


Publicado
Deforestación y sedimentación en los manglares del Golfo de Urabá Síntesis de los impactos sobre la fauna macrobéntica e íctica en el delta del río Turbo
Palabras clave:
hidroclimatología, deforestación, sedimentación, gasterópodos, ictiofauna, comunidades costeras. (es)Deforestación y sedimentación en los manglares del Golfo de Urabá: Síntesis de los impactos sobre la fauna macrobéntica eíctica en el delta del río Turbo
Deforestation and sedimentation in Uraba Gulf mangroves A synthesis of the impacts on macrobenthos and fishes in the Turbo River Delta
Juan Felipe Blanco-Libreros1 Alexander Taborda-Marín2 Viviana Amortegui-Torres3 Andrea Arroyave-Rincón4 Alejandro Sandoval5 Edgar Andrés Estrada6 Jenny Leal-Flórez7 Jairo Guillermo Vásquez Arango8 Alberto Vivas Narváez9
1. Doctor en Ciencias-Biología,ELICE-Universidad de Antioquia. blanco@exactas.udea.edu.co
2. M.Sc. en Ciencias del Mar, ELICE-Universidad de Antioquia
3. Bióloga, ELICE-Universidad de Antioquia
4. Bióloga ELICE-Universidad de Antioquia
5. Biólogo Marino, ELICE Universidad de Antioquia
6. Ecólogo de Zonas Costeras, ELICE-Universidad de Antioquia
7. Doctora en Ciencias Naturales, GISMAC-Universidad de Antioquia
8. Biólogo Marino, CORPOURABA
9. Ingeniero Forestal, CORPOURABA
Recibido para evaluación: 4 de septiembre de 2012, Aceptación: 22 de febrero de 2013, Recibido versión final: 29 de mayo de 2013
RESUMEN
Se presenta una síntesis de la investigación que relaciona la deforestación y el cambio del uso del suelo de la planicie costera de la cuenca del río Turbo con los efectos sobre la fauna estuarina de su delta. Se contextualiza esta cuenca dentro la variabilidad hidroclimatológica (caudales y precipitación) del costado oriental del golfo de Urabá. Se cuantificó la tasa de pérdida del bosque de la planicie costera debido al aumento de cultivos, lo cual puede explicar el incremento del transporte de sedimentos hacia los ríos y al mar. A pesar de la expansión deltaica, elárea de manglares disminuyó como producto del aumento de cultivos y potreros. Se documentó que la disminución de la densidad de un gasterópodo dominante (Neritina virginea) en claros del manglar, bordes y potreros, por alteraciones en su microhábitat, puede ser utilizada como un bioindicador. Las altas tasas de sedimentación podrían ser responsables de la pobreza faunística del bentos, pero no parecen tener un efecto significativo sobre las especies actualmente dominantes. La diversidad y abundancia de peces reflejó una mayor influencia de la sedimentación en las zonas más cercanas a la descarga fluvial. Se relacionó la condición social de las comunidades con los cambios en el paisaje. Se estableció el estado actual del ecosistema, que será la base para proponer medidas de manejo y conservación para prevenir y restaurar los daños causados a los manglares y a la zona costera.
Palabras clave: hidroclimatología, deforestación, sedimentación, gasterópodos, ictiofauna, comunidades costeras.
ABSTRACT
This synthesis relates deforestation and land use change in coastal plain of the Turbo River watershed and impacts upon estuarine fauna in its delta. This watershed is shown within the context of hidroclimatological (rainfall and discharge) variability across the eastern margin of Urabá Gulf. Coastal-plain forest conversion rate to crops were quantified, as a possible explanation of increased sediment transport to the river and the nearshore. Despite of the expansion of the delta, mangrove area was reduced as a consequence of conversion to crops and pastures. The dominant mangrove snail Neritina virginea was reduced in density in anthropogenic forest gaps and edges, as well as in pastures, due to altered microhabitats, and can be therefore used as a bio-indicator. The high sedimentation rates seem to be responsible for the faunistic poverty of the benthos, but do not seem responsible of deleterious effects on the dominant species. The diversity and abundance of fi shes was greatly altered by high sedimentation near the river mouth. Finally, social features of the human communities were related to landscape changes. Herewith, we reported on the current ecosystem status, as the baseline for proposing management and conservation guidelines in order to prevent and restore impacts on mangroves and the coastal zone in this region.
Key words: Hydroclimatology, deforestation, sedimentation, gastropods, fi shes, human coastal communities.
1. INTRODUCCIÓN
Los manglares son uno de los ecosistemas costeros más productivos y biodiversos en la tierra, proporcionando múltiples beneficios y jugando un papel vital en la sostenibilidad de los trópicos (Hogarth, 2007). Sin embargo, su existencia se encuentra en grave peligro debido al cambio de cobertura y uso del suelo hacia potreros y cultivos en las cuencas costeras, lo cual ha provocado una disminución de la extensión de los bosques inundables, y un aumento de la exportación de sedimentos hacia el mar (Valiela et al, 2001; Restrepo, 2005). Dicha problemática no es ajena a las costas del golfo de Urabá, las cuales muestran signos evidentes de erosión y sedimentación que podrían estar ligados a actividades antrópicas en las cuencas hidrográficas costeras (Correa y Vernette, 2004; Bernal et al., 2005; García-Valencia, 2007; Blanco, 2009).
Actualmente, en la región de Urabá, el cultivo de banano y plátano, y la ganadería, son las principales actividades agropecuarias en términos de extensión y contribución a la economía regional. García-Valencia et al. (2007), estimaron que, para toda región de Urabá, elárea cultivada cubría 62.235 ha (6,1%) y elárea de pastos 86.160ha (8,4%). Dentro de este contexto, la cuenca hidrográfica del río Turbo puede ser utilizada como modelo de estudio de la deforestación de las laderas y su influencia sobre el aumento en los sedimentos exportados por las corrientes de agua y su efecto sobre la fauna estuarina, ya que muchos de los tensores descritos para toda la región de Urabá ocurren de manera notoria en ella. Por otra parte, el río Turbo es el principal abastecimiento de agua para consumo humano en el casco urbano del municipio del mismo nombre, recibe descargas puntuales y difusas de aguas residuales domésticas y agropecuarias y, a lo largo de su cuenca hidrográfica, la población ha aumentado en las últimas décadas trayendo consigo la sobre-explotación de los recursos naturales, en especial los forestales (Keep, 2000; CORPOURABA, 2007). El traslado de la desembocadura a mediados del siglo XX y posiblemente el aumento de la deforestación de la cuenca y la sedimentación costera han contribuido a la rápida formación del nuevo delta del río Turbo, el cual está conformado por una planicie y una espiga dirigida al sur, denominada punta Yarumal, y por una laguna costera que se ha cerrado progresivamente, denominada bahía El Uno (Estrada y Gil, 2005; Taborda, 2008; Blanco, 2009).
Debido a que la deforestación de las cuencas es controlada por dinámicas humanas sociales, culturales y económicas, es importante comprender la forma en que los habitantes de la cuenca visualizan su territorio, ya que permite incluir elementos que enriquecen la interpretación biofísica, y también permiten evidenciar hasta dónde la situación ambiental obedece a prácticas locales o a otras fuerzas externas (Escobar, 1999). En el caso del municipio de Turbo es claro que los problemas a los que se enfrentan los habitantes de la zona rural provienen en gran medida de la concentración de la propiedad de la tierra y concentración de la población urbana, de la ilegalidad en la tenencia de la tierra y de los bajos niveles de desarrollo del capital humano (Keep, 2000).
Estas complejas problemáticas socio-económicas se pueden integrar conceptualmente alrededor de los procesos de deforestación-erosión-sedimentación, tal como se ha planteado dentro de los conceptos de sistemas ecológicos y sociales acoplados (Collins et al., 2011) y de cambio del uso de la tierra (Turner y Robbins, 2008) que se han aplicado principalmente a sistemas terrestres. Mediante estos conceptos se puede vincular la deforestación de bosques al crecimiento poblacional, a la expansión de actividades agropecuarias, y a la extracción de productos maderables, tal como lo demuestran muchos estudios ecológicos (Etter et al., 2006a; FAO, 2011), pero a su vez se pueden explorar las causas de dichos procesos humanos mediante el uso de las etno-ciencias (Rist y Dahdoh-Guebas, 2006). Con este cambio de paradigma se puede lograr no solo una comprensión más integral o multidisciplinar de los problemas, sino también aportar elementos entre disciplinas (Rist y Dahdoh-Guebas, 2006; Dahdouh-Guebas y Koehdam, 2008). La situación de la transformación del paisaje costero del Caribe colombiano requiere una mejor comprensión de las causas antropogénicas y de los efectos, no solo en elámbito físico sino ecológico, lo cual se beneficiaría de la aplicación de una aproximación inter- y trans-disciplinaria, que además aborde múltiples escalas espaciales y temporales (sensu Dahdouh-Guebas y Koehdam, 2008). Por ejemplo, en la planicie costera de la región Caribe, el aumento de los asentamientos humanos ha provocado un incremento delárea cultivada, la cual era de 1’083.044ha (22% delárea nacional) en 1996 (Etter et al., 2006b), y parte de dicha transformación es responsable no solo de la tala de los bosques secos y humedales, sino de los manglares (álvarez-León y Polanía, 1996).
En otras partes del mundo, la deforestación de las planicies costeras también ha tenido un efecto significativo en la cantidad de sedimentos que son exportados por las cuencas hidrográficas hacia el mar (Wang et al., 2005; Patrick et al., 2006), originando serios daños en litorales, ecosistemas estuarinos, arrecifes de corales y manglares, y afectando gravemente la fauna y flora asociadas (Warne et al., 2005; Wall et al., 2008; Larsen y Webb, 2009). En el Caribe colombiano, Restrepo (2005) ha documentado ampliamente el efecto de la deforestación de la cuenca del río Magdalena sobre el aumento de la producción y carga de sedimentos de los tributarios y su sedimentación sobre los arrecifes coralinos de las islas del Rosario, lo que trae consigo el deterioro de todo el ecosistema; pero infortunadamente no existen estudios similares en otras cuencas del país.
En el golfo de Urabá, los trabajos sobre los cambios de la línea de costa se concentran en el problema de la erosión, pero poco se ha estudiado la sedimentación más allá de cuantificar la progradación (Correa y Vernette 2004; Bernal et al., 2005; Rangel-Buitrago, 2009). También es escasa la información sobre la fauna de los manglares (e.g. Ortiz y Blanco, 2012, Blanco y Castaño 2012, y referencias allí descritas), aunque recientemente, se llevó a cabo una expedición científica para levantar dichos inventarios (Blanco et al., 2011). Esta falta de información, combinada con la diversidad y magnitud de amenazas naturales y antropogénicas es preocupante en el golfo de Urabá (e.g. Blanco et al., 2012), ya que, al igual que en todo el mundo, la fauna béntica de los manglares es de gran importancia debido a su alta abundancia, diversidad taxonómica y trófica, mientras que la fauna pelágica es, además, fuente principal de alimento y subsistencia para los humanos; todos estos bienes, funciones y servicios se han deteriorado por la perturbación del hábitat natural, el aumento de la sedimentación y de los agroquímicos (Cannicci et al., 2008; Nagelkerken et al., 2008)
Este trabajo sintetiza resultados de estudios realizados entre 2007 y 2011 en el marco de varios proyectos de investigación llevados a cabo con el objetivo de comprender integralmente los problemas de deforestación y sedimentación en la cuenca hidrográfica del río Turbo, incluyendo su zona costera, como modelo de estudio para el golfo de Urabá y el Caribe colombiano sur. Se analizó la variación espacial y temporal de procesos geográficos, hidrológicos, sedimentológicos, físicoquímicos y biológicos (comunidades estuarinas béntica eíctica) relacionados con la sedimentación en los ecosistemas de manglar en el contexto de su cuenca hidrográfica. Específicamente se preguntó: a) ¿Cuál es la magnitud de la producción y carga de sedimentos hacia el mar? ¿Hay evidencias de influencias antropogénicas?, b) ¿Cuál es la magnitud de la deforestación en las cuencas costeras? ¿Cuál es el mecanismo que la vincula con la producción de sedimentos?, c) ¿Cuáles son las magnitudes de la sedimentación en el estuario? y d) ¿Cuál es su efecto sobre el bentos y el necton?
2. MATERIALES Y MÉTODOS
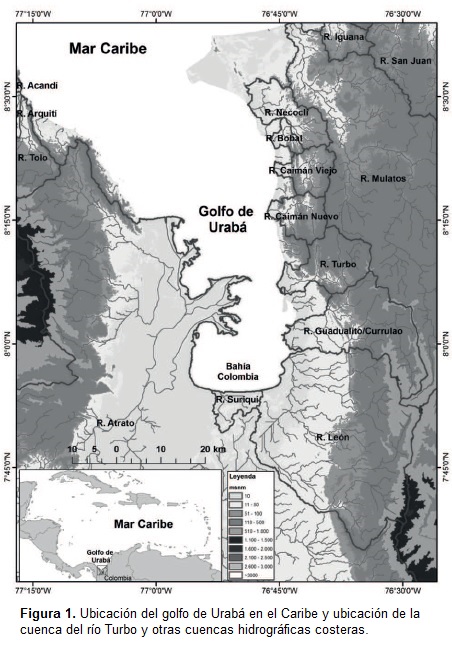
Área de estudio: El golfo de Urabá está localizado en el extremo suroccidental del Caribe colombiano. Territorialmente es compartido por los departamentos de Chocó y Antioquia; este último comprende los municipios de Necoclí, San Juan de Urabá, Arboletes y Turbo (Figura 1) abarcando un área estimada de 4.291Km2 (CORPOURABA, 2003). Presenta climas que varían de tropical seco al noreste a tropical húmedo al oriente y muy húmedo en los otros sectores, con una temperatura media diaria que fluctúa entre 26 y 28°C (CORPOURABá, 2003; Taborda, 2008). En el costado oriental del Golfo, en el municipio de Turbo, se ubica la cuenca del río del mismo nombre. En la desembocadura del río se forma punta Yarumal, la cual confina una laguna costera que recibe el nombre de bahía El Uno, y constituye un delta de 217,9 ha sobre el cual se ha desarrollado un ecosistema de manglar desde la década de los sesentas (Taborda, 2008). A la fecha aún prevalece un borde costero dominado por Rhizophora mangle y rodales interiores de Avicennia germinans y Laguncularia racemosa (CORPOURABA, 2003, 2007; Taborda, 2008). Son el río Turbo y su cuenca baja los principales modelos de estudio en la presente investigación. La cuenca, con una superficie aproximada de 150 km2, se caracteriza por tener una zona de llanuras que va desde la desembocadura en el golfo hasta el pie de monte de la serranía de Abibe (Keep 2000, POT de Turbo 2000b); en total, elárea de interés comprende 905,3 ha correspondientes a la cuenca baja.
Influencias antropogénicas sobre la hidrología y la exportación de sedimentos en el golfo de Urabá: Se recopilaron series de tiempo de caudales (m3/s) y transporte diario de materiales en suspensión (exportación de sedimentos: kton/día) de diez estaciones del Instituto Colombiano de Meteorología e Hidrología (IDEAM, Anuario Hidrológico 1990-1993) en la región de Urabá. Se calcularon las medias mensuales de ambas variables en todas las estaciones y se graficaron para establecer la ocurrencia de patrones estacionales consistentes en el espacio. Se exploró la falta de congruencia del patrón estacional entre el caudal y la exportación de sedimentos, ya que la misma es un indicador de influencias antropogénicas sobre la hidrología de la cuenca, particularmente sobre las fuentes de sedimentos hacia los cuerpos de agua (Restrepo, 2005). Se comparó la magnitud de la exportación de sedimentos del río Turbo con la de otros ríos tributarios de esta costa (ver métodos adicionales en Arroyave 2011; Arroyave et al., 2012).
Tasas de deforestación, exportación y depósito de sedimentos en la cuenca del río Turbo: Para cuantificar la tasa de deforestación en la cuenca baja del río Turbo se analizaron fotografías aéreas e imágenes satelitales que incluyeran elárea de estudio desde 1960 hasta el 2007. Se definieron y clasificaron las categorías de coberturas como manglar, bosque natural, rastrojo alto, rastrojo bajo, cultivos, pastos, suelo desnudo, río Turbo, cuerpos de agua, zona urbana. Con esto se realizó un mapa geo-referenciado de cobertura para cada década, en el que se calculó elárea y porcentaje ocupado para cada tipo de cobertura y se analizaron los cambios entre décadas y, al inicio y final, se calculó el cambio neto medio anual de cada cobertura entre periodos de tiempo por medio de métodos estándares (Etter et al., 2006ª; ver métodos adicionales en Taborda, 2008).
Para establecer la exportación de sedimentos, entre enero y septiembre de 2010 se midió, una vez por mes, la concentración de los sólidos suspendidos totales SST (mg/L) en muestras de agua tomadas a lo largo de la pluma turbia del río Turbo y la zona costera aledaña; dichos datos fueron analizados por el método de gravimetría en el laboratorio de aguas de CORPOURABA. Se realizaron mediciones en siete estaciones de muestreo distribuidas dentro del estuario fluvial (desembocadura), la zona costera (exterior de punta Yarumal) y la laguna costera (bahía El Uno). Para las estaciones situadas en la desembocadura fue calculada la exportación bruta de sedimentos (EBS) o carga de sedimentos (sediment load) con los datos medidos de SST y el aforo de caudal. Por otro lado, mediante el análisis de imágenes satelitales, fotografías aéreas y reconocimiento in situ de la zona fueron establecidas cuatro estaciones, en el borde externo de los manglares del delta, que representaran el gradiente natural para la cuantificación de las tasas de sedimentación en el estuario. En cada sitio se ubicaron nueve trampas (cilindros plásticos de 2,5L fijados al fondo), reemplazadas mensualmente, para secar y pesar los sedimentos.
Respuesta de los gasterópodos y la ictiofauna ante la deforestación y la sedimentación en el manglar: Se fotointerpretaron las coberturas de manglar del delta con base en un sobrevuelo de 2009 y se clasificaron en cuatro tipos: 1: manglar de borde (dominado por R. mangle), 2: parte interna de manglar de cuenca (dominado por A. germinans), 3: manglar interno mixto con evidencias de entresaca (dominado por L. racemosa), y 4: potrero con algunosárboles de A. germinans remanentes (Castaño, 2012; Blanco y Castaño, 2012). Por otro lado, en la interfase manglar de cuenca con el potrero se estudió el “efecto borde” utilizando las siguientes categorías: interior del manglar, borde y potrero (Amortegui, 2011, Amortegui et al., 2013).
Dado que el gasterópodo Neritina virginea (Prosobranchia: Neritidae) es el organismo indicador más abundante en la zona de estudio y en todo el golfo de Urabá (Ortiz y Blanco 2012; Blanco y Castaño, 2012) se utilizó como grupo respuesta ante la deforestación y la sedimentación. A lo largo de cada categoría se muestreó N. virgínea en cuadros dispuestos al azar y se tomaron parámetros fisicoquímicos del agua intersticial. Las densidades fueron comparadas entre tipos de coberturas y categorías con respecto al borde usando un ANOVA de una vía. Se realizaron tres muestreos durante 2009 y 2010.
Para evaluar cómo la deforestación cambia la estructura de la comunidad del bosque, se registraron parámetros de estructura en los individuos de mangle, adoptando la metodología de cuadrantes centrados en un punto para describir el bosque y para obtener ocurrencia de especies, densidad,área basal y frecuencia (Cintrón y Schaeffer-Novelli, 1983; Dahdouh-Guebas y Koedam, 2008), se ubicaron transectos de manera perpendicular al gradiente topográfi co de inundación, midiendoárboles desde el borde con el mar o río, hasta la parte trasera o borde donde limitan con otra cobertura natural o de uso antrópico (Figura 7). Estas variables se relacionaron con los atributos del paisaje en el vecindario circundante, buscando explicar cómo la estructura,área, forma y porcentaje de borde antrópico en el parche, responden a la cercanía de estos a una vía y centro urbano (ver métodos en Estrada, 2013). Finalmente, para evaluar el efecto de la sedimentación sobre N. virgínea, se cuantificó su densidad en 20 cuadros dispuestos al azar dentro de 7 estaciones ubicadas en el interior del bosque, cerca de las trampas de sedimentación en el borde del manglar durante muestreos mensuales realizados entre septiembre 2009 y octubre 2010.

Todos los análisis se realizaron con los programas estadísticos: Minitab 16.0, Sigmaplot 11.0, Spss 19,0, y la georeferenciación con el programa ArcGis 10.1
Aspectos socio-culturales de la cuenca baja del río Turbo: para analizar algunas condiciones socio-ambientales de la zona y su relación con los actores sociales, los cambios en el paisaje y las fuerzas externas de cambio, durante el año 2009 se realizaron tres visitas con una duración de una semana cada una, y una cuarta con duración de cinco semanas (Martínez y Urán, 2012). Se utilizaron métodos clásicos de la investigación en ciencias sociales, tales como las entrevistas estructuradas y semi-estructuradas, las charlas informales y los recorridos por la zona en compañía de algunos habitantes (Rist y Dahdouh-Guebas, 2006; Aguirre-Baztan, 1997).
3. ANÁLISIS DE RESULTADOS
Influencias antrópicas sobre la hidrología y en la exportación de sedimentos en el golfo de Urabá: Aunque entre caudal y exportación de sedimentos existió una relación lineal directa (r2= 68,8%) y estadísticamente significativa (p<0,001), se observó que mientras la primera variable presentó un claro comportamiento estacional (independientemente de la estación limnimétrica), la segunda no (Figura 2); ello se debe, posiblemente, a las condiciones litológicas de la cuenca, en la cual hay eventos de lluvias extremas y pulsos de arrastre de sedimentos. Solamente la exportación de sedimentos del río León (estación Barranquillita), con la cuenca más grande de la vertiente oriental, muestra un comportamiento estacional consistente con el del caudal, lo cual es común en los ríos con extensas planicies aluviales y con poca intervención antropogénica. Los otros ríos que drenan cuencas más pequeñas, y que atraviesan el distrito bananero-platanero de Carepa, Apartadó y Turbo, tienen exportaciones de sedimentos que varían de manera errática a lo largo del año, pero también de año a año (datos no mostrados, Arroyave et al., 2012), sugiriendo la prevalencia de controles antrópicos, en especial la deforestación, sobre los naturales. Se ha establecido ampliamente que el aumento natural de la exportación de sedimentos hacia los ríos se debe al aumento de la escorrentía por acción de la alta precipitación o la topografía pendiente y la morfología y litología de la cuenca, pero es usualmente magnificada debido a la deforestación y cambio de cobertura y uso de los suelos (Farnsworth y Milliman, 2003; Milliman y Syvitski, 1992). Esto explica el por qué los picos de exportación de sedimentos en algunos ríos hacia el golfo de Urabá, no necesariamente ocurren en los meses de mayor precipitación y caudal (ver estación El Dos-Turbo en abril, Figura 2).

Por consiguiente, la conversión de bosques inundables y de tierras bajas a usos agrícolas, en este caso extensas plantaciones de plátano y banano, y a tierras de pastoreo, promueven la alta producción de sedimentos (Blanco, 2009), que son fácilmente transportados por la escorrentía producida por la alta precipitación. Esto explica el por qué los ríos del golfo de Urabá presentan uno de los niveles de producción de sedimentos más altos del Caribe (Blanco y Restrepo, en revisión). Por ejemplo, el río Turbo, con unárea de cuenca de 160 km2, presentó un pico de exportación de sedimentos de 2 kton/día, dato reportado para el mes de Abril en el que se da inicio a la temporada de lluvias. En las cuencas hidrográficas de Apartadó y Chigorodó a lo largo de los últimos 45 años la deforestación se dio con una tasa anual de 2% aproximadamente, provocando un incremento significativo de los sedimentos exportados por estos ríos (1,2 kton/día y 1,8 kton/día, respectivamente), sobre todo entre la década de los noventas, cuando hubo un gran desarrollo agrícola en la región (Arroyave et al., 2012).
Tasas de deforestación, exportación y depósito de sedimentos en la cuenca baja del río Turbo. En cuanto a estas dinámicas, se destaca la desaparición de los bosques naturales y los manglares entre 1960 y 1975, una disminución de los pastos desde 1960 hasta 2007 y un aumento acelerado de los cultivos en el mismo periodo (Figura 3), los cuales para el último año de estimación ocuparon el 76,1% delárea.
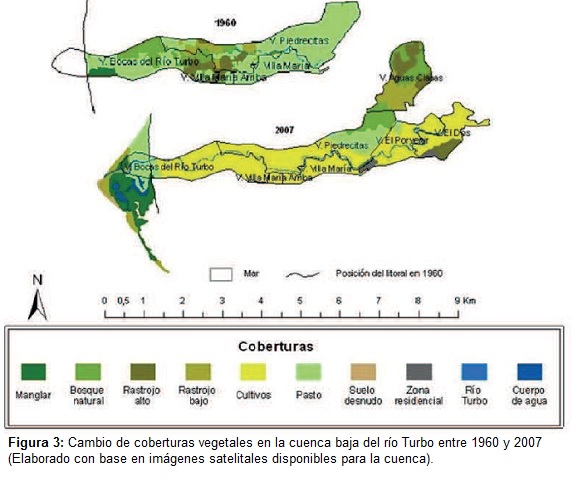
En cuanto a las coberturas naturales, se destacan los fragmentos de vegetación en estados intermedios de sucesión secundaria o recuperación (rastrojos). A partir de los mapas de coberturas, se cuantificó que las tasas de deforestación anual fueron de 2,64% durante el periodo 1960-1975, 5,65% durante 1975-1986 y, 1,36% durante 1960-2007. Por el contrario, en los periodos 1986-1994 y 1994-2007 hubo una ganancia de las coberturas naturales (0,54 y 0,018%, respectivamente) (Figura 4).

Dicha transición (bosque natural-pastos-cultivos) obedece a la ocupación y asentamiento del territorio de Urabá para usos múltiples (Keep, 2000; García, 1996). Desde elámbito biofísico, estas transiciones son típicas de regiones forestadas andinas y de tierras bajas caribeñas de Colombia y que han generado múltiples transformaciones en elárea afectada y diferentes grados de impacto sobre la flora y la biodiversidad en general (Etter y Wyngaarden, 2000).
Este grado de transformación de la cuenca baja del río Turbo puede explicar las altas tasas de exportación de sedimentos registradas por la estación El Dos del IDEAM, lo cual fue confirmado por nuestras mediciones. La EBS durante el 2010 mostró menores valores (1095 ± 2028 ton/mes) durante los meses de lasépocas seca y de transición, y mayores valores (31098 ± 51566 ton/mes) durante laépoca de lluvias (Figura 5B). Producto de esta exportación se observó un gradiente de las concentraciones de los SST, en dirección norte-sur ligado con la pluma turbia del río Turbo (Figura 5A). Asociado con esta pluma turbia, las mayores tasas de sedimentación se presentaron en las estaciones más próximas a la desembocadura y las menores en el extremo nororiental de la laguna costera y al sur, por lo que fue posible establecer dos zonas bien diferenciadas en términos de acumulación alta: desembocadura del río y sistema de canales y “ciénagas” aledañas, y espejo de agua de la bahía El Uno. Se planteó hipotéticamente que las crecientes inundan el manglar ubicado en la margen sur del río y aportan los sedimentos al sistema de canales y “ciénagas” (Ciénaga de Las Mujeres), ya que la comunicación con la bahía El Uno a través de su boca al sur es muy reducida y los niveles de sedimentos suspendidos y sedimentación de la misma son muy bajos. Aunque los vientos alisios del sur pueden generar corrientes y oleajes al interior de esta laguna costera, no parecen introducir porciones significativas de la pluma turbia del río Turbo a través de su reducida conexión con el golfo de Urabá (ver Figura 5). Finalmente, las cargas anuales de sedimentos correspondieron a 0,037 y 0,031 x106 ton/año para las estaciones de la desembocadura del río Turbo, en el mismo orden de magnitud de las reportadas por Restrepo (2005) para el mismo río (media 1966-1993: 0,0730x106 ton/ año), y otros del golfo (Guadualito = 0,0310; Apartadó = 0,0620; Grande = 0,0438 y Zungo = 0,0292 x106 ton/año). A lo largo del delta del río Turbo, el sitio de mayor sedimentación (2,54 ton/m2/año) tuvo valores entre 20 y 50 veces más altos que los reportados en otros manglares en el Trópico (Golbuu et al., 2003; Brunskill et al., 2004; Victor et al., 2006: rango: 4,5-127,8 kg/m2/año).

El delta del río Turbo y su laguna costera aledaña son una de lasáreas dentro del Golfo sometidas a mayores cambios antropogénicos. En primer lugar, la desembocadura original fue desviada hasta su posición actual a mediados del siglo veinte y, posteriormente, el curso natural en la parte baja ha sido desviado y rectificado repetidamente, por lo cual las crecientes pueden exportar más rápidamente los sedimentos a la zona costera. En segundo lugar, la minería, extracción de arenas, el lavado de suelos de zonas deforestadas y las captaciones de aguas, parecen haber alterado el balance hídrico y de sedimentos. Por estas razones, el delta del río Turbo es una de las pocasáreas de progradación y acreción a lo largo de la costa oriental del Golfo, en donde son habituales los procesos de erosión natural intensificados por la intervención humana, (Keep, 2000; Correa y Vernette, 2004; Restrepo, 2005).
Respuesta de los gasterópodos y la ictiofauna ante la deforestación y la sedimentación: Dado que los manglares son las barreras o filtros para el depósito de los sedimentos en sus suelos, primero se evaluó el efecto de la deforestación sobre la fauna. N. virgínea (densidad media±d.s.:16,2±26,3ind./ m2; N= 1027 cuadros observados) tuvo una densidad muy baja en los potreros en comparación con las franjas de R. mangle y A. germinans, durante el estudio basado en transectos (Figura 6D y F), Blanco y Castaño 2012). Adicionalmente, la ocurrencia de individuos expresada como el porcentaje de cuadrantes con presencia de individuos no mostró diferencias significativas entre las dos franjas de manglar, pero si una reducción significativa en lasáreas deforestadas dentro y fuera de ellas, sugiriendo que el número de grupos disminuye, aunque la especie presenta un patrón de distribución agregado (Figura 6B y C), modificado de Blanco et al., 2012). Con el estudio basado en parcelas, se demostró que la deforestación del manglar afecta negativamente a los gasterópodos del suelo dependiendo del grado de intervención por entresaca o potrerización por medio de un “efecto borde” (Amortegui et al., 2013). Por ejemplo, en este segundo estudio en el que se registraron 4671 individuos en total, se observó una disminución exponencial en la abundancia relativa desde el interior del manglar hacia el borde y el potrero a lo largo de unas pocas decenas de metros (87,24%, 11,8% y 0,90%, respectivamente), lo cual también fue consistente con la reducción de la ocurrencia de cuadros con individuos (Figura 6C y F). Los manglares no deforestados presentaron los mayores contenidos de materia orgánica, uniformidad en sedimentos y profundidad de charcas intermareales, mientras que los manglares afectados por entresaca (tala) o convertidos a potreros presentaron los mayores valores de pH y temperatura, mostrando que el conjunto de variables físico-químicas del agua intersticial y las propiedades de los sedimentos cambian significativamente a lo largo de la interfase manglar-potrero, siendo el aumento de la temperatura del aire y del suelo el efecto más notable de la “potrerización” del manglar, en la que generalmente se dejan algunosárboles de A. germinans en pie para proveer sombra al ganado (Blanco y Castaño 2012). Además de la pérdida de losárboles, la pérdida de plántulas y neumatóforos parece ser el mecanismo que incrementa el rigor ambiental y el riesgo de depredación (Blanco y Castaño, 2012, Amortegui, et al. 2013).

La pérdida de cobertura dada por la entresaca y potrerización del manglar son claros efectos de la deforestación en este ecosistema. La presión antrópica por reclamación de tierras con manglar para el establecimiento de pasturas están dando forma a estos bosques (Urrego et al. 2010), ya que al ubicarse en la parte trasera de los parches son susceptibles a ser intervenidos, principalmente en el costado oriental del golfo donde se ven sometidos a una reducción paulatina de la estrecha franja intermareal (Figura 7), adoptando formas complejas, sinuosas y alargadas que aumentan el borde expuesto del bosque. Desafortunadamente, las características deárea y forma responden a las condiciones antrópicas, al encontrar una fuerte relación en la longitud del borde en un parche y la proporción deéste influenciada por usos antrópicos, como potreros o cultivos (hasta un 73%; Estrada, 2013). Esta condición resulta más importante al encontrar una tendencia hacia la disminución del área, aumento de la densidad de parches por localidad, formas complejas y un desarrollo diamétrico y de altura menores en las especies de mangle, cuanto más cerca se encuentran los parches a una vía o el casco urbano del municipio de Turbo (Estrada, 2013). Además la entresaca de algunas clases diamétricas de R. mangle, que es la especie dominante en el Golfo, viene modificando la estructura de la comunidad: 1) al eliminar individuos de clases juveniles impidiendo que alcance un estado mayor, 2) al favorecer la introgresión del mangle invasor L. racemosa, la cual aumenta el valor de importancia cuando la extracción es más selectiva, y 3) al favorecer la dominancia del helecho de manglar Acrostichum aureum en el sotobosque cuando la tasa selectiva es severa, evitando la regeneración natural. Todo esto está llevando al ecosistema a una degradación ecológica criptica (Blanco et al., 2012). Las tasas de deforestación recientemente calculadas para la cabecera municipal de Turbo están cercanas al 2% anual para el periodo 1938-2009 y son una de las más altas del mundo (Estrada, 2013).
Lo anterior demuestra que los manglares del río Turbo y de la costa oriental del golfo de Urabá están experimentando una reducción delárea y un declive de las poblaciones de fauna béntica, similar a lo observado en otras partes del mundo. Los organismos macrobénticos son un componente importante para el manglar y, por lo tanto, cambios en sus poblaciones afectan directamente los procesos de reciclaje de nutrientes, la composición trófica y la aireación de suelos. Skilleter y Warren (2000) y Cannicci et al. (2008) mencionan que dichos organismos requieren condiciones específi cas al interior del bosque y son sensibles a los bordes creados por la deforestación continuada y otras alteraciones antropogénicas, cambiando su dinámica poblacional de manera inesperada, afectando negativamente el movimiento de las especies, la dispersión de juveniles, su densidad poblacional, distribución espacial y tamaño corporal.
En el delta del río Turbo no se observó un efecto negativo de la sedimentación sobre las poblaciones de N. virgínea, contrario a lo observado en los planos intermareales y submareales de los estuarios Neozelandeses, donde se ha estudiado extensamente el efecto de la sedimentación (sintetizado por Thrush et al. 2004). Entre las tres estaciones a lo largo del gradiente de sedimentación se encontró alta variabilidad en la densidad de N. virgínea (p<0,001). Contrario a lo esperado, la densidad promedio fue muy baja (3,87±8,93 ind./m2) en la estación con el nivel bajo de sedimentación y aumentó significativamente hacia los niveles intermedios (28,4±27,4 ind./m2) y altos (125,6±130,1 ind./m2). A pesar de este patrón, la varianza explicada fue relativamente baja (31.74%), lo cual sugiere que hay otros factores, posiblemente de microescala, igual o más importantes, o que estos niveles se encuentran por debajo de los niveles letales, lo cual apoya el análisis de regresión entre las tasas de sedimentación y las densidades promedios de cada estación y mes de muestreo que mostraron una relación positiva y significativa (r2=51,9%, p<0,001). Lo anterior se corroboró con un experimento in situ en el que se pudo determinar que los caracoles pueden aclimatarse luego de 15 a 17 días expuestos a diferentes gradientes de sedimentación (1, 3 y 18g) recuperando el peso corporal perdido durante los primeros días. También es posible que los sedimentos sean atrapados por las raíces de los mangles evitando que se depositen sobre los gasterópodos en las zonas abiertas. Sin embargo, no se descarta que la sedimentación tenga efectos negativos sobre otros grupos de la infauna como bivalvos y poliquetos, debido a que no se encontraron en los tamizajes de los sedimentos superficiales, los cuales se caracterizaron por ser arenas medias muy compactadas. Se recomienda estudiar en un futuro el efecto sobre los cangrejos, los cuales son otro grupo abundante y que tiene un papel de ingeniero ecosistémico.
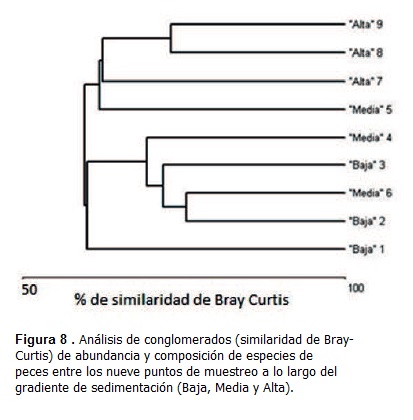
En cuanto a la ictiofauna, se recolectaron 803 especímenes pertenecientes a 17 familias y 28 especies, con una composición reflejo de lo esperado en estuarios tropicales (Vieira y Musick, 1994). La ordenación de las abundancias de estas especies en el análisis de similaridad de Bray-Curtis identificó patrones de asociación de especies y una posible zonación longitudinal de los peces (Figura 8), que parece parcialmente influenciada por los niveles de sedimentación, pero que también podría ser el resultado del efecto combinado de diferentes procesos fisicoquímicos y biológicos, entre ellos la disponibilidad de alimento yáreas de protección/reproducción, a lo largo de la bahía El Uno por lo cual sería mejor referirse a un gradiente ambiental y no únicamente de sedimentación (Sandoval, 2012). Se recomienda estudiar el cambio de comunidadesícticas desde la desembocadura del río Turbo hasta las aguas abiertas del golfo de Urabá para reducir los efectos de confusión observados dentro de la laguna costera (bahía El Uno).
La Tabla 1, muestra las diferentes dietas encontradas en las especies estudiadas. Estos resultados sugieren que la sedimentación puede tener un efecto en la dietas de manera directa por su posible influencia sobre la abundancia de algunas presas, particularmente las larvas de copépodos para Centropomus ensiferus. De manera indirecta, la sedimentación parece afectar el uso específico del espacio por parte de individuos de diferentes tallas de Notarius bonillai, ya que la dieta de los ejemplares encontrados en la zona de “alta” y “media” sedimentación (donde son más abundantes los individuos de tallas pequeñas) se caracterizó por la importancia de los copépodos mientras que en la zona “baja” (donde son abundantes los individuos de tallas grandes) fue compuesta principalmente de peces (centropomidos) y crustáceos (principalmente anomuros), lo cual refleja posibles cambios ontogénicos en la dieta del N. bonillai , y una posible segregación espacial en la zona de estudio.

En síntesis, se presentan indicios que sugieren que las altas concentraciones de sedimentos suspendidos o las altas tasas de sedimentación inciden negativamente sobre la abundancia de las especies de peces dominantes, posiblemente mediada por los efectos sobre la dieta de especies que se encuentran en la parte alta de la cadena alimenticia (carnívoros). No se descarta que los resultados también pueden estar directamente relacionados con la disponibilidad de alimento y a la oferta de refugio o protección del sistema (Laegdsgaard y Johnson 2001), y otros factores que influyen sobre la coexistencia en los estuarios, tales como la segregación del aprovechamiento de diferentes recursos alimenticios y del uso de diferentes hábitats (Sánchez-Gil, et al. 2008), independientemente del gradiente de sedimentación.
Aspectos socio-culturales de los habitantes de las zonas aledañas al manglar del delta del río Turbo: En estaárea se encuentran grupos poblacionales que cuentan con diferentes orígenes, intereses económicos y formas de relacionarse con el entorno, de acuerdo con sus historias particulares y con los servicios que el ecosistema les provee a cada grupo (Martínez y Urán, 2012). Estos grupos son: campesinos, habitantes recién-llegados al Uno, pescadores, carboneritos, los nuevos dueños y ganaderos, y cangrejeros. Las dinámicas actuales de poblamiento son tan altas que los actores sociales (locales) que se consideraban la mayor amenaza para el ecosistema han sido sobrepasados por la aparición de nuevos actores (foráneos) y por el cambio de la vocación de la tierra de agrícola y extracción de productos del bosque a pecuaria. La extracción tradicional de varas de mangle para construcción y para la producción de carbón vegetal, ciertamente impacta el manglar, pero este uso no ha derivado en su desaparición. Entre tanto, el fenómeno llamado localmente “potrerización” del manglar, el cual consiste en la tala de mangle y posterior siembra de pastos para ganadería, es la intervención antrópica que tiene los efectos de mayor impacto. La velocidad a la que se está deforestando el mangle bobo (L. racemosa ), ahora dominante en las franjas traseras del manglar, es aproximadamente de 5 días/ha. Teniendo en cuenta que la zona de manglar para el año 2007 era de 85,5ha (CORPURABA 2008), la desaparición del manglar es inminente. Tal vez logren permanecer algunos relictos de mangle colorado (R. mangle) en las orillas y de mangle de humo (A. germinans) dispersos en la parte trasera, pero esto no alcanzaría a mantener las funciones ecológicas del manglar. Quienes financian esta potrerización no ven utilidad económica en mantener las funciones ecológicas del manglar o revertir los cambios, ya que según su lógica estas tierras pueden ser más eficientes si se dedican de lleno a la ganadería, lo que para ellos constituye una forma de explotación más rentable. Eso también significa que el uso del mangle para esta nueva actividad supera con creces a la extracción de varas de mangle, tal y como lo pone de manifiesto el hecho de que durante más de cincuenta años se haya usado este ecosistema por parte de las comunidades allí asentadas, sin generar tasas de deforestación como las actuales. Estos actores también realizan modificaciones del curso del río (principalmente rectificaciones y en algunos casos excavaciones de nuevos cursos que unen meandros) en la planicie costera de manera eventual cuando se ven afectados por las inundaciones, lo cual no solo altera la velocidad de flujo de agua, sino que promueve el arrastre de sedimentos hacia el mar y los manglares. Tales situaciones acumuladas no solo afectan negativamente a los manglares y su fauna béntica y pelágica, sino que podrían reducir los servicios ambientales que presta a la comunidad, dado que cambia la percepción que los habitantes locales tienen de ellos, en detrimento del uso sostenible de los recursos naturales.
4. CONCLUSIONES
Es evidente que la deforestación en las cuencas hidrográficas costeras en el golfo de Urabá es una de las más altas y aceleradas del país, debido principalmente al intenso aprovechamiento de lasáreas con potencial agrícola y pecuario, lo que ha llevado a: 1) un detrimento de las coberturas vegetales boscosas en las planicies costeras, las cuales están relegadas a pequeños fragmentos de bosque natural, rastrojos yáreas de manglar pequeñas y estrechas en las zonas de acreción, tal como se observó en el delta del río Turbo, y 2) un aumento significativo en la magnitud de la producción y carga de sedimentos que son transportados por las diferentes corrientes de agua hacia el mar. En el delta del río Turbo, la exportación de sedimentos suspendidos y depósito de sedimentos siguió un patrón de distribución norte-sur a lo largo de la pluma turbia y la zona costera correspondiente a la progradación del delta, diferenciándose espacialmente dos importantesáreas: la desembocadura del río y la laguna costera. Esta exportación y depósito de sedimentos es modulada por la dinámica hidroclimatológica regional en la cual se resaltan eventos discretos de lluvias torrenciales y periodos prolongados de sequía. Finalmente, la conversión del manglar a potrero en el delta del río Turbo, produce un detrimento de los microhábitats físicos que sirven de refugio para los gasterópodos dominantes ante los depredadores, el rigor ambiental y la inundación de la marea, lo cual se observó claramente con N. virgínea que estuvo fuertemente restringida a las charcas remanentes en los potreros, lo que podría estar unido a la desecación de los suelos. Por lo anterior, la densidad y talla de estos gasterópodos pueden ser bioindicadores complementarios para las variables de grano grueso tales como la ausencia deárboles, la apertura del dosel y las huellas de ganado. La sedimentación parece no ser un tensor significativo para esta especie en elárea de estudio debido a que los efectos no parecen ser directos. Por otra parte, existe una relación débil entre los niveles de sedimentación y los atributos del ensambleíctico del delta del río Turbo, sin embargo, hay indicios para sugerir que puede existir una mayor influencia de la sedimentación sobre diversidad, abundancia y dieta de los peces depredadores tope (principalmente C. enciferus y N. bonillai) en la parte de “alta” sedimentación. Sin embargo, es necesario explorar si la baja riqueza observada en el bentos y el pelagos es producto de la alta sedimentación, por lo cual los efectos observados ya serían crónicos.
Este estudio demuestra que las poblaciones y comunidades bénticas y pelágicas asociadas a los manglares pueden verse afectadas por procesos de deforestación ex situ que afectan la producción de sedimentos que finalmente se depositan en la zona costera. Esta situación es magnificada por la tala selectiva y la conversión del manglar a potreros, actividades llevadas a cabo por diferentes actores y con dinámicas y motivaciones económicas diferentes. El presente trabajo demuestra la utilidad de la aproximación interdisciplinaria y de múltiples escalas espaciales y temporales para comprender problemas complejos como la sedimentación y la deforestación en las cuencas hidrográficas costeras.
AGRADECIMIENTOS
Los autores agradecen a la Universidad de Antioquia, a la Corporación Académica Ambiental de la Universidad de Antioquia, y a la Corporación para el desarrollo sostenible del Urabá, CORPOURABA, con los cuales se realizó el convenio 038/2009 que permitió la elaboración del proyecto “Impactos de la sedimentación sobre la estructura trófica, macrobentónica eíctica y el procesamiento de la hojarasca del manglar en el delta del río Turbo, golfo de Urabá, Caribe Colombiano”, de donde se deriva este documento. Resultados adicionales se obtuvieron del proyecto “Respuesta de los manglares y el macrobentos asociado a los cambios naturales y antrópicos en dos escalas de tiempo en el delta del río Turbo y bahía El Uno (golfo de Urabá)” (código UNAL-DIME 20101007166) a cargo de la investigadora Ligia Estela Urrego-Giraldo. Las aero-fotografías se obtuvieron con el proyecto “Expedición Estuarina, golfo de Urabá, Fase 1” parte de la “Expedición Antioquia 2013” financiada por la Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional y EAFIT. La fotointerpretación y verificación detallada de la costa oriental se realizaron se realizó con el proyecto “Fragmentación y deforestación del manglar en la costa oriental del golfo de Urabá” financiado por el CODI de la Universidad de Antioquia. Contribución ELICE No. 8.
REFERENCIAS BIBLIOGRAFICAS
Aguirre-Baztan A. 1997. Etnografía: metodología cualitativa en la investigación sociocultural. Editorial Alfaomega. Mexico. 356 P.
Álvarez-León R., Polanía, J. 1996. Los manglares del Caribe colombiano: síntesis de su conocimiento. Revista Académica Colombiana de Ciencias Exactas Físicas y Naturales: 20 (78): pp. 447-464
Amortegui V. 2011. Intervención antrópica (potrerización) en rodales de mangle negro Avicennia germinans (Magnoliopsida: Avicenniaceae) en relación a la distribución y abundancia de Neritina virginea (Gastropoda: Neritidae) en el golfo de Urabá, Caribe colombiano. [Tesis de pregrado]. [Medellín (Colombia)]: Instituto de Biología, Universidad de Antioquia. 71 P.
Amortegui V., Taborda A., Blanco J. F. 2013. Edge effect on a Neritina virginea (Neritomorpha, Neritinidae) population in a black mangrove stand (Magnoliopsida, Avicenniaceae: Avicennia germinans) in the southern Caribbean. Pan-American Journal of Aquatic Sciences. En prensa.
Arroyave A. 2011. Exportación de sedimentos desde cuencas hidrográficas de la vertiente oriental del golfo de Urabá (Caribe Colombiano) y su relación con factores climáticos y antrópicos: un análisis de series de tiempo y fotografías aéreas. [Tesis de pregrado]. [Medellín (Colombia)]: Instituto de Biología, Universidad de Antioquia. 93 P.
Arroyave A., Blanco J.F., Taborda A. 2012. Exportación de sedimentos desde cuencas hidrográfi cas de la vertiente oriental del golfo de Urabá: infl uencias climáticas y antrópicas. Revista Ingenierías Universidad de Medellín: 10(20): pp. 13-30.
Bernal G., Montoya L.J., Garizábal C., Toro M. 2005. La complejidad de la dimensión física en la problemática costeras del golfo de Urabá, Colombia. Gestión y Ambiente: 8 (1): pp. 123–135.
Blanco J.F. 2009. Banana crop expansion and increased river-borne sediment exports to the Gulf of Urabá, Caribbean Coast of Colombia. AMBIO, A Journal of the Human Environment. Royal Swedish Academy of Sciences: 38: pp. 181-183.
Blanco J.F., Londoño-Mesa M., Quan-Young L., Urrego-Giraldo L., Polanía J., Osorio A., Bernal G., Correa I. 2011. The Urabá Gulf Mangrove Expedition of Colombia. ISME/GLOMIS: 9 (3): pp. 8-10.
Blanco J.F., Estrada A., Ortiz L.F., Urrego L.E. 2012. Ecosystem-Wide Impacts of Deforestation in Mangroves: The Urabá Gulf (Colombian Caribbean) Case Study. ISRN Ecology: Article ID 958709. doi:10.5402/2012/958709
Blanco J.F., Castaño M.C. 2012. Efecto de la conversión del manglar a potrero sobre la densidad y tallas de dos gasterópodos en el delta del río Turbo (golfo de Urabá, Caribe colombiano). Revista de Biología Tropical: 60 (4): pp. 1707-1719.
Brusca R.C., Brusca G.J. 2003. Invertebrates. 2nd edition. Sinauer Associated, Sunderland, Massachusetts. 936 P.
Brunskill G., Zagorskis I., Pfi tzner J., Ellison J. 2004. Sediment and trace element depositional history from the Ajkwa River estuarine mangroves of Irian Jaya (West Papua), Indonesia. Continental Shelf Research 24: pp. 2535–2551.
Cannicci S., Burrows D., Fratini S., Smith III T., Offenberg J., Dahdouh-Guebas F. 2008. Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquatic Botany: 89: pp. 186–200.
Castaño M.C. 2012. Respuesta poblacional de dos gasterópodos bentónicos Neritina virginea (Gastropoda: Neritidae) y Melampus coffeus (Gastropoda: Melampidae) a los cambios en la cobertura del manglar (deforestación) en el delta del rio Turbo, golfo de Urabá [Tesis de pregrado]. [Medellín (Colombia)]: Instituto de Biología, Universidad de Antioquia. 48 P.
Cintrón G., Schaeffer-Novelli Y. 1983. Introducción a la Ecología del Manglar. Ofi cina Regional de Ciencia y Tecnología de la UNESCO para América Latina y el Caribe, ROSTLAC. Montevideo, Uruguay. 109 P.
Collins S.L., Carpenter S.R., Swinton S.M., Orenstein D.E., Childers D.L., Gragson T.L., Grimm N.B., Grove J.M., Harlan S.L., Kave J.P., Knapp A.K., Kofi nas G.P., Magnuson J.J., McDowell W.H., Melack J.M., Ogden L.A., Robertson G.P., Smith M.D., Whitmer A.C. 2011. An integrated conceptual framework for long-term social-ecological research. Frontiers in Ecology and the Environment: 9 (6): pp. 351-357.
CORPOURABA 2007. Actos resolutivos 001669 del 25 de septiembre y 001708 del 2 de octubre de 2007. Declaración para la ordenación la cuenca hidrográfi ca del río Turbo.
CORPOURABA 2003. Caracterización y zonificación de los manglares del golfo de Urabá. Departamento de Antioquia. Convenio 201671. FONAM–CORPOURABA, Alcaldía de Turbo, Universidad de Antioquia y Capitanía de Puerto de Turbo. Apartadó, Antioquia, Colombia. 80 P.
Correa I.D., Vernette G. 2004. Introducción al problema de la erosión litoral en Urabá (sector Arboletes- Turbo) costa Caribe colombiana. Boletín de Investigaciones Marinas y Costeras. Santa Marta, Colombia, 33: pp. 5-26.
Dahdouh-Guebas F., Koedam N. 2008. Long-term retrospection on mangrove development using transdisciplinary approaches: a review. Aquatic Botany, 89 (2): pp. 80-92.
Escobar E. 1999. El fi nal del salvaje. Naturaleza, cultura y política en la antropología contemporánea. CEREC- ICAN, Santa Fe de Bogotá. 401 P.
Estrada E.A., Gil S.M. 2005. Análisis de las condiciones morfológicas actuales como indicadoras de la evolución de la fl echa litoral, en la actual desembocadura del río Turbo [Tesis de pregrado]. [Turbo (Colombia)]: Ecología de Zonas Costeras, Universidad de Antioquia. 92 P.
Estrada E.A. 2013. Pérdida y fragmentación de los manglares, asociado a factores antrópicos y naturales en el golfo de Urabá. [Tesis de Maestría]. [Medellín (Colombia)]: M. Sc. Biología, Universidad de Antioquia. Etter A., McAlpine C., Stuart P., Pullar D., Possingham W. 2006a. Characterizing a tropical deforestation wave: a dynamic spatial analysis of a deforestation hotspot in the Colombian Amazon. Global Change Biology: 12: pp. 1409-1420.
Etter A., McAlpine C., Wilson K., Phinn S., Possingham H. 2006b. Regional patterns of agricultural land use and deforestation in Colombia. Agriculture, Ecosystems and Environment 114: pp. 369-386.
Etter A., Wyngaarden W. 2000. Patterns of Landscape Transformation in Colombia, with Emphasis in the Andean Region. Ambio 29 (7): pp. 432-439.
FAO, Organización de las Naciones Unidas para la Alimentación y la Agricultura. 2011. Situación de los Bosques del Mundo. Series bienales. Roma, Italia. Farnsworth K., Milliman J. 2003. Effects of climatic and anthropogenic change on small mountainous rivers: the Salinas River example. Global and Planetary Change: 39 (1-2): pp. 53-64.
Fischer W., Krupp F., Schneider W., Sommer C., Carpenter K.E., Niem V.H. (Red.). 1995. Pacifi co Centro-Oriental; Guía FAO para la identifi cación de especies para los fi nes de pesca. FAO; Roma, (Vol- II-III). 1652 P. García C.I. 1996. Urabá. Región, Actores y Confl icto. 1960-1990. Gente Nueva Editorial. 288 P. García-Valencia C. 2007. Atlas del golfo de Urabá: una mirada al Caribe de Antioquia y Chocó. Instituto de Investigaciones Marinas y Costeras –INVEMAR– y Gobernación de Antioquia. Serie de Publicaciones Especiales de INVEMAR Nº 12. Santa Marta, Colombia. 180 P.
Golbuu Y., Victor S., Wolanski E., Richmond R. 2003. Trapping of fi ne sediment in a semi-enclosed bay, Palau, Micronesia. Estuarine, Coastal and Shelf Science: 57: pp. 941–949.
Hogarth P.J. 2007. The biology of Mangroves and seagrasses. Second Edition.Published in the United States by Oxford University Press Inc., New York. 272 P.
Horn H.S. 1966. Measurement of overlap in comparative ecological studies. American Naturalist, 100: pp. 419-424.
Keep C.F. 2000. Monografi a de Turbo. Alcaldía del Municipio de Turbo, Antioquia. 543 P. Krebs C.J. 1989. Ecological methodology.Harper and Row, New York, 550 P.
Labropoulou M., Eleftheriou A. 1997. The foraging ecology of two pairs of congeneric demersal fi sh species: importance of morphological characteristics in prey selection. Journal of Fish Biology: 50: pp. 324-340
Laegdsgaard P., Johnson C. 2001. Why do juvenile fi sh utilize mangrove habitats? Journal of Experimental Marine Biology: 257(2): pp. 229-253.
Larsen M.C., Webb M.T. 2009. Potential Effects of Runoff, Fluvial Sediment, and Nutrient Discharges on the Coral Reefs of Puerto Rico.Journal of Coastal Research: 25: pp. 189-208
Martínez L., Urán A. 2012. Aspectos socio-culturales en elárea de manglar del delta del rio Turbo, golfo de Urabá: nuevas categorías de análisis socio ambiental. InformeTécnico. 29 P.
Milliman J.D., Syvitski J.P.M. 1992. Geomorphic/tectonic control of sediment discharge to the ocean: the importance of small mountainous rivers. Journal of Geology: 100: pp. 525-544.
Nagelkerken I., Blaber S.J.M., Bouillon S., Green P., Haywood M., Kirton L.G., Meynecke J.O., Pawlik J., Penrose H.M., Sasekumar A., Somerfi eld P.J. 2008. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquatic Botany: 89: pp. 155-185.
Ortiz L.F., Blanco J.F. 2012. Distribución de los gasterópodos del manglar Neritina virginea (Neritidae) y Littoraria angulifera (Littorinidae) en la Ecorregión Darién, Caribe Colombiano. Revista de Biología Tropical: 60 (1): pp. 219-232.
Patrick N.J., Sheridan G.J., Noske P.J. 2006. Changes in sediment loads and discharge from small mountain catchments following wildfi re in south eastern Australia. Journal of Hydrology: 331: pp. 495–510.
Rangel-Buitrago N. 2009. Contribución antropogénica a los cambios geomorfológicos y evolución reciente de la costa Caribe colombiana. Gestión y Ambiente: 12 (2): 43-56.
Restrepo J.D. 2005. Los Sedimentos del río Magdalena: Refl ejo de la Crisis Ambiental. Fondo Editorial Universidad EAFIT-Colciencias, Medellín, Colombia. 267 P.
Rist S., Dahdouh-Guebas F. 2006. Ethnosciences: A step towards the integration of scientifi c and indigenous forms of knowledge in the management of natural resources for the future. Environment, Development and Sustainability: 8 (4): pp. 467-493.
Ruppert E.E., Barnes R.D. 1996. Zoología de los invertebrados. Mc Graw -Hill, México. 114 P
Sánchez-Gil P., Yáñez-Arancibia A., Tapia-García M., Day J.W., Wilson C.A., Cowan J.H. 2008. Ecological and biological strategies of Etropus crossotus and Citharichthys spilopterus (Pleuronectiformes: Paralichthyidae) related to the estuarine plume, Southern Gulf of Mexico. Journal of Sea Research: 59: pp. 173-185.
Sandoval L.A. 2012. Efecto de la sedimentación sobre la ictiofauna en el delta del rio Turbo (golfo de Urabá, Caribe colombiano). [Tesis de Maestría]. [Medellín (Colombia)]: M. Sc. Ciencias Ambientales, Universidad de Antioquia.
Skilleter G.A., Warren S. 2000.Effects of habitat modifi cation in mangroves on the structure of mollusc and crab assemblages. Journal of Experimental Marine Biology and Ecology: 244: pp. 107–129.
Smith P.E., Zaret M.T. 1982. Bias in estimating niche overlap. Ecology: 63 (5): pp. 1248-1253.
Taborda A. 2008. Cambios de la cobertura del suelo en la cuenca baja del rio Turbo entre 1960 y 2007, departamento de Antioquia, golfo de Urabá, Caribe colombiano. Corporación para el Desarrollo Sostenible de Urabá, CORPOURABA. Informe Técnico. 64 P.
Thrush S.F., Hewitt J.E., Cummings V.J., Ellis J.I., Hatton C., Lohrer A.M., Norkko A. 2004. Muddy waters: elevating sediment input to coastal and estuarine habitats. Frontiers in Ecology and the Environment: 2: pp. 299-306.
Turner B.L., Robbins P. 2008. Land-change science and political ecology: similarities, differences, and implications for sustainability science. Annual Review of Environmental Resources: 33: pp. 295-316.
Urrego L., Molina E., Suárez Y., Polania J. 2010. Distribución, Composición y Estructura de los manglares del golfo de Urabá. Capitulo 3. pp 82-110 En: Blanco JF, Londoño M, Urrego L, Polonia J, Osorio A, Bernal G, Correa ID. 2010. Expedición Estuarina, Golfo de Urabá, Fase 1; Expedición Antioquia 2013, Gobernación de Antioquia, Universidad de Antioquia, Universidad Nacional de Colombia, Universidad EAFIT. (Informe Final). Medellín.
Valiela I., Bowen J.L., York J.K. 2001. Mangrove forest: one of the world´s threatened major tropical environments. BioScience: 51: pp. 807-815.
Victor S., Neth L., Golbuu Y., Wolanski E., Richmond R. 2006. Sedimentation in mangroves and coral reefs in a wet tropical island, Pohnpei, Micronesia. Estuarine, Coastal and Shelf Science: 66: pp. 409-416.
Vieira J.P, Musick J.A. 1994. Fish faunal composition in warm-temperate and tropical estuaries of western Atlantic. Atlantica: 16: pp. 31-53.
Wall G.R., Nystrom E.A., Litten S. 2008. Suspended Sediment Transport in the Freshwater Reach of the Hudson River Estuary in Eastern New York. Estuaries and Coasts: 31: pp. 542–553
Wang C.H.A., Chin L.S., Gupta A. 2005. Sediment Released to Coastal Waters of Southeast Asia: Seasonal and Spatial Distributions. Conference on Deltas HO Chi Minh City
Warne A.G., Webb R.M.T., Larsen M.C. 2005. Water, Sediment, and Nutrient Discharge Characteristics of Rivers in Puerto Rico, and their Potential Infl uence on Coral Reefs. U.S. Geological Survey Scientific Investigations Report. 206: 58 P.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2013 Gestión y Ambiente

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-CompartirIgual 4.0.
Los artículos que sean publicados en la revista Gestión y Ambiente, también serán publicados en el sitio web http://www.revistas.unal.edu.co/index.php/gestion/index y en formatos electrónicos como PDF, HTML, XML, entre otros. Además, en diferentes redes sociales de difusión del conocimiento. Gestión y Ambiente adopta directrices de ética por Committee on Publication Ethics (COPE) sobre buenas prácticas de conducta (evitar conductas como plagio, falsificación, autoría ficticia, entre otros), describe conflictos de interés o en competencia, contribuciones de autoría y fuentes de financiación. Todo lo publicado se considerará propiedad de la revista Gestión y Ambiente, pero pueden usarse bajo la licencia Creative Commons “Reconocimiento-No Comercial-Compartir Igual International (BY-NC-SA) 4.0”













