H2S toxicity conditions (Kirk, 2004).
Currently, the relationship between iron toxicity severity, expressed symptoms and crop yield has not been clearly established (Asch et al., 2005). However, the principal symptom associated with Fe2+ is called browning or yellowing of the leaves, fundamentally characterized in Oryza sativa plants, which causes a reduction in growth, principally in height and tillering, and an increase in sterility of the panicle (Audebert, 2006a). In addition, the physiological behavior of the plants under Fe2+ toxicity conditions has been documented for various vegetative species under laboratory and natural conditions, fundamentally based on parameters such as the accumulation of dry and fresh mass, root and shoot lengths, number of leaves, and leaf area (Nenova, 2006). Similarly, excess iron toxicity in the soil has been related to the behavior of photosynthetic apparatus in broccoli plants, principally based on measurements of the fluorescence of chlorophyll a (Peña-Olmos and Casierra-Posada, 2013).
Taking into account the fact that, in Colombia, the majority of soils have acidity problems (Casierra-Posada et al., 2008) and that excess iron toxicity is a frequent limitation associated with low pH values in the soil (Nenova, 2006). The present study aimed to evaluate the growth of Brassica oleracea var. Italica plants subjected to excess Fe2+ stress.
Materials and methods
The evaluation was developed in a glass greenhouse of the Facultad de Ciencias Agropecuarias of the Universidad Pedagogica y Tecnologica de Colombia - UPTC (Tunja, Colombia), with an average temperature inside the greenhouse of 15.8°C, 72.0% relative humidity and an average photosynthetically active radiation (PAR) of 1,380 µmol m-2 s-1. One-month-old 'Legacy' hybrid broccoli plantlets (Brassica oleracea var. Italica) were used as the initial plant material, sown in glass containers with a nutritive solution of the following composition in mg L-1: 40.3 nitric nitrogen; 4.0 ammonical nitrogen; 20.4 phosphorus; 50.6 potassium; 28.8 calcium; 11.4 magnesium; 1.0 sulfur; 1.12 iron; 0.012 manganese; 0.012 copper; 0.0264 zinc; 0.106 boron; 0.0012 molybdenum and 0.00036 cobalt.
Approximately 20 d after transplant, iron sulfate was added to the plants at concentrations of 100 and 200 mg L-1 in order to induce excess iron; a control without an iron sulfate application was also used. The pH of the solutions was 5.5, 5.3 and 6.2, respectively. The iron additions were done progressively in order to avoid damaging the plants. 44 d after transplant, the leaf area was determined using an LI-3000-A® integrated meter (LI-COR, Lincoln, NE), the DW of the roots, the stems, the leaves and total DW were also determined through oven drying at 105°C until constant weight. In addition, the distribution of the dry material in the different organs was determined as a percentage of the DW assigned to each organ in respect to the total DW of the plant. Based on the DW, the root:shoot ratio was calculated as a coefficient between the root DW and the aerial parts DW. At the begining of the experiment, the initial leaf area and DW were recorded for 20 plants in order to have the initial values needed for the calculation of the growth indices (absolute growth rate (AGR), relative growth rate (RGR) and net assimilation rate [NAR]).
The greenhouse was prepared using tubes and hoses for the connection of an aeration system for the glass containers in order to oxygenate the nutritive solution of the plants. The experimental units were arranged with a completely random design with three treatments and 15 repetitions per treatment. An analysis of variance (ANAVA) was carried out and the treatments were compared using a Tukey mean comparison test (P≤0.05). The statistical analysis was done with version 19.0.0 of IBM-SPSS statistics (IBM Corporation, New York, NY).
Results and discussion
The total DW of the plants was markedly lower in the plants subjected to iron toxicity; the concentrations of 100 and 250 mg L-1 of the metal induced a reduction in the total DM of the plants of 73.63 and 84.81%, respectively, in regard to the control plants (P≤0.05) (Fig. 1).
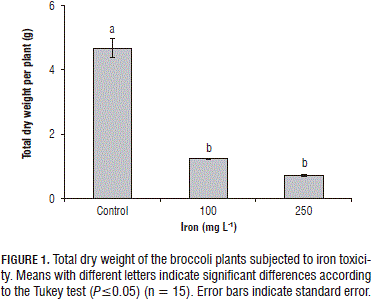
Mehraban et al. (2008) found that elevated concentrations of Fe2+ in the substrate strongly decreased radicle development, supported on the DW, especially when the Fe2+ doses surpassed 50 mg L-1 because high concentrations of the metal increased the peroxidation of lipids, particularly in the radicle zone, which was accompanied by stunted growth of the same. This radicle behavior affected the complete development of the plants because reductions in radicle volume and losses of radicle hairs can induce a drastic decrease in the relative content of water in plants (Dorlodot et al., 2005).
In rice, the DW of roots, veins and foliar lamina did not demonstrate significant differences when the plants were subjected to high iron concentrations in the substrate (Majerus et al., 2007). However, the progressive decrease in the DM in the roots, stems, and leaves in the present study is evidence of impeded growth, produced by a high flow of Fe2+ ions towards the interior of the plant. Becker and Asch (2005) indicated that a strategy used by rice plants to offset excess Fe2+ is the accumulation of the metal in the stems and leaves; highly elevated concentrations of this metal cause early defoliation in these plants in an attempt to attenuate the toxic effects in the plant with a consequent decrease, and in some cases complete cessation of growth in different organs of the plant, as occurred in the present study.
Decreases in the total DW of pea plants were reported by Nenova (2006) at 41 d after sowing and with a Fe2+ concentration of 40 mg L-1 in the substrate. This was due to the fact that the iron toxicity induced the formation of reactive oxygen species, which could be the cause of the growth inhibition. However, Batty and Younger (2003) confirmed that growth inhibition may occur before Fe2+ concentrations in the plant tissues are toxic, which suggests that a strong flow of Fe2+ towards the plant tissues creates a barrier that impedes the entrance of other nutrients that are essential for the growth and development of the plants. For example, in the cultivation of rice, high applications of Fe2+ in the substrate increase the concentrations of the metal in the roots, veins and foliar lamina, which reduce the concentrations of K, Ca, Mg, Mn and P in the shoots of the plants, which affects the growth and development of the crop (Majerus et al., 2007).
The control plants presented the highest foliar area value, followed by the plants subjected to the 100 and 250 mg L-1 Fe2+ concentrations in the nutritive solution, in which the values were reduced by 60.87 and 87.98%, respectively (P≤0.05) (Fig. 2).
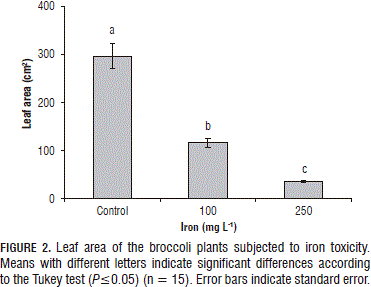
By definition, leaves are reserve organs and Fe2+ is an immobile element, and so is constantly accumulated within the vacuoles of foliar tissue cells and cannot be redistributed during senescence (Audebert, 2006b). The strong decrease in the leaf area of the plants subjected to excess iron in the preset study suggests that the constant flow of the metal through the conductor tissues of the plants was sufficiently strong to cause premature loss of the leaves in the evaluated plants. This may have been due to the low levels of soluble protein, total soluble sugars, chlorophyll a, b and total chlorophyll, as induced by the Fe2+ (Mehraban et al., 2008), which decreased foliar growth and expansion and the useful leaf life, and, consequently, caused a reduction in crop yield (Audebert, 2006b). Similarly, excess iron toxicity induced an increase in the lipids peroxidation of leaves in Eugenia uniflora (De Oliveira-Jucoski et al., 2013), which affected the structure and process of cellular division, drastically decreasing foliar expansion.
The assignment of DM to the different organs presented significant differences between the evaluated treatments (P≤0.05) (Fig. 3). The distribution of photoassimilates towards the leaves increased with the 100 mg L-1 Fe2+ application in the substrate, while the 250 mg L-1 addition produced a higher flow of photosynthates towards the stem and lower development in the leaves and roots, respectively.
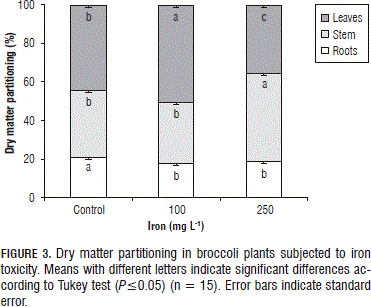
The reduction of the DM percentage destined for the roots in the plants subjected to iron toxicity in the present study corresponds with the high accumulation of Fe2+ in the plants, which reduced plant growth (Mehraban et al., 2008); however, the chlorophyll contents also decreased as a consequence of oxidative stress (Gajewska and Skłodowska, 2007), in this case, due to the increase of the Fe2+ concentration in the plant tissues (Mehraban et al., 2008), which would limit the photosynthetic activity of the same and also cause a smaller proportion of photoassimilates produced in the leaves and translocated towards the roots for growth and development. Peña-Olmos and Casierra-Posada (2013), who worked with the same plants as the present study, found that the electron transport rate (ETR) in the leaves decreased drastically with an increase in the iron concentration of the substrate, which directly affected the photosynthetic activity of the plants.
The increase in the percentage of DM in the leaves of the plants subjected to 100 mg L-1 Fe2+ could be the result of a compensatory behavior of the plants, due to the drastic decrease of the radicle development and the consequent decrease in the uptake of essential nutrients as a result of the high flow of Fe2+ ions towards the interior of the plants requiring a higher efficiency of the same in the capture of light in order to supplement the nutritional requirements throughout the structure, offsetting the low radicle development and producing an osmotic readjustment in the plant. Peña-Olmos and Casierra-Posada (2013), found in broccoli that the maximum quantum efficiency of photosystem II (Fv/Fm) decreased notably with an increase in the Fe2+ concentration of the substrate, concluding that the stress levels of the plants were elevated, possibly due to the accumulation of Fe2+ ions in plant tissues. Many studies have reported that abiotic stress induces an increase in accumulated sugars in the aerial parts of plants and exercises a strong influence on the partition of assimilates in different plant species (Majerus et al., 2007).
For example, Majerus et al. (2007) found that the concentration of soluble sugars decreased in the roots with the application of Fe2+, reaching 80% less in plants stressed with 500 mg L-1 Fe2+ in comparison with the control plants. However, the same authors indicated that the concentrations of soluble sugars increased with Fe2+ applications both in the veins and in the foliar lamina. This increase in the concentration of soluble sugars in leaves, added with the decrease of same in the roots, suggests that sucrose load and its subsequent translocation from the aerial part towards the leaves are inhibited under conditions of excess Fe2+ stress, especially in susceptible crops. An increase in concentrations of sucrose, glucose and fructose, as a result of a modification in the activities of the sucrose phosphate synthase, sucrose synthase and invertase, was found in Lupinus albus leaves subjected to a water deficit (Pinheiro et al., 2001), which, possibly deals with the protection of cellular structures and/or with the osmotic regulation of plants under conditions of severe stress. This did not occur in the plants under 250 mg L-1 Fe2+, possibly due to fact that the concentration of the metal was so high that it almost completely stopped vegetative growth.
The highest accumulation of DM which was found in the stems of the plants treated with 250 mg L-1 Fe2+ may have possibly been due to the plants having two options in the presence of stress induced by excess iron. The first is to tolerate the elevated concentrations of iron in the foliar tissues and the second is to create a barrier to impede the entrance of excess iron into the vegetative tissues. A tolerance to excess iron in broccoli has yet to have been reported; therefore, it is possible to think that the second option is more likely for the counteraction of iron toxicity. This, due to the fact that plants create a barrier of oxidation at the rhizosphere level in order to decrease the entrance of Fe2+ ions towards the vegetative tissues, was stabilized by the canalization of molecular oxygen through the stem and towards the roots, using gas conducting tissue or aerenchyma (Becker and Asch, 2005); for which, plants under conditions of excessive Fe2+ in the substrate must invest a higher percentage of photoassimilates in the formation of gas conducting tissue in the stem.
Table 1 presents the absolute growth rate (AGR), the relative growth rate (RGR) and the net assimilation rate (NAR) of the evaluated broccoli plants. The AGR presented significant differences between the treatments (P≤0.05) with the 100 and 250 mg L-1 Fe2+ applications producing decreases in this variable of 74.77 and 85.98%, respectively. Similarly, the 100 and 250 mg L-1 Fe2+ applications caused a 28.89 and 42.22% decrease in the RGR, respectively (P≤0.05). The Fe2+ toxicity induced a decrease in the NAR value with the addition of 100 mg L-1 of 49.09 and 40.00% with the application of 250 mg L-1.
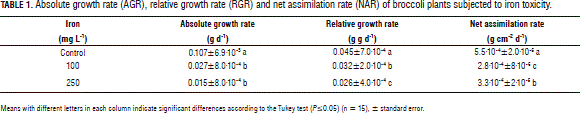
Nenova (2006) found in pea plants that, 34 d after the induction of toxicity with 40 mg L-1 Fe2+, the AGR value decreased 69.12% in comparison with the plants subjected to 2 mg L-1 Fe2+. This was possibly related to a decreased in the electron transport rate, affecting the Calvin cycle and causing a decrease in the carboxylation:oxigenation ratio of Rubisco, which would favor photorespiration (Kampfenkel et al., 1995). Peña-Olmos and Casierra-Posada (2013) found that, in broccoli, the electron transport rate (ETR) decreased notably with an increase in Fe2+ concentration in the substrate. Furthermore, these authors determined the photochemical extinction coefficient (qP) and concluded that the value of this variable is inversely proportional to the Fe2+ content in the substrate, which directly affects the photosynthetic capacity of the plant as well as its growth.
Nenova (2006) reported that the RGR value was reduced 51.95% in plants subjected to applications of 40 mg L-1 Fe2+ 34 d after the induction of the toxicity, in comparison to plants with optimal Fe2+ applications (2 mg L-1). Similarly, Snowden and Wheeler (1993), who worked with 43 plant species native to British swamps, found that high Fe2+ concentrations (10-100 mg L-1) significantly reduced the RGR in all the species except two (Eriophorum angustifolium and Juncus effusus); furthermore, the same authors confirmed that the dicotyledonous plants were more susceptible to Fe2+ toxicity, possibly due to the fact that monocotyledons are generally more porous in the shoots, which facilitates the diffusion of oxygen towards the roots of the plants. Among the dicotyledons, six species reduced their RGR 60% when 10 mg L-1 Fe2+ was added, in comparison with the control plants, showing evidence of sensitivity of the dicotyledonous class to excess Fe2+ stress. Similarly, Oliveira-Jucoski et al. (2013) found that a high accumulation of iron in the plant tissues of Eugenia uniflora, induced by elevated concentrations of the metal in the substrate, produced a reduction in the RGR.
Kampfenkel et al. (1995) confirmed that the rate of photosynthesis in conditions of saturated CO2 in leaves of Nicotiana plumbaginofolia decreased 40% after exposure to excess Fe2+ as a consequence of an increase in foliar Fe2+ content; which in turn caused a 25% decrease in the starch content of the studied leaves. In the present study, the NAR decreased in the plants subjected to excess Fe2+ and this variable is directly proportional to the rate of photosynthesis of the plants. The reduction in the synthesis of sucrose and starch in the leaves exposed to Fe2+ toxicity was accompanied by an inhibition of photosynthesis, an increase in reduced A quinones (QA) and high photochemical extinction coefficient values qN (photochemical losses related to heat, pH gradient and photoinhibition), all under conditions of high radiation and with very few variations with low radiation (Neuhaus et al., 1989; Neuhaus and Stitt, 1991). This increase in the reduced QA was the result of the low pool of the same and as a consequence of increased non-photochemical losses. This induces a low electron transport rate which, together with the ionic stress produced by the excess Fe2+, causes a decrease in the synthesis of photoassimilates in the leaves of plants. However, in addition to a decrease in the rate of photosynthesis as a consequence of excess Fe2+, a increase in the respiration rate of the stressed leaves is produced, which supposes a higher consumption or rupture of hexoses, for which the leaves subjected to high Fe2+ concentrations suffered a strong decrease in the contents of glucose and fructose in comparison with the controls (Kampfenkel et al., 1995). This is supported by studies carried out by Peña-Olmos and Casierra-Posada (2013), who found an increase of initial fluorescence (F0) in the same plants as the present study. This increase was possibly due to damage in the reaction centers of photosystem II (PSII) (Vieira et al., 2010) or to a decrease in the transference of excitation energy from the antenna complex towards the reaction centers (Baker and Rosenqvist, 2004), which may have produced a decrease in the photoinhibition of PSII.
The root:shoot ratio presented significant differences between the treatments (P≤0.05). The addition of 100 and 250 mg L-1 Fe2+ to the substrate in which the broccoli plants grew induced a reduction of 19.29 and 13.72% in the value of this variable with respect to the control plants (Fig. 4).

Snowden and Wheeler (1993) suggested that plants can be classified as tolerant to excess Fe2+ by taking into account the root:shoot ratio. In this way, these authors considered that if Fe2+ applications induced decreases in said ratio with respect to control plants, the plants should be considered sensitive to excess iron due to the fact that the root is more affected than the aerial part of the plant. A higher root:shoot ratio in tolerant plants is possibly due to fact that a high Fe2+ concentration in the substrate causes a higher accumulation of ochre deposits in the roots, which would increase the radicle mass. Similarly, Kampfenkel et al. (1995) suggested that under conditions of excess Fe2+, it is separated and stored in the necrotic points that are developed in the leaves as a consequence of the same stress, as well as in the vacuoles of the mesophyll cells, with which the DW of the leaves would increase in relation to the roots of the plant, producing a decrease in the root:shoot ratio.
Conclusions
The analysis of variance carried out with the data obtained for the different parameters of the plant growth evaluation of the present study allowed for the conclusion that the iron toxicity produced stress in the evaluated plants, which affected the proportion and accumulation of DM in the different plant organs. The reduction in the leaf area expansion and in the evaluated growth indices evidenced a decrease in crop development. And so, it is evident that the exposure of Brassica oleracea var. Italica plants to high Fe2+ levels in the substrate negatively affects the general function of these plants.
Acknowledgements
This study was developed with the support of the Research Directorate (Dirección de Investigaciones - DIN) of the Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja, within the Jóvenes Investigadores UPTC program, and with the support of the Plant Ecophysiology Research Group of the Faculty of Agricultural Sciences of the UPTC.
Literature cited
Asch, F., M. Becker, and D.S. Kpongor. 2005. A quick and efficient screen for resistance to iron toxicity in lowland rice. J. Plant Nutr. Soil Sci. 168, 764-773.
Audebert, A. 2006a. Diagnosis of risk and approaches to iron toxicity management in lowland rice farming. pp. 6-17. In: Audebert, A., L.T. Narteh, P. Kiepe, D. Millar, and B. Beks (eds.). Iron toxicity in rice-based systems in West Africa. Africa Rice Center (WARDA), Cotonou, Benin.
Audebert, A. 2006b. Iron partitioning as a mechanism for iron toxicity tolerance in lowland rice. pp. 34-46. In: Audebert, A., L.T. Narteh, P. Kiepe, D. Millar, and B. Beks (eds.). Iron toxicity in rice-based systems in West Africa. Africa Rice Center (WARDA), Cotonou, Benin.
Baker, N.R. and E. Rosenqvist. 2004. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 55, 1607-1621.
Batty, L.C. and P.L. Younger. 2003. Effects of external iron concentration upon seedling growth and uptake of Fe+2 and phosphate by the common reed, Phragmites australis (Cav.) Trin ex. Steudel. Ann. Bot. 92, 801-806.
Becker, M. and F. Asch. 2005. Iron toxicity in rice - conditions and management concepts. J. Plant Nutr. Soil Sci. 168, 558-573.
Casierra-Posada, F., J.F. Cárdenas-Hernández, and H.A. Roa. 2008. Efecto del aluminio sobre la germinación de semillas de trigo (Triticum aestivum L.) y de maíz (Zea mayz L.). Rev. Orinoquia 12(1), 45-56.
De Oliveira-Jucoski, G., J. Cambraia, C. Riveiro, J. Alves-De Oliveira, S. Oliveira-De Paula, and M.A. Oliva. 2013. Impact of iron toxicity on oxidative metabolism in young Eugenia uniflora L. plants. Acta Physiol. Plant 35(5), 1645-1657.
Dorlodot, S., S. Lutts, and P. Bertin. 2005. Effects of ferrous iron toxicity on the growth and mineral composition of an interspecific rice. J. Plant Nutr. 28(1), 1-20.s
Gajewska, E. and M. Sklodowska. 2007. Relations between tocopherol, chlorophyll and lipid peroxides contents in shoots of Ni-treated wheat. J. Plant Physiol. 164(3), 364-366.
Hanke, F. 2008. La nutrición de la planta y su problemática en la agricultura. Editorial Juan de Castellanos, Tunja, Colombia.
Kampfenkel, K., M. Van Montagu, and D. Inzé. 1995. Effects of iron excess on Nicotiana plumbagnifolia plants, implications to oxidative stress. Plant Physiol. 107, 725-735.
Kirk, G.J.D. 2004. The biogeochemistry of submerged soils. John Wiley & Sons, Chichester, UK.
Majerus, V., P. Bertin, and S. Lutts. 2007. Effects of iron toxicity on osmotic potential, osmolytes and polyamines concentrations in the African rice (Oryza glaberrima Steud.). Plant Sci. 173, 96-105.
Mehraban, P., A.A. Zadeh, and H.R. Sadeghipour. 2008. Iron toxicity in rice (Oryza sativa L.), under different potassium nutrition. Asian J. Plant Sci. 7, 251-259.
Nenova, V. 2006. Effect of iron supply on growth and photosystem II efficiency of pea plants. Gen. Appl. Plant Physiol. (Special Issue) 32, 81-90.
Neuhaus, H.E., A.L. Kruckeberg, R. Feil, and M. Stitt. 1989. Reduced activity mutants of phosphoglucose isomerase in the cytosol and chloroplast of Clarkia xantiana. Planta 178, 110-122.
Neuhaus, H.E. and M. Stitt. 1991. Inhibition of photosynthetic sucrose synthesis by imidodiphosphate, an analog of inorganic pyrophosphate. Plant Sci. 76, 49-55.
Peña-Olmos, J.E. and F. Casierra-Posada. 2013. The photochemical efficiency of photosystem II (PSII) in broccoli plants (Brassica oleracea var Italica) affected by excess of iron. Rev. Orinoquia 17(1), 15-22.
Pinheiro, C., M.M. Chaves, and C.P. Ricardo. 2001. Alterations in carbon and nitrogen metabolism induced by water deficit in the stems and leaves of Lupinus albus L. J. Exp. Bot. 52, 1063-1070.
Snowden, R.E.D. and B.D. Wheeler. 1993. Iron toxicity to fen plant species. J. Ecol. 81, 35-46.
Vieira, D.A.P., T.A. Portes, E. Stacciarini-Seraphin, and J.B. Teixeira. 2010. Fluorescência e teores de clorofilas emabacaxizeiro cv. pérolasubmetido a diferentes concentrações de sulfato de amônio. Rev. Bras. Frutic. 32(2), 360-368.
References
Asch, F., M. Becker, and D.S. Kpongor. 2005. A quick and efficient screen for resistance to iron toxicity in lowland rice. J. Plant Nutr. Soil Sci. 168, 764-773.
Audebert, A. 2006a. Diagnosis of risk and approaches to iron toxicity management in lowland rice farming. pp. 6-17. In: Audebert, A., L.T. Narteh, P. Kiepe, D. Millar, and B. Beks (eds.). Iron toxicity in rice-based systems in West Africa. Africa Rice Center (WARDA), Cotonou, Benin.
Audebert, A. 2006b. Iron partitioning as a mechanism for iron toxicity tolerance in lowland rice. pp. 34-46. In: Audebert, A., L.T. Narteh, P. Kiepe, D. Millar, and B. Beks (eds.). Iron toxicity in rice-based systems in West Africa. Africa Rice Center (WARDA), Cotonou, Benin.
Baker, N.R. and E. Rosenqvist. 2004. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. J. Exp. Bot. 55, 1607-1621.
Batty, L.C. and P.L. Younger. 2003. Effects of external iron concentration upon seedling growth and uptake of Fe+2 and phosphate by the common reed, Phragmites australis (Cav.) Trin ex. Steudel. Ann. Bot. 92, 801-806.
Becker, M. and F. Asch. 2005. Iron toxicity in rice - conditions and management concepts. J. Plant Nutr. Soil Sci. 168, 558-573.
Casierra-Posada, F., J.F. Cárdenas-Hernández, and H.A. Roa. 2008. Efecto del aluminio sobre la germinación de semillas de trigo (Triticum aestivum L.) y de maíz (Zea mayz L.). Rev. Orinoquia 12(1), 45-56.
De Oliveira-Jucoski, G., J. Cambraia, C. Riveiro, J. Alves-De Oliveira, S. Oliveira-De Paula, and M.A. Oliva. 2013. Impact of iron toxicity on oxidative metabolism in young Eugenia uniflora L. plants. Acta Physiol. Plant 35(5), 1645-1657.
Dorlodot, S., S. Lutts, and P. Bertin. 2005. Effects of ferrous iron toxicity on the growth and mineral composition of an interspecific rice. J. Plant Nutr. 28(1), 1-20.s
Gajewska, E. and M. Sklodowska. 2007. Relations between tocopherol, chlorophyll and lipid peroxides contents in shoots of Ni-treated wheat. J. Plant Physiol. 164(3), 364-366.
Hanke, F. 2008. La nutrición de la planta y su problemática en la agricultura. Editorial Juan de Castellanos, Tunja, Colombia.
Kampfenkel, K., M. Van Montagu, and D. Inzé. 1995. Effects of iron excess on Nicotiana plumbagnifolia plants, implications to oxidative stress. Plant Physiol. 107, 725-735.
Kirk, G.J.D. 2004. The biogeochemistry of submerged soils. John Wiley & Sons, Chichester, UK.
Majerus, V., P. Bertin, and S. Lutts. 2007. Effects of iron toxicity on osmotic potential, osmolytes and polyamines concentrations in the African rice (Oryza glaberrima Steud.). Plant Sci. 173, 96-105.
Mehraban, P., A.A. Zadeh, and H.R. Sadeghipour. 2008. Iron toxicity in rice (Oryza sativa L.), under different potassium nutrition. Asian J. Plant Sci. 7, 251-259.
Nenova, V. 2006. Effect of iron supply on growth and photosystem II efficiency of pea plants. Gen. Appl. Plant Physiol. (Special Issue) 32, 81-90.
Neuhaus, H.E., A.L. Kruckeberg, R. Feil, and M. Stitt. 1989. Reduced activity mutants of phosphoglucose isomerase in the cytosol and chloroplast of Clarkia xantiana. Planta 178, 110-122.
Neuhaus, H.E. and M. Stitt. 1991. Inhibition of photosynthetic sucrose synthesis by imidodiphosphate, an analog of inorganic pyrophosphate. Plant Sci. 76, 49-55.
Peña-Olmos, J.E. and F. Casierra-Posada. 2013. The photochemical efficiency of photosystem II (PSII) in broccoli plants (Brassica oleracea var Italica) affected by excess of iron. Rev. Orinoquia 17(1), 15-22.
Pinheiro, C., M.M. Chaves, and C.P. Ricardo. 2001. Alterations in carbon and nitrogen metabolism induced by water deficit in the stems and leaves of Lupinus albus L. J. Exp. Bot. 52, 1063-1070.
Snowden, R.E.D. and B.D. Wheeler. 1993. Iron toxicity to fen plant species. J. Ecol. 81, 35-46.
Vieira, D.A.P., T.A. Portes, E. Stacciarini-Seraphin, and J.B. Teixeira. 2010. Fluorescência e teores de clorofilas emabacaxizeiro cv. pérolasubmetido a diferentes concentrações de sulfato de amônio. Rev. Bras. Frutic. 32(2), 360-368.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
CrossRef Cited-by
1. sunil kulkarni, Ajaygiri Goswami. (2019). Effect of Excess Fertilizers and Nutrients: A Review on Impact on Plants and Human Population. SSRN Electronic Journal, https://doi.org/10.2139/ssrn.3358171.
2. H. R. Moussa, Mohamed A. Taha, Eldessoky S. Dessoky, Eman Selem. (2023). Exploring the perspectives of irradiated sodium alginate on molecular and physiological parameters of heavy metal stressed Vigna radiata L. plants. Physiology and Molecular Biology of Plants, 29(3), p.447. https://doi.org/10.1007/s12298-023-01286-9.
3. Reginald B. Kogbara, Wubulikasimu Yiming, Srinath R. Iyengar, Osman A.E. Abdalla, Hayel M. Al-Wawi, Udeogu C. Onwusogh, Karim Youssef, Marwa Al-Ansary, Parilakathoottu A. Sunifar, Dhruv Arora. (2020). Effect of gas-to-liquid biosludge on soil properties and alfalfa yields in an arid soil. Journal of Cleaner Production, 250, p.119524. https://doi.org/10.1016/j.jclepro.2019.119524.
4. F. Casierra-Posada, J.A. Camargo-Parra, M.C. Acosta. (2018). Sensitivity of sweet pepper plants (Capsicum annuum L.) to iron excess. European Journal of Horticultural Science, , p.382. https://doi.org/10.17660/eJHS.2018/83.6.6.
5. Viktorija Vaštakaitė-Kairienė, Aušra Brazaitytė, Jurga Miliauskienė, Erik S. Runkle. (2022). Red to Blue Light Ratio and Iron Nutrition Influence Growth, Metabolic Response, and Mineral Nutrients of Spinach Grown Indoors. Sustainability, 14(19), p.12564. https://doi.org/10.3390/su141912564.
6. F. Casierra-Posada, J. D. Cortés-Bayona, J. Cutler. (2017). Effect of Iron Excess on Growth of Sisal Plants (Furcraea hexapetala). Gesunde Pflanzen, 69(3), p.123. https://doi.org/10.1007/s10343-017-0396-6.
7. Noreen Zahra, Muhammad Bilal Hafeez, Kanval Shaukat, Abdul Wahid, Mirza Hasanuzzaman. (2021). Fe toxicity in plants: Impacts and remediation. Physiologia Plantarum, https://doi.org/10.1111/ppl.13361.
Dimensions
PlumX
Article abstract page views
Downloads
License
Copyright (c) 2014 Agronomía Colombiana

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
© Centro Editorial de la Facultad de Ciencias Agrarias, Universidad Nacional de Colombia
Reproduction and quotation of material appearing in the journal is authorized provided the following are explicitly indicated: journal name, author(s) name, year, volume, issue and pages of the source. The ideas and observations recorded by the authors are their own and do not necessarily represent the views and policies of the Universidad Nacional de Colombia. Mention of products or commercial firms in the journal does not constitute a recommendation or endorsement on the part of the Universidad Nacional de Colombia; furthermore, the use of such products should comply with the product label recommendations.
The Creative Commons license used by Agronomia Colombiana journal is: Attribution - NonCommercial - ShareAlike (by-nc-sa)

Agronomia Colombiana by Centro Editorial of Facultad de Ciencias Agrarias, Universidad Nacional de Colombia is licensed under a Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional License.
Creado a partir de la obra en http://revistas.unal.edu.co/index.php/agrocol/.
















