



Published
Dinámica de la Biomasa Aérea en un Bosque Pluvial Tropical del Chocó Biogeográfico.
Keywords:
Cambio climático global, C, Tasa de incremento neto de la BA (TINBA), Productividad primaria neta (PPN), Parcelas permanentes. (es)Downloads
DINÁMICA DE LA BIOMASA AÉREA EN UN BOSQUE PLUVIAL TROPICAL DEL CHOCÓ BIOGEOGRÁFICO
DYNAMICS OF TREE ABOVEGROUND BIOMASS IN A TROPICAL RAIN FOREST OF THE CHOCÓ BIOGEOGRÁFICO
Harley Quinto Mosquera1 y Flavio Moreno Hurtado2
1 Docente Ocasional. Universidad Tecnológica del Chocó "Diego Luis Córdoba". Facultad de Ciencias Básicas. Barrio Nocolás Medrano, Ciudadela Universitaria. A.A. 292, Quibdó, Colombia. <hquintom@unal.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <fhmoreno@unal.edu.co>
Recibido: Febrero 02 de 2010; aceptado: Abril 18 de 2011
Resumen. El estudio de la biomasa aérea (BA) de los bosques tropicales es fundamental para entender el balance del C global en el contexto del cambio climático. La BA se cuantificó en un bosque maduro de Salero (Chocó Biogeográfico), mediante ecuaciones diseñadas para bosques húmedos tropicales, a partir de datos de densidad de madera, diámetro (DAP) y altura de árboles (con DAP = 10 cm) medidos en dos sub-parcelas permanentes ("E" y "U") de 1 ha, las cuales se monitorearon en los años 1998, 2005 y 2008. La BA fue 237,31 t·ha-1, 259,99 t·ha-1 y 217,97 t·ha-1 respectivamente en la sub-parcela "E". Mientras que en la "U" fue de 178,94 t·ha-1y 179,17 t·ha-1 en los años 2005 y 2008; las diferencias de BA a través del tiempo fueron no significativas. Los incrementos promedios anuales de BA de sobrevivientes fueron 4,42 y 3,18 t·ha-1 año-1 en las sub-parcelas "E" y "U" respectivamente. Además, en sub-parcela "E" en condiciones imperturbadas, se presentó una tasa de incremento neto de la BA (TINBA) de 2,61 t·ha-1 año-1, en concordancia con la hipótesis del incremento en la BA en los bosques húmedos tropicales. La productividad primaria neta aérea (PPNA) en Salero de carbono fue de 5,21 t· ha-1 año-1, por lo tanto los resultados no apoyaron la hipótesis de la disminución en la productividad de los bosques tropicales con el incremento en la precipitación.
Palabras clave: Cambio climático global, C, tasa de incremento neto de la BA (TINBA), productividad primaria neta (PPN), parcelas permanentes.
Abstract. The study of the aboveground biomass (AB) of tropical forests is fundamental to understand the balance of the global C in the context of the climatic change. We quantified the AB in a mature forest of Salero (Chocó Biogeográfico), by means of equations designed for tropical humid forests, starting from data of wooden density, diameter (D) and height of trees (with D = 10 cm) measured in two permanent sub-parcels (E and U) of 1 hectare (ha), which were measured in the years 1998, 2005 and 2008. Inthis years the AB was of 237.31 t·ha-1, 259.99 t·ha-1 and 217.97 t·ha-1 respectively in the E sub-parcel. While in the U it was of 178.94 t·ha-1 and 179.17 t·ha-1 in the years 2005 and 2008; the differences of the aboveground biomass through the time were not significant. The increments annual averages in survivors' BA were of 4.42 and 3.18 t·ha-1 year-1 in the E and U sub-parcels respectively. Also, in sub-parcel E under conditions imperturbadas, the rate of net increment was presented in the AB (TINBA) of 2.61 t·ha-1 year-1, in agreement with the hypothesis of the increment in the BA in the tropical humid forests. The PPNA of carbon in Salero was of 5.21 t·ha-1 year-1, therefore the results didn't support the hypothesis of the decrease in the productivity of the tropical forests with the increment in the precipitation.
Key words: Change climatic global, carbon, net increment of the aboveground biomass, net primary productivity, permanent plot.
Los bosques tropicales son importantes en el balance del carbono (C) global, debido a que representan cerca del 59% del C total acumulado en los ecosistemas boscosos del mundo (Dixon et al., 1994). Dado que estos ecosistemas capturan CO2 atmosférico mediante el proceso de fotosíntesis y lo acumulan en sus tejidos (Chazdon y Montgomery, 2002; Clark et al., 2001a), el estudio de la dinámica de la biomasa y el C es fundamental para comprender el papel que éstos desempeñan en el ciclo global de este elemento (Clark et al., 2001a; Sierra et al., 2007) y en la mitigación del cambio climático (Brown, 1997; IPPC, 2001). En consecuencia, existe gran interés por evaluar las existencias, los patrones de captura y el balance de C de los bosques tropicales del mundo con el objetivo de develar si estos ecosistemas están o no acumulando C atmosférico adicional (Brown, 1997; Phillips et al., 1998; Malhi et al., 1998; Clark et al., 2001a; IPPC, 2001; Clark, 2002; Chave et al., 2003; Baker et al., 2004a; Houghton, 2005). Recientemente, estos estudios han cobrado mayor importancia por la necesidad de estimar las cantidades de C que podrían ser emitidas a la atmósfera en caso de deforestación (Houghton, 2005), lo cual es un insumo fundamental para la estructuración de proyectos de reducción de emisiones por deforestación y degradación (REDD) (Angelsen et al., 2009).
Según la teoría ecológica clásica (Odum, 1969; Whitmore, 1975; Bormann y Likens, 1979), los bosques naturales primarios imperturbados o climáxicos no acumulan C atmosférico adicional, ni presentan cambios netos en su biomasa total, pues se mantienen en equilibrio dinámico durante largos períodos de tiempo. Sin embargo, varios estudios realizados en la Amazonia han documentado que tales bosques sí están capturando C atmosférico en forma significativa, como lo evidencian los resultados de Phillips et al. (1998); Malhi et al. (1998); Baker et al. (2004b) y Malhi et al. (2004) en 104 parcelas neotropicales. Este comportamiento se debe posiblemente al aumento de CO2 atmosférico, el cual probablemente ha estado funcionando como fertilizante natural en las últimas décadas (Phillips et al., 1998; Malhi et al., 1998, 2004; Baker et al., 2004b).
No obstante, estos trabajos han sido fuertemente criticados, debido a la ocurrencia de posibles errores metodológicos (Clark, 2002); adicionalmente, no se han registrado tendencias similares en los bosques tropicales de Asia y África (Houghton, 2005). Feeley et al. (2007) encontraron que en los bosques maduros de Malasia y Barro Colorado (Panamá) se presentaron disminuciones en las tasas de crecimiento de un alto porcentaje de las especies, debido especialmente a la disminución en la precipitación total y a incrementos en la temperatura mínima diaria. Estos resultados contradicen la hipótesis de incremento de la productividad como lo plantean los estudios de la Amazonia citados atrás. En consecuencia, el debate está lejos de concluir, y al contrario, crece la incertidumbre sobre la capacidad de captura de C adicional por los bosques tropicales de cara al cambio climático.
De otro lado, la relación entre captura de C y precipitación en bosques tropicales es confusa principalmente porque son pocos los estudios que han analizado dicha relación y sus resultados son a veces contradictorios (Brown y Lugo, 1982; Clark y Clark, 1994; Schuur, 2003; Malhi et al., 2004). Por ejemplo, Schuur (2003) afirmó que la productividad disminuye notablemente cuando la precipitación alcanza valores superiores a 5.000 mm anuales, debido a factores como la disminución de nutrientes en el suelo, incremento de la lixiviación, aumento de la nubosidad y reducción de la tasa de descomposición de la materia orgánica. Malhi et al. (2004) por su parte, reportaron que la productividad de madera gruesa aérea no tuvo una relación obvia con la precipitación. Recientemente un análisis mundial sobre el balance de CO2 (Luyssaert et al., 2007), muestra que la productividad incrementa con el aumento de la precipitación, hasta que la precipitación alcanza valores de 1.500 mm anuales; donde sobrepasa estos valores la relación no es clara. Más aún, la productividad de los bosques tropicales ha sido pobremente estudiada cuando los niveles de precipitación sobrepasan los 5.000 mm anuales.
En Colombia, desde hace varios años se ha venido estableciendo una red de parcelas permanentes con el objetivo de conocer la dinámica y los patrones de distribución de especies en los bosques del país (Vallejo et al., 2005). Esta red representa una oportunidad excelente para probar hipótesis y mejorar la comprensión sobre el ciclo del C en los ecosistemas colombianos. No obstante, sólo en unos pocos sitios existen datos de más de cinco años de medición, entre los que se destacan los estudios realizados en la cuenca media del río Porce entre los años 1999 y 2006 (Orrego y Del Valle, 2003; Orrego et al., 2003; Yepes, 2008).
Con el ánimo de contribuir al entendimiento de la productividad del bosque y su relación con factores como la precipitación, se planteó el objetivo de estimar la biomasa aérea (BA) y su dinámica en el tiempo durante un período de 10 años en un bosque pluvial tropical del departamento del Chocó, respondiendo los siguientes interrogantes: ¿Cómo se distribuye la BA en función de las especies, las familias botánicas y los grupos funcionales? ¿Cómo varía la BA en un bosque pluvial tropical primario a través del tiempo? ¿Cómo se compara la productividad primaria neta aérea de este bosque con la de otros bosques tropicales sometidos a condiciones de precipitación más moderadas?
Área de Estudio. El estudio se realizó en una parcela permanente de investigación ubicada en un bosque pluvial tropical maduro de tierras bajas localizado en el corregimiento de Salero, municipio de Unión Panamericana, departamento del Chocó, Colombia, en las coordenadas 5º18'950'' de Latitud Norte y a los 76º36'742'' de Longitud Oeste (Figura 1). Este bosque se encuentra en la zona de vida de bosque pluvial tropical (bp-T) (Holdridge, 1982); la precipitación promedia anual de la región es cercana a 10.000 mm, la temperatura promedia varía entre 26 ºC y 30 ºC, la humedad relativa es de 90% y la altura sobre el nivel del mar oscila entre 100-150 m (García et al., 2003).

Figura 1. Ubicación de la parcela permanente de investigación en biodiversidad en el corregimiento de Salero, Unión Panamericana, Chocó, Colombia.
Los bosques de Salero son primarios, con muy poca intervención antrópica y con una alta diversidad de especies de plantas vasculares (Palacios et al., 2003). En este bosque se instaló una parcela permanente de investigación (PPIB) localizada al nororiente de la comunidad de Salero, desde la cual se tarda hora y media por trocha para llegar al sitio de la parcela, denominado localmente Catangüero. Este último se encuentra en el margen derecho de la quebrada Candelaria, la cual recoge las aguas de los riachuelos que escurren de la PPIB, entre ellos el Catangüero. En estos bosques el terreno es disectado, con diversos grados de pendiente; allí es común encontrar árboles caídos debido a distintos procesos de su dinámica natural.
Vegetación. Las familias más abundantes y frecuentes que caracterizan este ecosistema son Sapotaceae, Lecythidaceae, Arecaceae, Euphorbiaceae, Mirysticaceae, Chrysobalanaceae, Moraceae, Clusiaceae, Caesalpiniaceae, Vochysiaceae, Apocynaceae, Fabaceae y Bombacaceae (Asprilla et al., 2003). Entre las especies predominantes están el juanchito (Mabea chocoensis), palma mil pesos (Jessenia bataua), lechero (Brosimum utile), vaina (Matisia bullata), guasco (Eschweilera pittieri), palma memé (Wettinia quinaria), carbonero (Licania sp.), guayabo (Croton jorgei) y caimito (Chrysophyllum sp.), entre otras. (Palacios et al., 2003).
MÉTODOS
Establecimiento y re-censos de la parcela permanente. Generalmente para el estudio de la dinámica del C en los bosques se han empleado parcelas permanentes de investigación (Phillips et al., 1998; Clark et al., 2001; Del Valle et al., 2003; Chave et al., 2003; Baker et al., 2004a), las cuales permiten conocer los cambios en las existencias de biomasa y C en el bosque cuando se realizan mediciones sucesivas a lo largo de varios años. En este estudio se empleó una parcela permanente de investigación establecida entre abril y agosto del año 1998 mediante la metodología de BIOTROP (Melo y Vargas, 2003), y actualmente hace parte de la red nacional de parcelas permanentes de Colombia (Vallejo et al., 2005).
La parcela consiste en un cuadrado de 500x500 m (25 ha), dividido en 25 sub-parcelas rectangulares de 500x20 m (1 ha), que se identifican con las letras del abecedario desde la "A" hasta la "Y". Cada una de estas sub-parcelas está dividida en 25 cuadrantes de 20x20 m (400 m2) y a su vez, cada uno de los cuadrantes está dividido en cuatro cuadrículas de 10x10 m (100 m2) (García et al., 2003). El inventario de los árboles con DAP=10 cm se realizó en las sub-parcelas "E" y "U" de la parcela (Figura 2). Entre junio y septiembre del año 2005, y entre julio y agosto del año 2008 se realizaron el segundo y tercer censo de la parcela. En cada uno de los censos se registraron los individuos muertos, la forma de mortalidad y los individuos reclutados (que alcanzaron el DAP=10 cm).

Figura 2. Esquema de la parcela permanente de investigación en biodiversidad de Salero (Chocó, Colombia).
Identificación botánica. Se identificaron todas las morfoespecies hasta el máximo nivel taxonómico posible (NN, especie, género, familia botánica) en el herbario de la Universidad Tecnológica del Chocó "D.L.C." "Herbario Chocó". Esta identificación se hizo utilizando las claves especializadas de Forero y Gentry (1989), Gentry (1993) y Mahecha (1997). Además se contó con la colaboración del Biólogo Álvaro Cogollo del Jardín Botánico "Joaquín Antonio Uribe" de Medellín en la identificación del material.
Clasificación en grupos ecológicos de especies. Las especies identificadas taxonómicamente en las sub-parcelas "E" y "U", fueron clasificados de acuerdo a sus requerimientos de luz en climáxicas y pioneras, acorde con lo propuesto por Whitmore (1998). Además se empleó información de varios listados e inventarios de especies que incluían las características ecológicas de éstas. Entre los listados empleados están los citados por Swaine y Hall (1983); Whitmore (1998); Turner (2001); Unalmed (2001) y Corantioquia (2006), y los inventarios realizados por Córdoba et al. (2005) en claros naturales en bosques del Chocó, en los que se mencionan principalmente las especies pioneras o demandantes de luz. Para los individuos que se encontraban sin identificar taxonómicamente en la parcela de Salero (94 árboles), se asumió que pertenecían a especies climáxicas, en consideración a que éstas se encuentran en mayor proporción en los bosques primarios poco intervenidos; además las especies pioneras suelen ser comunes y por tanto fácilmente identificables.
Medición de diámetros en las sub-parcelas. Se midió la circunferencia a la altura del pecho en cm (1,30 m sobre el nivel del suelo) con cinta métrica a todos los árboles con DAP=10 cm en cada cuadrícula; posteriormente, los valores de circunferencia se transformaron a DAP. El perímetro del tronco del árbol donde se midió el diámetro por primera vez se marcó con pintura asfáltica amarilla con el objeto de garantizar que posteriores mediciones se hagan en la misma zona de la primera medición. Tales mediciones se hicieron en áreas libres de nudos o ramas. Además, se registró la ubicación de cada individuo en la cuadrícula. Todos los árboles medidos fueron marcados con placas de aluminio. También se identificó el hábito de crecimiento en las categorías de árbol, bejuco o liana y palma, se anotaron las características vegetativas y observaciones particulares de cada individuo.
Modelación de la altura total y comercial. Para modelar las alturas de los árboles se midió la altura total a 471 individuos, de los cuales 72 fueron árboles climáxicos medidos en la parcela y 399 individuos pioneros registrados en una zona alterada cercana a la parcela. Para estas mediciones se empleó el clinómetro Suunto® a distancias fijas de 15 y 20 m de observación. Para obtener la altura total de cada árbol se tuvo en cuenta el ángulo a la cima de la copa y la base del árbol, la distancia de observación y la altura de la persona que realiza la medición (Mostacedo y Fredericksen, 2000).
Con la información de la altura total, el DAP y el grupo ecológico de los 471 árboles medidos directamente en campo, se generaron modelos lineales de altura total en función del DAP. Con base en dichos modelos se estimó la altura a los demás árboles, teniendo en cuenta su grupo funcional. Estos datos de altura total fueron necesarios para estimar la biomasa aérea en algunos modelos. Por otro lado, dado que uno de los modelos empleados para estimar la biomasa aérea de los árboles requiere de la altura comercial (definida como aquella que va desde el suelo hasta el inicio de las ramas grandes de la copa del árbol) para su estimación y, debido a que esta altura no fue medida directamente en campo, se generó un modelo para estimarla. Para elaborar el modelo se empleó una base de datos de la Amazonia que incluía valores de altura (comercial y total) de especies como Coussarea sp., Posoqueria sp. Sterculia sp., Pouteria sp., Euterpe precatoria, Mabea sp., entre otras (Cuartas, 1993), que también han sido registradas en Salero (Palacios et al., 2003).
Densidad específica de la madera. Los valores de esta variable se tomaron de una base de datos internacional creada principalmente para bosques de la Amazonia (Baker et al., 2004 a); en los casos en los que alguna especie o género encontrado en Salero no se encontrara reportado en esta base de datos, se empleó el promedio del género o de la familia de la especie; para los individuos indeterminados taxonómicamente se utilizó el promedio general de densidad de la parcela de Salero.
Estimación de la biomasa aérea del bosque. Dado que la elección de un modelo o ecuación para estimar la BA de árboles es crucial, porque puede ser una de las principales fuentes de error en la estimación (Clark et al., 2001; Sarmiento et al., 2005), se recomienda emplear ecuaciones obtenidas localmente (Araujo et al., 1999; Chambers et al., 2001; Del Valle et al., 2003). Sin embargo, cuando se carece de ecuaciones locales se pueden emplear ecuaciones ya existentes, ideadas en condiciones climáticas y edáficas similares a la de la zona de estudio, que pueden dar resultados muy confiables (Clark et al., 2001), especialmente cuando las mediciones realizadas en campo con anterioridad son similares a las estimaciones del modelo. En este estudio, se cuantificó la BA de los árboles mediante siete modelos (Tabla 1) construidos con información de bosques húmedos tropicales de diversas zonas del mundo (Rodríguez, 1989; Brown, 1997; Chamberset al., 2001; Baker et al., 2004a; Chave et al., 2005). Se tomó como unidad de muestreo a los cuadrantes de 20 x 20 m2 de las sub-parcelas.
Tabla 1. Ecuaciones empleadas para estimar la BA en Salero (Chocó, Colombia). BA es la biomasa aérea de los árboles en kg, DAP es el diámetro en cm, Ln es el logaritmo neperiano, H es la altura total en m, Hc es la altura comercial en m, pi es la densidad de la madera.

El modelo de Rodríguez (1989) fue creado localmente en bosques del Chocó biogeográfico; para la estimación de la BA se usa DAP, altura total y altura comercial. Los modelos de Brown (1997) y Chambers et al. (2001) por su parte, solo incluyen DAP. Por último, los modelos de Baker et al. (2004a) y Chave et al. (2005) estiman la BA en función del DAP, altura total y densidad de la madera. Se examinaron las estimaciones de BA total con cada una de las ecuaciones y en cada uno de los censos realizados en la parcela. Para los análisis de BA a través del tiempo y en grupos ecológicos, se seleccionó el modelo con mayor coeficiente de correlación promedio en relación con las estimaciones de los otros modelos, como lo han realizado otros investigadores (Chave et al., 2003).
Dinámica de la biomasa aérea. Con la información de los inventarios de los años 1998, 2005 y 2008, se estimó la BA total a través del tiempo, las pérdidas por mortalidad de árboles, los incrementos por crecimiento y reclutamiento de árboles, y la tasa de incremento neto de la BA (TINBA), mediante la ecuación propuesta por Beers (1962), la cual consiste en: TINBA = ?s + I - C - M. En donde ?s corresponde al cambio en BA de los árboles sobrevivientes, calculada con la BA final (segunda medición) menos la BA inicial (primera medición) de cada individuo sobreviviente; I es la BA de los árboles reclutados (individuos nuevos que alcanzaron = 10 cm de DAP) registrados en la segunda medición; C es la BA de los árboles cortados y M es la BA de los árboles muertos entre las dos mediciones. Todas las estimaciones de BA fueron realizadas en t ·ha-1.
Por otro lado, en el periodo comprendido entre 2005 y 2008 se presentaron talas selectivas de árboles en ambas sub-parcelas, debido a esto los análisis de la variación de la BA se realizaron excluyendo e incluyendo los valores de BA de los arboles deforestados en cada sub-parcela, tomando como referencia el valor en el año 2005. En total, se registraron 186 y 97 individuos deforestados en las sub-parcela E y U respectivamente.
Análisis de datos. Se calcularon las medidas de tendencia central (media y mediana) y de dispersión (desviación estándar, coeficiente de variación y error estándar) de los datos. Mediante el coeficiente de correlación de Pearson se evaluó la correlación entre las distintas estimaciones de BA generadas con los siete modelos. Así mismo, se evaluaron los supuestos de normalidad y homogeneidad de varianzas con los estadísticos de Bartlett's y Hartley's; cuando los datos fueron normales y se cumplieron los supuestos se realizó un análisis de varianza ANAVA (Hoshmand, 1998). Cuando no se cumplieron los supuestos del ANAVA se empleó el estadístico de Kruskal Wallis para evaluar la existencia de diferencias significativas entre las medianas de la BA en función de los períodos de medición (1998, 2005 y 2008), sub-parcelas, cuadrantes, y grupo ecológico de especies. Mediante la prueba de rangos múltiples de Bonferroni se evaluó entre cuáles grupos se presentaron diferencias significativas de BA. Dado que en el proceso de depuración de la base de datos de campo se encontró que cerca del 30% de la información del DAP de 1998 en la sub-parcela "U" presentaba inconsistencias con relación con las mediciones de 2005 y 2008, se optó por excluir dichos datos del análisis general de la BA.
Para probar la hipótesis de la disminución de la PPN de los bosques tropicales con el incremento en la precipitación (Schuur, 2003), se estimó la productividad primaria neta aérea (PPNA) de la siguiente forma: PPNA= ?s + PH. Donde ?s es el incremento en BA de árboles sobrevivientes y PH es la producción de hojarasca. La producción de hojarasca (7,2 t·ha-1·año-1) de Salero empleada, fue la publicada por Quinto et al. (2007). En la estimación de la PPNA no fueron incluidas las pérdidas por consumidores y de compuestos orgánicos volátiles que hacen parte de la PPN porque representan proporciones poco significativas de la PPN total y generalmente no son medidas (Clark et al. 2001a).
Con la información de la base de datos de Clark et al. (2001b) incluidos los datos de Salero y el Bajo Calima, se establecieron rangos de precipitación promedio anual (de 500-2.000, 2.000-4.000, 4.000-6.000, 6.000-8.000 y de 8.000-10.000 mm anuales) con los cuales se realizó una prueba de Kruskal-Wallis para evaluar las diferencias significativas entre las medianas de la PPNA de cada rango de precipitación, y mediante el estadístico de rangos múltiples de Bonferroni se evaluaron los posibles grupos al nivel de los rangos de precipitación con relación a la PPNA (Guttman, 1982). Todos los análisis de los datos se realizaron con el programa Statgraphics Centurion XVI (Statistical Graphics Corporation, 2002).
RESULTADOS
Modelación de las alturas de los árboles en el bosque pluvial tropical de Salero, Chocó. Los modelos lineales generados para estimar la altura total y comercial en función del DAP, de las especies arbóreas climáxicas y pioneras del bosque pluvial de Salero (Chocó) se detallan en la Tabla 2. Todos los modelos fueron significativos y variaron en el tamaño de muestra y en el coeficiente de determinación (Tabla 2).
Tabla 2. Modelos lineales de la altura total y comercial de las especies arbóreas pioneras y climáxicas del bosque de Salero (Chocó, Colombia). En los modelos Ht es la altura total en m. Hc es la altura comercial en m. DAP es el diámetro a la altura del pecho en cm. E.E. es el Error estándar de los coeficientes. r2 es el Coeficiente de determinación. N. árboles muestreados. E.A.P. Error absoluto promedio.

Estimaciones de biomasa aérea en el bosque pluvial tropical de Salero, Chocó. Las estimaciones de BA de árboles realizadas con los siete modelos empleados presentaron diferencias estadísticamente significativas entre ellos (F= 42,98; P<0,05). Mediante la prueba de rangos múltiples de Bonferroni se encontró que los valores de los modelos 4, 5, 6 y 7 fueron estadísticamente similares. Las mayores estimaciones de BA se presentaron con el modelo 3, con 293,25 t·ha-1 promedio; mientras que con el modelo 1 se presentaron las menores estimaciones de BA, con 157,27 t·ha-1 promedio (Tabla 3).
Tabla 3. Estimaciones de biomasa aérea en la parcela permanente de investigación de Salero (Chocó, Colombia), con los siete modelos de estimación. El modelo 1 fue publicado por Rodríguez (1989), el modelo 2 por Brown (1997), el modelo 3 por Chambers et al. (2001), el modelo 4 por Baker et al. (2004) y los modelos 5, 6 y 7 por Chave et al. (2005).

Las estimaciones de BA de los modelos fueron diferentes con relación a los diámetros de los árboles. Dichas diferencias fueron menores en los árboles pequeños y se hicieron cada vez mayores en los árboles más grandes, especialmente en los modelos que emplean la densidad de la madera y la altura total de los árboles en sus cálculos (Figura 3). Sin embargo, las variaciones en la BA entre individuos influyeron de forma significativa sobre las estimaciones de BA total, a pesar de que dichas variaciones se compensaron entre sí en algunas de las estimaciones de BA al nivel de bosque (Tabla 3).

Figura 3. Estimación de la biomasa aérea de árboles en función del DAP para los siete modelos empleados. Estimación realizada con los datos del año 2005 de la sub-parcela "E" del bosque pluvial tropical de Salero (Chocó, Colombia).
Se registraron correlaciones significativas entre la BA estimada con los distintos modelos. Los modelos 5 y 6 presentaron las mayores correlaciones (0,984%), debido a esta alta correlación se optó por elegir el modelo 5 para realizar las evaluaciones de BA de los árboles. Por otro lado, el modelo 1 mostró la menor correlación (0,965%) (Tabla 4).
Tabla 4. Matriz de correlación de Pearson de los siete modelos de biomasa aérea empleados. Las correlaciones se realizaron con la BA de los individuos registrados en el año 2005 en la sub-parcela "E" en Salero (Chocó, Colombia). La última fila muestra el promedio de correlación de cada modelo. 
Distribución de la BA en las sub-parcelas de Salero, Chocó. En el año 1998 en la sub-parcela "E" se registraron en total 709 individuos. La BA estimada a partir de los cuadrantes de 20x20 m fue en promedio (± desviación estándar) de 237,31 ±101,61 t·ha-1. Así mismo, la BA en la sub-parcela "E" fue de 259,99 ± 109,25 t·ha-1, y de 179,23 ± 89,33 t·ha-1 en la sub-parcela "U" en el año 2005. En dicha medición se determinaron diferencias significativas (T= 7,43; P= 0,006) entre las medianas al nivel de las sub-parcelas. =5,28; P=0,021) en la BA total entre sub-parcelas en la medición del año 2008, cuando fueron incluidos los valores de BA de los árboles deforestados (Tabla 5).
Tabla 5. Estimaciones de biomasa aérea en el bosque pluvial de Salero (Chocó, Colombia). Las pruebas de hipótesis de varianzas fueron realizadas con la prueba Kruskal-Wallis. D.E. Desviación estándar.

Las estimaciones de BA total de los años 1998, 2005 y 2008 de la sub-parcela E, no mostraron diferencias significativas en sus medianas. Así mismo, la BA total de los años 2005 y 2008 de la sub-parcela U, no presentó diferencias significativas. No obstante, las estimaciones de BA total expresaron una tendencia al incremento con el paso del tiempo cuando fueron incluidos los valores de los árboles deforestados; aunque, dichas evaluaciones no evidenciaron diferencias significativas en sus medianas (Tabla 5).
En las especies, se encontró que Pouteria sp1, Eschweilera pittieri, Eschweilera sp, Brosimum utile, Qualea sp1 y Tetragastris panamensis fueron las que registraron mayor BA en las sub-parcelas en todos los períodos de medición. En todas las evaluacciones la especie más abundante fue Pouteria sp1 (Tabla 6). La distribución de la BA a nivel de los géneros mostró que Pouteria, Eschweilera, Licania, Brosimum, Tetragastris y Qualea fueron los que mayor BA registraron (Tabla 7). Por su parte, las familias Sapotaceae, Lecythidaceae, Chrysobalanaceae, Burseraceae, Moraceae, Myristicaceae y Vochysiaceae fueron las de mayor BA (Tabla 8). Cabe mencionar que Euphorbiaceae y Arecaceae, dos familias abundantes en los bosques estudiados, con 58 y 59 individuos respectivamente, presentaron baja BA (6,7 y 5,7 t·ha-1 respectivamente). La distribución de la BA en los niveles de especie, género y familias botánicas fue similar en todas las mediciones (Tablas 6, 7 y 8).
Tabla 6. Estimaciones de la biomasa aérea al nivel de especies en la parcela permanente de investigación de Salero (Chocó, Colombia). n es el número de individuos registrados.

Tabla 7. Estimaciones de la biomasa aérea al nivel de géneros en la parcela permanente de investigación de Salero (Chocó, Colombia). n es el número de individuos registrados.

Tabla 8. Estimaciones de la biomasa aérea al nivel de familias botánicas en la parcela permanente de investigación de Salero (Chocó, Colombia). n es el número de individuos registrados.

Al nivel de los cuadrantes, en la sub-parcela "E" en los años 2005 y 2008 el intervalo de clase modal (cerca del 32% en el 2005 y del 45% en el 2008) indicó una BA de entre 170 t·ha-1 y 270 t·ha-1. Mientras que en la sub-parcela "U" el intervalo modal (el 36% de los cuadrantes) presentó una BA entre 65 t·ha-1 y 137,5 t·ha-1 en el año 2005, y de entre 125 t·ha-1 y 200 t·ha-1 (el 36% de los cuadrantes) en el año 2008 (Figura 4).

Figura 4. Histograma de frecuencia de la biomasa aérea al nivel de cuadrantes en las sub-parcelas "E" y "U" en los años 2005 y 2008 en el bosque pluvial tropical de Salero (Chocó, Colombia).
En los grupos ecológicos de especies, la mayor BA y cantidad de individuos se manifestó en las especies climáxicas en todos los períodos de medición. En el año 2005, la BA de las pioneras fue mayor en la sub-parcela "E" que en la sub-parcela U, a pesar de exhibir una cantidad similar de individuos (156 en la "E" y 152 en la "U") (Tabla 9).
Tabla 9. Distribución de la biomasa aérea en grupos ecológicos de especies en las sub-parcelas "E" y "U" en los años 1998, 2005 y 2008 en el bosque pluvial tropical de Salero (Chocó, Colombia).

Dinámica de la BA en el bosque pluvial tropical en Salero, Chocó. En el período comprendido entre los años 1998 y 2005 se presentó el mayor incremento en la BA de los árboles sobrevivientes (4,42 t·ha-1·año-1) en la sub-parcela E, en comparación con los incrementos del período entre los años 2005 y 2008 de 1,69 t·ha-1·año-1 y 3,18 t·ha-1·año-1 en las sub-parcelas E y U respectivamente. Por otro lado, la TINBA fue positiva en el período entre los años 1998 y 2005 (con 2,62 t·ha-1·año), y en el período entre los años 2005 y 2008 fue negativa en ambas sub-parcelas (con -14,86 t·ha-1·año y -5,64 t·ha-1·año) (Tabla 10). Estos valores negativos se debieron a la alta producción de necromasa a causa de la intervención antrópica (extracción selectiva de madera). Aparte de esto, el reclutamiento en términos de BA fue evidente solo en el período entre 1998 y 2005. En consecuencia, el bosque incrementó significativamente su BA neta solamente en el período entre 1998 y 2005 (Tabla 10).
Tabla 10. Dinámica de la BA en el bosque pluvial tropical de Salero (Chocó, Colombia).

PPNA y precipitación promedia anual en bosques tropicales. En el bosque pluvial tropical de Salero, se registró una PPNA promedio (incremento en BA de sobrevivientes + producción de hojarasca) de carbono 5,21 t·ha-1·año-1. Los valores de PPNA de los rangos de precipitación (de 500 - 2.000; 2.000 - 4.000; 4.000 - 6.000; 6.000 - 8.000 y 8.000 - 10.000 mm anuales) presentaron un promedio general de carbono de 5,25 t·ha-1·año-1; así mismo, en el rango de precipitación de entre 500-2.000 mm anuales se registró la mayor PPNA con un promedio de carbono de 5,88 t·ha-1·año-1, mientras que en el rango de precipitación de entre 4.000 -6.000 mm anuales se observó la menor PPNA con un promedio de carbono de 2,43 t·ha-1·año-1 (Figura 5).

Figura 5. Análisis de varianza de la productividad primaria neta aérea con relación a los rangos de precipitación promedia anual en bosques tropicales. Las X son los promedios de PPNA y las líneas horizontales son los intervalos de confianza de los datos. Los valores dentro de los circulos azules y rojos muestran los grupos formados mediante la prueba de Bonferroni. Datos tomados de Clark et al. (2001) e incluidos los valores de Salero (Chocó, Colombia) y Bajo Calima (Valle del Cauca, Colombia).
Mediante el estadístico de Kruskal-Wallis se detectaron diferencias significativas entre las medianas de la PPNA (estadístico = 12,99; P = 0,011) de los rangos de precipitación evaluados; y mediante el estadístico de rangos múltiples de Bonferroni se identificaron dos grupos de rangos de precipitación con relación a la PPNA, el primer grupo estuvo formado por los rangos de 500-2.000, 2.000-4.000 y 8.000-10.000 mm anuales, mientras que el segundo estuvo conformado por los rangos 4.000-6.000, 6.000-8.000 y 8.000-10.000 mm anuales (Figura 5). El hecho de que el rango de entre 8.000-10.000 mm anuales (correspondiente a Salero) hiciera parte de ambos grupos estadísticos, mostró la poca influencia de la precipitación sobre la PPNA.
DISCUSIÓN
Exactitud de la estimación de la biomasa aérea con los modelos utilizados. Los modelos 1 y 3 empleados para estimar la BA de árboles en Salero (Chocó), generaron valores estadísticamente diferentes a los demás modelos (Tabla 2), lo cual también ha sido reportado por diversos investigadores, quienes han documentado que distintas ecuaciones generan estimaciones diferentes de BA (Araujo et al., 1999; Clark et al., 2001; Keller et al., 2001; Sarmiento et al., 2005). Se ha sugerido que los modelos que solo emplean una variable como el DAP en la estimación de la BA, podrían dar origen a mayores errores, pues se ignoran las variaciones en arquitectura y densidad de la madera de las especies (Clark et al., 2001; Chave et al., 2003), especialmente en bosques con alta diversidad de especies como los del Chocó (Faber y Gentry, 1991). Sin embargo, en el presente estudio se observó que cuando se analizan los valores de BA a nivel de bosque (BA total), las estimaciones tienden a ser muy similares, debido probablemente a que los valores se compensan entre sí (Tabla 3), por lo cual el margen de error es técnicamente aceptable, a pesar de que el margen de error de los inventarios estén alrededor del 20%; así ha sido documentado también en bosques de la Amazonia brasilera por investigadores como Baker et al. (2004a). Por otro lado, las variaciones en la BA al nivel de los cuadrantes de las sub-parcelas E y U, están parcialmente explicadas por las variaciones en las características edáficas de la zona, que han sido analizadas con mayor detalle en otros estudios, como Quinto y Moreno (2010) y Quinto y Moreno (sin publicar).
Comparación de la BA de Salero (Chocó) con otras estimaciones en bosques tropicales. La BA en el bosque pluvial tropical de Salero tuvo un rango de entre 179,2 y 259,9 t·ha-1, con un promedio general de 216,8 t·ha-1. Dicha BA fue ligeramente superior a los informes de BA realizadas por Mosquera y Sánchez (2005), Faber y Gentry (1991) y Rodríguez (1989) de 176 t·ha-1, 169-193 t·ha-1 y 190 t·ha-1 respectivamente, realizadas con anterioridad en distintos bosques del Chocó Biogeográfico. En general, los valores de BA encontrados en Salero (Chocó) de 216,8 t·ha-1, se encuentran en la porción inferior del rango de BA de árboles (entre 167 y 419 t·ha-1) referido por Sarmiento et al. (2005) para selvas neotropicales de baja altitud. Estos resultados de BA de Salero (Chocó) también se encuentran dentro del rango (entre 45-649 t·ha-1, con 275,16 t·ha-1 promedio) expresado por Clark et al. (2001b) para bosques tropicales de distintas zonas del mundo.
Incremento de la BA del bosque pluvial tropical de Salero, Chocó. La BA del bosque pluvial no cambió significativamente en el tiempo (Tabla 5), debido a esto, los resultados en términos de la BA total, no apoyan la hipótesis del incremento en BA en bosques tropicales planteada por Phillips et al. (1998), Malhi et al. (1998, 2004) y Baker et al. (2004b). En el bosque pluvia tropical de Salero, Chocó, se encontró un incremento promedio anual de BA de árboles sobrevivientes de entre 1,69 y 4,42 t·ha-1·año-1 (Tabla 10), el cual está dentro del rango de incremento en BA de árboles reportado por Clark et al. (2001b) de entre 0,6 a 7,6 t·ha-1·año-1, para diversos bosques tropicales. Por otro lado, solo la TINBA de la sub-parcela "E" entre 1998 y 2005, presentó un promedio de 2,61 t·ha-1·año-1 (Tabla 10), que no incluyó el cero en sus intervalos de confianza (Tabla 11). Por tal razón, el bosque presentó un incremento positivo de BA, sólo en dicho período, superior a los incrementos reportados por Chave et al. (2008) para diferentes bosques tropicales (Tabla 11). Este hecho (TINBA de 2,61 t·ha-1·año-1), está acorde con lo planteado por la hipótesis del incremento en la BA en los bosques húmedos tropicales registrado en las ultimas décadas (Phillips et al., 1998; Malhi et al., 1998, 2004; Baker et al., 2004b).
Tabla 11. Valores de BA, incremento, reclutamiento, mortalidad y cambio neto de la biomasa aérea de bosques primarios tropicales incluidos los datos de Salero (Chocó, Colombia), modificado de Chave et al. 2008. BA es biomasa aérea (t·ha-1). Los datos son de la sub-parcela "E" (años 1998-2005).

Al analizar las variaciones a través del tiempo de la BA en términos de la TINBA fue posible detectar los cambios en las capturas y emisiones de carbono de los bosques, en comparación con los análisis en términos de la BA total, debido a que los cambios en la BA total a través del tiempo no apoyaron la hipótesis del incremento en la captura de carbono de los bosques tropicales (Phillips et al., 1998, Baker et al., 2004b). Así mismo, los valores negativos en las TINBA del periodo entre 2005 y 2008 (Tabla 11) fueron producto de las deforestaciones; frente a lo cual, solo se puede aseverar que en la parcela debido a la intervención se incrementó la emisión de carbono a la atmosfera, la fragmentación y la masa de detritos, como se ha documentado en otros bosques tropicales (Laurance et al., 1998; Gerwing, 2002; Broadbent et al., 2008).
PPNA y precipitación promedia anual en bosques tropicales. Recientemente, Schuur (2003) manifestó que en los bosques tropicales con precipitación promedia anual (PPA) superior a 5.000 mm, la PPNA tiende a disminuir debido a la reducción de la tasa de descomposición y de la disponibilidad de nutrientes; además, por el incremento en la lixiviación y la nubosidad que merman la PPN. Sin embargo, en este estudio, mediante estadístico de rangos múltiples no se encontraron diferencias significativas entre los promedios de PPNA de los bosques tropicales con precipitaciones entre 500-4.000 y de 8.000-10.000 mm anuales (Figura 5); con lo cual, los resultados obtenidos, contradicen la hipótesis de Schuur (2003), ya que no fue evidente una disminución en la PPNA con el incremento en la precipitación (Figura 5).
El hecho de que la PPNA registrada fuese alta, a pesar de alta precipitación (contrario a Schuur (2003), se debió posiblemente a las siguientes razones: 1) No hay limitación de oxígeno para la descomposición en el suelo por exceso de agua, debido a que en estos ecosistemas a pesar de las altas precipitaciones los suelos tienen buen drenaje. 2) las tasas de descomposición de hojarasca de las zonas en las que se presentan anegamientos temporales, posiblemente son similares a las de las áreas bien drenadas. Puesto que en los suelos en los que el nivel freático oscila unos pocos centímetros con las inundaciones, la hojarasca se encuentra sometida a períodos alternativos de anegamiento y aireación, dependiendo de las lluvias, esto produce pulsos de oxidación de la hojarasca haciendo que se descomponga, en las áreas inundadas, a tasas similares a las registradas en suelos sin inundación (Del Valle, 2003). 3) Los niveles de precipitación en el bosque pluvial tropical cercano a Salero (Chocó), varían significativamente a lo largo del día. A tal punto que se presentan lluvias con mayor frecuencia e intensidad en las horas de la noche (Murillo et al. 2005). Esto implica que, en las horas de mayor radiación solar, entre las 10:00 y las 15:00 horas, la fotosíntesis no es inhibida por la precipitación, y por ende la PPN puede registrar valores altos.
Para Baker et al. (2003), en bosques tropicales no estacionales, en los que la disponibilidad de agua no es una limitante, la radiación solar es la variable ambiental más influyente sobre las tasas de crecimiento. De manera similar, en el bosque pluvial tropical de Salero (Chocó), a pesar de presentar una alta PPA, las lluvias no constituyeron una limitante para la PPN, debido probablemente a que en la zona se registra una alta radiación solar, los suelos son bien drenados, las tasas de descomposición de la hojarasca son altas, como se han documentado en bosques del municipio de Lloró (Chocó) muy cercanos al área de estudio (Quinto y Ramos, 2009), y hay una buena disponibilidad de nutrientes para las plantas (Quinto y Moreno, 2010). Por otro lado, cuando se supera el margen de los 10.000 mm en PPA en bosques tropicales, el panorama para la PPN es aún incierto, pues no ha sido muestreado.
En síntesis, los valores de PPNA obtenidos en este estudio, contradicen la hipótesis de la disminución de la PPNA de los bosques tropicales cuando la precipitación es muy alta, propuesta por Schuur (2003). Además, se evidenció que el bosque en condiciones imperturbadas captó de forma significativa carbono atmosférico, acorde con la hipótesis del incremento en BA de los bosques tropicales de Phillips et al., 1998; Malhi et al., 1998, 2004; Baker et al., 2004a. Por último, queda probado el efecto negativo de las intervenciones antrópicas, cuando son realizadas sin planes de manejo sostenible del recurso, sobre las existencias y tasas de incremento neto de la BA, porque cuando se deforesta masivamente los bosques las emisiones de carbono superan las tasas de captura, de ahí la importancia de la conservación y el manejo sostenible de estos complejos ecosistemas, especialmente en regiones tropicales como el Chocó Biogeográfico.
AGRADECIMIENTOS
Este trabajo contó con el apoyo del proyecto "CRECIMIENTO Y PRODUCTIVIDAD DEL BOSQUE EN UN GRADIENTE ALTITUDINAL: Efectos Potenciales en el Funcionamiento del Sistema de Transmisión de Energía de Colombia", cofinanciado por la Empresa de Interconexión Eléctrica S. A. (ISA) y la Universidad Nacional de Colombia, Sede Medellín, mediante el programa de becas del Fondo de Investigaciones I.S.A. - Universidad Nacional.
BIBLIOGRAFÍA
Angelsen, A., S. Brown, C. Loisel, L. Peskett, Ch. Streck y D. Zarin. 2009. Reducing emissions from deforestation and forest degradation (REDD): an options assessment report. Prepared for the government of Norway.Meridian Institute, http://www.REDD-OAR.org. 100 p.; consulta: febrero 2009.
Araujo, T.M., N. Higuchiand and J.A. Carvalho. 1999. Comparison of formulae for biomass content determination in a tropical rain forest site in the state of Para, Brazil. Forest Ecology and Management 117(1-3): 43-52.
Asprilla, A., C. Mosquera, H. Valoyes, H. Cuesta y F. García. 2003. Composición florística de un bosque pluvial tropical (bp-T) en la parcela permanente de investigación en biodiversidad (PPIB) en Salero, Unión Panamericana, Chocó. pp. 39-44. En: García, F., Y. Ramos, J. Palacios, J.E. Arroyo, A. Mena y M. González (eds.). Salero diversidad biológica de un bosque pluvial tropical (bp-T). Editora Guadalupe Ltda., Bogotá. 207 p.
Baker, T.R., M.D. Swaine, y D.F. Burslem. 2003. Variation in tropical forest growth rates: combined effects of functional group composition and resource availability. Perspectives in Plant Ecology, Evolution and Systematics 6(1-2): 21-36.
Baker, T.R., O.L. Phillips, Y. Malhi, S. Almeida, L. Arroyo, A. Di Fiore, T. Erwin, T.J. Killeen, S.G. Laurance, W.F. Laurance, S.L. Lewis, J. Lloyd, A. Monteagudo, D.A. Neill, S. Patiño, N.A. Pitman, J.N. Silva and R. Vásquez. 2004a. Variation in wood density determines spatial patterns in Amazonian forest biomass. Global Change Biology 10: 545-562.
Baker, T. R., O.L. Phillips, Y. Malhi, S. Almeida, L. Arroyo, A. Di Fiore, T Erwin, N Higuchi, T. Killeen, S.G. Laurance, W.F. Laurance, S.L. Lewis, A. Monteagudo, D.A. Neill, P.N Vargas, N.C.A. Pitman, N. Silva y R. Vásquez. 2004b. Increasing biomass in Amazonian forest plots. Philosophical Transactions of the Royal Society. 359:353-365.
Beers, T.W. 1962. Components of forest growth. Journal of Forestry 60: 245-248.
Bormann, F.H. and G.E. Likens. 1979. Catastrophic disturbance and the steady state in northern hardwood forests. American Scientist 67(6): 660-669.
Broadbent, E.N., G. Asner, M. Keller, D. Knapp, P.J. Oliveira and J.N. Silva. 2008. Forest fragmentation and edge effects from deforestation and selective logging in the Brazilian Amazon. Biological Conservation 141(7): 1745-1757.
Brown, S. and A.E. Lugo. 1982. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 14: 161-187.
Brown, S. 1997. Estimating biomass and biomass change of tropical forests: A primer. Food and Agriculture Organization (FAO), Rome. 57 p. (Forestry Paper No. 134).
Chambers, J.Q., J. Dos Santos, R.J. Ribeiro and N. Higuchi. 2001. Tree damage, allometric relationships, and above-ground net primary production in central Amazon forest. Forest Ecology and Management 152: 73-84.
Chave, J., R. Condit, S. Lao, J.P. Caspersen, R.B. Foster and S.P. Hubbell. 2003. Spatial and temporal variation of biomass in a tropical forest: results from a large census in Panamá. Journal of Ecology 91(2): 240-252.
Chave, J., C. Andalo, S. Brown, M. Cairns, J. Chambers, D. Eamus, H. Folster, F. Fromard, N. Higuchi, T. Kira, J. Lescure, B. Nelson, H. Ogawa, H. Puig, B. Riera and T. Yamakura. 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145: 87-99.
Chave, J., R. Condit, H. Muller, S. Thomas, P. Ashton, S. Bunyavejchewin, L. Co, H. Dattaraja, S. Davies, S. Esufali, C. Ewango, K. Feeley, R. Foster, N. Gunatilleke, S. Gunatilleke, P. Hall, T. Hart, C. Hernández, S. Hubbell, A. Itoh, S. Kiratiprayoon, J. La Frankie, S. Loo de Lao, J. Makana, M. Noor, A. Kassim, C. Samper, R. Sukumar, H. Suresh, S. Tan, J. Thompson, M. Tongco, R. Valencia, M. Vallejo, G. Villa, T. Yamakura, J. Zimmerman and E. Losos. 2008. Assessing evidence for a pervasive alteration in tropical tree communities. Plos Biology 6(3): 1-19.
Chazdon, R. y R. Montgomery. 2002. La adquisición de carbono en las plantas. pp. 167-192. En: Guariguata M. y G. Kattan (eds). Ecología y conservación de bosques neotropicales. Libro Universitario Regional, Cartago, Costa Rica. 691 p.
Clark, D.A. 2002. Are tropical forests an important carbon sink? Reanalysis of the long-term plot data. Ecological Applications 12(1): 3-7.
Clark, D.A. y D.B. Clark. 1994. Climate-induced annual variation in canopy tree growth in a Costa Rican tropical rain forest. Journal of Ecology 82(4): 865-872.
Clark, D.B. and D.A. Clark. 2000. Landscape-scale variation in forest structure and biomass in a tropical rain forest. Forest Ecology and Management 137: 185-198.
Clark, D.A., S. Brown, D. Kicklighter, J. Chambers, J. Thomlinson and J. Ni. 2001a. Measuring net primary production in forest: concepts and field methods. Ecological Aplications 11(2): 356-370.
Clark, D.A., S. Brown, D. Kicklighter, J. Chambers, J. Thomlinson, E. Holland and J. Ni. 2001b. Net primary production in forest: an evaluation and sinthesis of existing field data. Ecological Aplications 11(2): 356- 370.
Corporación Autónoma Regional del Centro de Antioquia (CORANTIOQUIA). 2006. Diversidad y productividad de los bosques de Antioquia. Proyecto "Manejo y conservación de la flora". Informe final. Primera fase. CORANTIOQUIA - Universidad Nacional de Colombia, Medellín. 87 p.
Córdoba, J.A., D. González, Y. Ramos, D. Serna, y N. Panesso. 2005. Regeneración natural en claros de un bosque pluvial tropical en Pacurita, Chocó - Colombia. Revista Institucional. Universidad Tecnológica del Chocó D.L.C. 23: 11-19.
Cuartas, C. 1993. Estimadores cuantitativos de la biomasa aérea de especies dominantes en un bosque inundable de la Amazonia colombiana. Trabajo de grado de Ingeniería Forestal. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 124 p.
Del Valle, J.I. 2003. Descomposición de la hojarasca fina en bosques pantanosos del Pacifico colombiano. Interciencia 28(3):148- 153.
Del Valle, J.I., F. Moreno y S. Orrego. 2003. Los bosques tropicales y su contribución a la mitigación del cambio climático. pp. 3-43. En: Orrego S.A., J.I. Del Valle y F.H. Moreno (eds.). Medición de la captura de C en ecosistemas forestales tropicales de Colombia. Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia, Medellín. 314 p.
Dixon, R.K., S. Brown, R. Houghton, A. Solomon, M. Trexler and J. Wisniewski. 1994. Carbon pools and flux of global forest ecosystems. Science 263: 185-190.
Faber, D. and A.H. Gentry. 1991. The structure and diversity of rain forests at bajo Calima, Chocó region, western Colombia. Biotropica 23(1): 2-11.
Feeley, K.J., S. Wright, M. Supardi, A. Kassim and S. Davies. 2007. Decelerating growth in tropical forest trees. Ecology Letters 10: 461-469.
Forero, E. y A. Gentry. 1989. Lista anotada de plantas del departamento del Chocó, Colombia. Instituto de Ciencias Naturales - Universidad Nacional de Colombia. Museo de Historia Natural. Editorial Guadalupe Ltda., Bogotá. 142 p.
García, F., Y. Ramos, J. Palacios, J. Arroyo, E. Mena y M. González. 2003. Salero diversidad biológica de un bosque pluvial tropical (bp-T). Editorial Guadalupe Ltda., Bogotá, Colombia. 207 p.
Gentry, A. 1993. A field guide to the families and genera of woody plants of Northwest South America (Colombia, Ecuador, Perú). University of Chicago. Chicago. 895 p.
Gerwing, J. 2002. Degradation of forests through logging and fire in the eastern Brazilian Amazon. Forest Ecology and Management 157: 131-141.
Guttman, I. 1982. Linear models: an introduction. First edition. Wiley, John and Sons Incorporated, USA. 358 p.
Hogan, K.P. y J.L. Machado. 2002. La luz solar: consecuencias biológicas y medición. pp. 119-143. En: Guariguata, M. y G. Kattan (eds). Ecología y conservación de bosques neotropicales. Libro Universitario Regional. LUR. Costa Rica. 691 p.
Holdridge, L.P. 1982. Ecología basada en las zonas de vida. Instituto Interamericano para la Agricultura. San José, Costa Rica. 216 p.
Hoshmand, A.R. 1998. Statistical methods for environmental and agricultural sciences. Second edition. CRC Press LLC, New York, USA. 439 p.
Houghton, R.A. 2005. Aboveground forest biomass and the global carbon balance. Global Change Biology 11:945-958.
Intergovernmental Panel on Climate Change (IPCC). 2001. Summary for policymakers. A report of working group I of the Intergovernmental Panel on Climate Change. Wembley, United Kingdom. 34 p.
Keller, M., M. Palace and G. Hurtt. 2001. Biomass estimation in the Tapajos National Forest, Brazil. Examination of sampling and allometric uncertainties. Forest Ecology and Management 154(3): 371-382.
Laurance, W., L.V. Ferreira, J.M. Rankinand S.G. Laurance. 1998. Rain forest fragmentation and the dynamics of amazonian tree communities. Ecology 79(6): 2032-2040.
Luyssaert, I., M. Inglima, A. Jung, D. Richardson, M. Reichstein, D. Papale, S.L. Piao, E.D. Schulze, L. Wingate, G. Matteucci, L. Aragao, M. Aubinet, C. Beer, C. Bernhofer, K. Black, D. Bonal, J. Bonnefond, J. Chambers, P. Ciais, B. Cook, K. Davis, A.J. Dolman, B. GiElen, M. Goulden, J. Grace, A. Granier, A. Grelle, T. Griffis, T. Grunwald, G. Guidolotti, P.J. Hanson, R. Harding, D.Y. Hollinger, L.R. Hutyra, P. Kolari, B. Kruijt, W. Kutsch, F. Lagergren, T. Laurila, B.E. Law, G. Le Maire, A. Lindroth, D. Loustau, Y. Malhi, J. Mateus, M. Migliavacca, L. Misson, L. Montagnani, J. Moncrieff, E. Moors, J. Munger, E. Nikinmaa, S. Ollinger, G. Pita, C. Rebmann, O. Roupsard, N. Saigusa, M. Sanz, G. Seufert, C. Sierra, M. Smith, J. Tang, R. Valentini, T. Vesala and I. Janssens. 2007. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Global Change Biology 13: 2509-2537.
Mahecha, G. 1997. Fundamentos y metodologías para la identificación de plantas. Proyecto Biopacífico, Ministerio del Medio Ambiente, GEF - PNUD. Lerner Ltda., Bogotá. 282 p.
Malhi, Y., A. Nobre, J. Grace, B. Kruijt, M. Pereira, A. Culf, and S. Scott. 1998. Carbon dioxide transfer over a Central Amazonian rain forest. Journal of Geophysical Research 103: 31593-31612.
Malhi, Y., T. Baker, O. Phillips, S. Almeida, E. Alvarez, L. Arroyo, J. Chave, C. Czimczik, A. Difiore, N. Higuchi, T. Killeen, S. Laurance, W. Laurance, S. Lewis, L. Mercado, A. Monteagudo, D. Neill, P. Nuñez, S. Patiño, N. Pitman, C. Quesada, R. Salomao, J. Silva, A. Lezama, R. Martínez, J. Terborgh, B. Vinceti and J. Lloyd. 2004. The above-ground coarse wood productivity of 104 neotropical forest plots. Global Change Biology 10: 563-591.
Melo, O. y R. Vargas. 2003. Evaluación ecológica y silvicultural de ecosistemas boscosos. Universidad del Tolima, Impresiones CONDE, Ibagué. 235 p.
Mosquera, Y. y C. Sánchez. 2005. Propuesta para la venta de servicios ambientales con participación comunitaria en el Medio Atrato. Chocó - Colombia. Trabajo de grado. Facultad de Ingeniería. Programa de Ingeniería Ambiental. Universidad Tecnológica del Chocó "Diego Luís Córdoba". Quibdó, Chocó, Colombia. 135 p.
Mostacedo, B. y T. Fredericksen. 2000. Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal. Proyecto Bolfor. Editora El País, Santa Cruz, Bolivia. 82 p.
Murillo, W., R. Palomino, S. Córdoba, G. Aragón y E. Banguero. 2005. El régimen diario de la precipitación en el municipio de Quibdó (Colombia). Revista de Climatología 5: 1-7.
Odum, E. 1969. The strategy of ecosystem development. An understanding of ecological succession provides a basis for resolving mans conflict with nature. Science, New Series 164(3877): 262-270.
Orrego, S.A. y J.I. Del Valle. 2003. Existencias y tasas de incremento neto de la biomasa y del C en bosques primarios intervenidos y secundarios. pp. 215-242. En: Orrego, S.A., J.I. Del Valle y F.H. Moreno (eds.). Medición de la captura de C en ecosistemas forestales de Colombia: Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia, Sede Medellín, Departamento de Ciencias Forestales y Centro Andino para la Economía en el Medio Ambiente, Bogotá. 314 p.
Orrego, S.A., J.I. Del Valle y F.H. Moreno. 2003. Medición de la captura de C en ecosistemas forestales de Colombia: Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia, Sede Medellín, Departamento de Ciencias Forestales y Centro Andino para la Economía en el Medio Ambiente, Bogotá. 314 p.
Palacios, J., Y. Ramos, A. Mosquera, F.L. Castro, F. García, J. Arroyo y A. Cogollo. 2003b. Estructura de un bosque pluvial tropical (bp-T) en Salero, Unión Panamericana, Chocó. pp. 45-61. En: García, F., Y. Ramos, J. Palacios, J.E. Arroyo, A. Mena y M. González. Salero diversidad biológica de un bosque pluvial tropical (bp-T). Editorial Guadalupe Ltda, Bogotá. 207 p.
Phillips, O.L., Y. Malhi, N. Higuchi, W. Laurance, P. Núñez, M. Vásquez, S. Laurance, L. Ferreira, M. Stern, S. Brown and J. Grace. 1998. Changes in the carbon balance of tropical forests: Evidence from long-term plots. Science 282: 439-442.
Quinto, H., Y. Ramos y D. Abadía. 2007. Cuantificación de la caída de hojarasca como medida de la productividad primaria neta en un bosque pluvial tropical en Salero, Unión Panamericana, Chocó, Colombia. Revista Institucional Universidad Tecnológica del Chocó D.L.C. 26: 28-41.
Quinto, H. y Y. Ramos. 2009. Descomposición de hojarasca en un bosque pluvial tropical intervenido en el municipio de Lloró, Chocó, Colombia. Revista Institucional Universidad Tecnológica del Chocó D.L.C. 28: 32-43.
Quinto, H. y F.H. Moreno. 2010. Dinámica de la biomasa aérea en bosques primarios de Colombia y su relación con la precipitación y la altitud. Tesis de Posgrado en Bosques y Conservación Ambiental. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 75 p.
Rodríguez, J.C. 1989. Consideraciones sobre la biomasa, composición química y dinámica del bosque pluvial tropical de colinas bajas. Bajo Calima. Buenaventura, Colombia. Corporación Nacional de Investigación y Fomento Forestal - CONIF, Bogotá, D.E. 36 p. (Serie Documental No. 16).
Sarmiento, G., M. Pinillos and I. Garay. 2005. Biomass variability in tropical american lowland rainforests. Ecotropicos 18(1): 1-20.
Schuur, E.A.G. 2003. Productivity and global climate revisited: the sensitivity of tropical forest growth to precipitation. Ecology 84(5): 1165-1170.
Sierra, C.A., J.I. Del Valle, S.A. Orrego, F.H. Moreno, M.E. Harmon, M. Zapata, G.J.Colorado, M.A. Herrera, W. Lara, D.E. Restrepo, L.M. Berrouet, L.M. Loaiza, and J.F. Benjumea. 2007. Total carbon stocks in a tropical forest landscape of the Porce region, Colombia. Forest Ecology and Management 243(2-3): 299-309.
Statistical Graphics Corporation. 2002. Statgraphics Plus Version 5.1, www.statgraphics.com.
Swaine M.D. and J.B. Hall. 1983. Early succession on cleared forest land in Ghana. Journal of Ecology 71(2): 601-627.
Turner, I.M. 2001. The ecology of trees in the tropical rain forest. First publisher. Cambridge University Press, Cambridge. 298 p.
Universidad Nacional de Colombia, Sede Medellín. (UNALMED). 2001. Evaluación de las plantaciones de Acacia mangium Willd en áreas degradadas del Bajo Cauca. Informe Técnico. Universidad Nacional de Colombia - CORANTIOQUIA, Medellín. 325 p.
Vallejo, M., A.C. Londoño, R. López, G. Galeano, E. Álvarez, y W. Dévia. 2005. Establecimiento de parcelas permanentes en bosques de Colombia (métodos para estudios ecológicos largo plazo). Instituto de Investigaciones de Recursos Biológicos Alexander von Humboldt, Bogotá D.C. 310 p.
Whitmore, T.C. 1975. Tropical rain forest of the far east. Oxford University Press. Scotland. Reino Unido. 260 p.
Whitmore, T.C. 1998 Anintroduction to tropical rain forests. Second edition. Oxford University Press, Oxford. 226 p.
Yepes, A. 2008. Dinámica de un bosque primario premontano de los Andes Colombianos. Tesis de Posgrado en Bosques y Conservación Ambiental. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 75 p.
DINÁMICA DE LA BIOMASA AÉREA EN UN BOSQUE PLUVIAL TROPICAL DEL CHOCÓ BIOGEOGRÁFICO
DYNAMICS OF TREE ABOVEGROUND BIOMASS IN A TROPICAL RAIN FOREST OF THE CHOCÓ BIOGEOGRÁFICO
Harley Quinto Mosquera1 y Flavio Moreno Hurtado2
1 Docente Ocasional. Universidad Tecnológica del Chocó "Diego Luis Córdoba". Facultad de Ciencias Básicas. Barrio Nocolás Medrano, Ciudadela Universitaria. A.A. 292, Quibdó, Colombia. <hquintom@unal.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <fhmoreno@unal.edu.co>
Recibido: Febrero 02 de 2010; aceptado: Abril 18 de 2011
Resumen. El estudio de la biomasa aérea (BA) de los bosques tropicales es fundamental para entender el balance del C global en el contexto del cambio climático. La BA se cuantificó en un bosque maduro de Salero (Chocó Biogeográfico), mediante ecuaciones diseñadas para bosques húmedos tropicales, a partir de datos de densidad de madera, diámetro (DAP) y altura de árboles (con DAP = 10 cm) medidos en dos sub-parcelas permanentes ("E" y "U") de 1 ha, las cuales se monitorearon en los años 1998, 2005 y 2008. La BA fue 237,31 t·ha-1, 259,99 t·ha-1 y 217,97 t·ha-1 respectivamente en la sub-parcela "E". Mientras que en la "U" fue de 178,94 t·ha-1y 179,17 t·ha-1 en los años 2005 y 2008; las diferencias de BA a través del tiempo fueron no significativas. Los incrementos promedios anuales de BA de sobrevivientes fueron 4,42 y 3,18 t·ha-1 año-1 en las sub-parcelas "E" y "U" respectivamente. Además, en sub-parcela "E" en condiciones imperturbadas, se presentó una tasa de incremento neto de la BA (TINBA) de 2,61 t·ha-1 año-1, en concordancia con la hipótesis del incremento en la BA en los bosques húmedos tropicales. La productividad primaria neta aérea (PPNA) en Salero de carbono fue de 5,21 t· ha-1 año-1, por lo tanto los resultados no apoyaron la hipótesis de la disminución en la productividad de los bosques tropicales con el incremento en la precipitación.
Palabras clave: Cambio climático global, C, tasa de incremento neto de la BA (TINBA), productividad primaria neta (PPN), parcelas permanentes.
Abstract. The study of the aboveground biomass (AB) of tropical forests is fundamental to understand the balance of the global C in the context of the climatic change. We quantified the AB in a mature forest of Salero (Chocó Biogeográfico), by means of equations designed for tropical humid forests, starting from data of wooden density, diameter (D) and height of trees (with D = 10 cm) measured in two permanent sub-parcels (E and U) of 1 hectare (ha), which were measured in the years 1998, 2005 and 2008. Inthis years the AB was of 237.31 t·ha-1, 259.99 t·ha-1 and 217.97 t·ha-1 respectively in the E sub-parcel. While in the U it was of 178.94 t·ha-1 and 179.17 t·ha-1 in the years 2005 and 2008; the differences of the aboveground biomass through the time were not significant. The increments annual averages in survivors' BA were of 4.42 and 3.18 t·ha-1 year-1 in the E and U sub-parcels respectively. Also, in sub-parcel E under conditions imperturbadas, the rate of net increment was presented in the AB (TINBA) of 2.61 t·ha-1 year-1, in agreement with the hypothesis of the increment in the BA in the tropical humid forests. The PPNA of carbon in Salero was of 5.21 t·ha-1 year-1, therefore the results didn't support the hypothesis of the decrease in the productivity of the tropical forests with the increment in the precipitation.
Key words: Change climatic global, carbon, net increment of the aboveground biomass, net primary productivity, permanent plot.
Los bosques tropicales son importantes en el balance del carbono (C) global, debido a que representan cerca del 59% del C total acumulado en los ecosistemas boscosos del mundo (Dixon et al., 1994). Dado que estos ecosistemas capturan CO2 atmosférico mediante el proceso de fotosíntesis y lo acumulan en sus tejidos (Chazdon y Montgomery, 2002; Clark et al., 2001a), el estudio de la dinámica de la biomasa y el C es fundamental para comprender el papel que éstos desempeñan en el ciclo global de este elemento (Clark et al., 2001a; Sierra et al., 2007) y en la mitigación del cambio climático (Brown, 1997; IPPC, 2001). En consecuencia, existe gran interés por evaluar las existencias, los patrones de captura y el balance de C de los bosques tropicales del mundo con el objetivo de develar si estos ecosistemas están o no acumulando C atmosférico adicional (Brown, 1997; Phillips et al., 1998; Malhi et al., 1998; Clark et al., 2001a; IPPC, 2001; Clark, 2002; Chave et al., 2003; Baker et al., 2004a; Houghton, 2005). Recientemente, estos estudios han cobrado mayor importancia por la necesidad de estimar las cantidades de C que podrían ser emitidas a la atmósfera en caso de deforestación (Houghton, 2005), lo cual es un insumo fundamental para la estructuración de proyectos de reducción de emisiones por deforestación y degradación (REDD) (Angelsen et al., 2009).
Según la teoría ecológica clásica (Odum, 1969; Whitmore, 1975; Bormann y Likens, 1979), los bosques naturales primarios imperturbados o climáxicos no acumulan C atmosférico adicional, ni presentan cambios netos en su biomasa total, pues se mantienen en equilibrio dinámico durante largos períodos de tiempo. Sin embargo, varios estudios realizados en la Amazonia han documentado que tales bosques sí están capturando C atmosférico en forma significativa, como lo evidencian los resultados de Phillips et al. (1998); Malhi et al. (1998); Baker et al. (2004b) y Malhi et al. (2004) en 104 parcelas neotropicales. Este comportamiento se debe posiblemente al aumento de CO2 atmosférico, el cual probablemente ha estado funcionando como fertilizante natural en las últimas décadas (Phillips et al., 1998; Malhi et al., 1998, 2004; Baker et al., 2004b).
No obstante, estos trabajos han sido fuertemente criticados, debido a la ocurrencia de posibles errores metodológicos (Clark, 2002); adicionalmente, no se han registrado tendencias similares en los bosques tropicales de Asia y África (Houghton, 2005). Feeley et al. (2007) encontraron que en los bosques maduros de Malasia y Barro Colorado (Panamá) se presentaron disminuciones en las tasas de crecimiento de un alto porcentaje de las especies, debido especialmente a la disminución en la precipitación total y a incrementos en la temperatura mínima diaria. Estos resultados contradicen la hipótesis de incremento de la productividad como lo plantean los estudios de la Amazonia citados atrás. En consecuencia, el debate está lejos de concluir, y al contrario, crece la incertidumbre sobre la capacidad de captura de C adicional por los bosques tropicales de cara al cambio climático.
De otro lado, la relación entre captura de C y precipitación en bosques tropicales es confusa principalmente porque son pocos los estudios que han analizado dicha relación y sus resultados son a veces contradictorios (Brown y Lugo, 1982; Clark y Clark, 1994; Schuur, 2003; Malhi et al., 2004). Por ejemplo, Schuur (2003) afirmó que la productividad disminuye notablemente cuando la precipitación alcanza valores superiores a 5.000 mm anuales, debido a factores como la disminución de nutrientes en el suelo, incremento de la lixiviación, aumento de la nubosidad y reducción de la tasa de descomposición de la materia orgánica. Malhi et al. (2004) por su parte, reportaron que la productividad de madera gruesa aérea no tuvo una relación obvia con la precipitación. Recientemente un análisis mundial sobre el balance de CO2 (Luyssaert et al., 2007), muestra que la productividad incrementa con el aumento de la precipitación, hasta que la precipitación alcanza valores de 1.500 mm anuales; donde sobrepasa estos valores la relación no es clara. Más aún, la productividad de los bosques tropicales ha sido pobremente estudiada cuando los niveles de precipitación sobrepasan los 5.000 mm anuales.
En Colombia, desde hace varios años se ha venido estableciendo una red de parcelas permanentes con el objetivo de conocer la dinámica y los patrones de distribución de especies en los bosques del país (Vallejo et al., 2005). Esta red representa una oportunidad excelente para probar hipótesis y mejorar la comprensión sobre el ciclo del C en los ecosistemas colombianos. No obstante, sólo en unos pocos sitios existen datos de más de cinco años de medición, entre los que se destacan los estudios realizados en la cuenca media del río Porce entre los años 1999 y 2006 (Orrego y Del Valle, 2003; Orrego et al., 2003; Yepes, 2008).
Con el ánimo de contribuir al entendimiento de la productividad del bosque y su relación con factores como la precipitación, se planteó el objetivo de estimar la biomasa aérea (BA) y su dinámica en el tiempo durante un período de 10 años en un bosque pluvial tropical del departamento del Chocó, respondiendo los siguientes interrogantes: ¿Cómo se distribuye la BA en función de las especies, las familias botánicas y los grupos funcionales? ¿Cómo varía la BA en un bosque pluvial tropical primario a través del tiempo? ¿Cómo se compara la productividad primaria neta aérea de este bosque con la de otros bosques tropicales sometidos a condiciones de precipitación más moderadas?
Área de Estudio. El estudio se realizó en una parcela permanente de investigación ubicada en un bosque pluvial tropical maduro de tierras bajas localizado en el corregimiento de Salero, municipio de Unión Panamericana, departamento del Chocó, Colombia, en las coordenadas 5º18'950'' de Latitud Norte y a los 76º36'742'' de Longitud Oeste (Figura 1). Este bosque se encuentra en la zona de vida de bosque pluvial tropical (bp-T) (Holdridge, 1982); la precipitación promedia anual de la región es cercana a 10.000 mm, la temperatura promedia varía entre 26 ºC y 30 ºC, la humedad relativa es de 90% y la altura sobre el nivel del mar oscila entre 100-150 m (García et al., 2003).
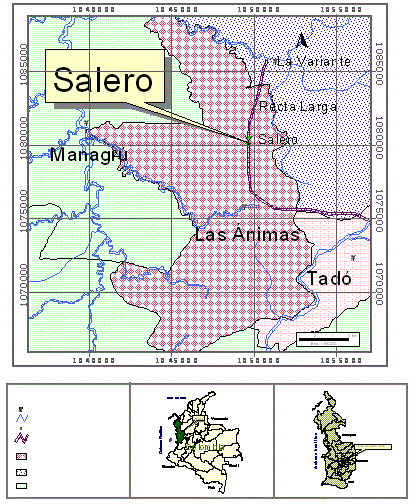
Figura 1. Ubicación de la parcela permanente de investigación en biodiversidad en el corregimiento de Salero, Unión Panamericana, Chocó, Colombia.
Los bosques de Salero son primarios, con muy poca intervención antrópica y con una alta diversidad de especies de plantas vasculares (Palacios et al., 2003). En este bosque se instaló una parcela permanente de investigación (PPIB) localizada al nororiente de la comunidad de Salero, desde la cual se tarda hora y media por trocha para llegar al sitio de la parcela, denominado localmente Catangüero. Este último se encuentra en el margen derecho de la quebrada Candelaria, la cual recoge las aguas de los riachuelos que escurren de la PPIB, entre ellos el Catangüero. En estos bosques el terreno es disectado, con diversos grados de pendiente; allí es común encontrar árboles caídos debido a distintos procesos de su dinámica natural.
Vegetación. Las familias más abundantes y frecuentes que caracterizan este ecosistema son Sapotaceae, Lecythidaceae, Arecaceae, Euphorbiaceae, Mirysticaceae, Chrysobalanaceae, Moraceae, Clusiaceae, Caesalpiniaceae, Vochysiaceae, Apocynaceae, Fabaceae y Bombacaceae (Asprilla et al., 2003). Entre las especies predominantes están el juanchito (Mabea chocoensis), palma mil pesos (Jessenia bataua), lechero (Brosimum utile), vaina (Matisia bullata), guasco (Eschweilera pittieri), palma memé (Wettinia quinaria), carbonero (Licania sp.), guayabo (Croton jorgei) y caimito (Chrysophyllum sp.), entre otras. (Palacios et al., 2003).
MÉTODOS
Establecimiento y re-censos de la parcela permanente. Generalmente para el estudio de la dinámica del C en los bosques se han empleado parcelas permanentes de investigación (Phillips et al., 1998; Clark et al., 2001; Del Valle et al., 2003; Chave et al., 2003; Baker et al., 2004a), las cuales permiten conocer los cambios en las existencias de biomasa y C en el bosque cuando se realizan mediciones sucesivas a lo largo de varios años. En este estudio se empleó una parcela permanente de investigación establecida entre abril y agosto del año 1998 mediante la metodología de BIOTROP (Melo y Vargas, 2003), y actualmente hace parte de la red nacional de parcelas permanentes de Colombia (Vallejo et al., 2005).
La parcela consiste en un cuadrado de 500x500 m (25 ha), dividido en 25 sub-parcelas rectangulares de 500x20 m (1 ha), que se identifican con las letras del abecedario desde la "A" hasta la "Y". Cada una de estas sub-parcelas está dividida en 25 cuadrantes de 20x20 m (400 m2) y a su vez, cada uno de los cuadrantes está dividido en cuatro cuadrículas de 10x10 m (100 m2) (García et al., 2003). El inventario de los árboles con DAP=10 cm se realizó en las sub-parcelas "E" y "U" de la parcela (Figura 2). Entre junio y septiembre del año 2005, y entre julio y agosto del año 2008 se realizaron el segundo y tercer censo de la parcela. En cada uno de los censos se registraron los individuos muertos, la forma de mortalidad y los individuos reclutados (que alcanzaron el DAP=10 cm).
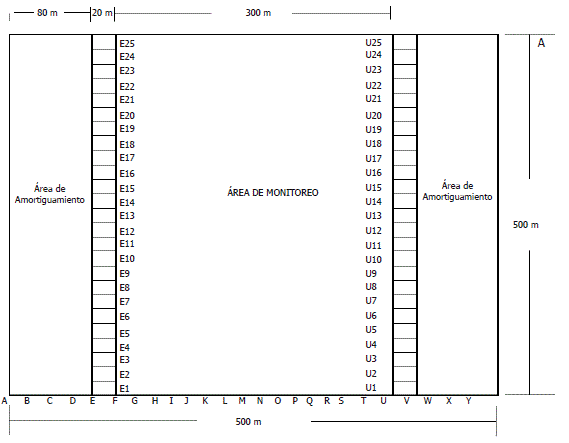
Figura 2. Esquema de la parcela permanente de investigación en biodiversidad de Salero (Chocó, Colombia).
Identificación botánica. Se identificaron todas las morfoespecies hasta el máximo nivel taxonómico posible (NN, especie, género, familia botánica) en el herbario de la Universidad Tecnológica del Chocó "D.L.C." "Herbario Chocó". Esta identificación se hizo utilizando las claves especializadas de Forero y Gentry (1989), Gentry (1993) y Mahecha (1997). Además se contó con la colaboración del Biólogo Álvaro Cogollo del Jardín Botánico "Joaquín Antonio Uribe" de Medellín en la identificación del material.
Clasificación en grupos ecológicos de especies. Las especies identificadas taxonómicamente en las sub-parcelas "E" y "U", fueron clasificados de acuerdo a sus requerimientos de luz en climáxicas y pioneras, acorde con lo propuesto por Whitmore (1998). Además se empleó información de varios listados e inventarios de especies que incluían las características ecológicas de éstas. Entre los listados empleados están los citados por Swaine y Hall (1983); Whitmore (1998); Turner (2001); Unalmed (2001) y Corantioquia (2006), y los inventarios realizados por Córdoba et al. (2005) en claros naturales en bosques del Chocó, en los que se mencionan principalmente las especies pioneras o demandantes de luz. Para los individuos que se encontraban sin identificar taxonómicamente en la parcela de Salero (94 árboles), se asumió que pertenecían a especies climáxicas, en consideración a que éstas se encuentran en mayor proporción en los bosques primarios poco intervenidos; además las especies pioneras suelen ser comunes y por tanto fácilmente identificables.
Medición de diámetros en las sub-parcelas. Se midió la circunferencia a la altura del pecho en cm (1,30 m sobre el nivel del suelo) con cinta métrica a todos los árboles con DAP=10 cm en cada cuadrícula; posteriormente, los valores de circunferencia se transformaron a DAP. El perímetro del tronco del árbol donde se midió el diámetro por primera vez se marcó con pintura asfáltica amarilla con el objeto de garantizar que posteriores mediciones se hagan en la misma zona de la primera medición. Tales mediciones se hicieron en áreas libres de nudos o ramas. Además, se registró la ubicación de cada individuo en la cuadrícula. Todos los árboles medidos fueron marcados con placas de aluminio. También se identificó el hábito de crecimiento en las categorías de árbol, bejuco o liana y palma, se anotaron las características vegetativas y observaciones particulares de cada individuo.
Modelación de la altura total y comercial. Para modelar las alturas de los árboles se midió la altura total a 471 individuos, de los cuales 72 fueron árboles climáxicos medidos en la parcela y 399 individuos pioneros registrados en una zona alterada cercana a la parcela. Para estas mediciones se empleó el clinómetro Suunto® a distancias fijas de 15 y 20 m de observación. Para obtener la altura total de cada árbol se tuvo en cuenta el ángulo a la cima de la copa y la base del árbol, la distancia de observación y la altura de la persona que realiza la medición (Mostacedo y Fredericksen, 2000).
Con la información de la altura total, el DAP y el grupo ecológico de los 471 árboles medidos directamente en campo, se generaron modelos lineales de altura total en función del DAP. Con base en dichos modelos se estimó la altura a los demás árboles, teniendo en cuenta su grupo funcional. Estos datos de altura total fueron necesarios para estimar la biomasa aérea en algunos modelos. Por otro lado, dado que uno de los modelos empleados para estimar la biomasa aérea de los árboles requiere de la altura comercial (definida como aquella que va desde el suelo hasta el inicio de las ramas grandes de la copa del árbol) para su estimación y, debido a que esta altura no fue medida directamente en campo, se generó un modelo para estimarla. Para elaborar el modelo se empleó una base de datos de la Amazonia que incluía valores de altura (comercial y total) de especies como Coussarea sp., Posoqueria sp. Sterculia sp., Pouteria sp., Euterpe precatoria, Mabea sp., entre otras (Cuartas, 1993), que también han sido registradas en Salero (Palacios et al., 2003).
Densidad específica de la madera. Los valores de esta variable se tomaron de una base de datos internacional creada principalmente para bosques de la Amazonia (Baker et al., 2004 a); en los casos en los que alguna especie o género encontrado en Salero no se encontrara reportado en esta base de datos, se empleó el promedio del género o de la familia de la especie; para los individuos indeterminados taxonómicamente se utilizó el promedio general de densidad de la parcela de Salero.
Estimación de la biomasa aérea del bosque. Dado que la elección de un modelo o ecuación para estimar la BA de árboles es crucial, porque puede ser una de las principales fuentes de error en la estimación (Clark et al., 2001; Sarmiento et al., 2005), se recomienda emplear ecuaciones obtenidas localmente (Araujo et al., 1999; Chambers et al., 2001; Del Valle et al., 2003). Sin embargo, cuando se carece de ecuaciones locales se pueden emplear ecuaciones ya existentes, ideadas en condiciones climáticas y edáficas similares a la de la zona de estudio, que pueden dar resultados muy confiables (Clark et al., 2001), especialmente cuando las mediciones realizadas en campo con anterioridad son similares a las estimaciones del modelo. En este estudio, se cuantificó la BA de los árboles mediante siete modelos (Tabla 1) construidos con información de bosques húmedos tropicales de diversas zonas del mundo (Rodríguez, 1989; Brown, 1997; Chamberset al., 2001; Baker et al., 2004a; Chave et al., 2005). Se tomó como unidad de muestreo a los cuadrantes de 20 x 20 m2 de las sub-parcelas.
Tabla 1. Ecuaciones empleadas para estimar la BA en Salero (Chocó, Colombia). BA es la biomasa aérea de los árboles en kg, DAP es el diámetro en cm, Ln es el logaritmo neperiano, H es la altura total en m, Hc es la altura comercial en m, pi es la densidad de la madera.
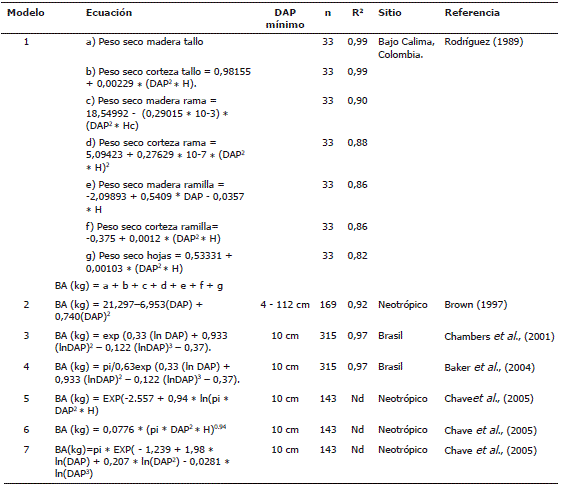
El modelo de Rodríguez (1989) fue creado localmente en bosques del Chocó biogeográfico; para la estimación de la BA se usa DAP, altura total y altura comercial. Los modelos de Brown (1997) y Chambers et al. (2001) por su parte, solo incluyen DAP. Por último, los modelos de Baker et al. (2004a) y Chave et al. (2005) estiman la BA en función del DAP, altura total y densidad de la madera. Se examinaron las estimaciones de BA total con cada una de las ecuaciones y en cada uno de los censos realizados en la parcela. Para los análisis de BA a través del tiempo y en grupos ecológicos, se seleccionó el modelo con mayor coeficiente de correlación promedio en relación con las estimaciones de los otros modelos, como lo han realizado otros investigadores (Chave et al., 2003).
Dinámica de la biomasa aérea. Con la información de los inventarios de los años 1998, 2005 y 2008, se estimó la BA total a través del tiempo, las pérdidas por mortalidad de árboles, los incrementos por crecimiento y reclutamiento de árboles, y la tasa de incremento neto de la BA (TINBA), mediante la ecuación propuesta por Beers (1962), la cual consiste en: TINBA = ?s + I - C - M. En donde ?s corresponde al cambio en BA de los árboles sobrevivientes, calculada con la BA final (segunda medición) menos la BA inicial (primera medición) de cada individuo sobreviviente; I es la BA de los árboles reclutados (individuos nuevos que alcanzaron = 10 cm de DAP) registrados en la segunda medición; C es la BA de los árboles cortados y M es la BA de los árboles muertos entre las dos mediciones. Todas las estimaciones de BA fueron realizadas en t ·ha-1.
Por otro lado, en el periodo comprendido entre 2005 y 2008 se presentaron talas selectivas de árboles en ambas sub-parcelas, debido a esto los análisis de la variación de la BA se realizaron excluyendo e incluyendo los valores de BA de los arboles deforestados en cada sub-parcela, tomando como referencia el valor en el año 2005. En total, se registraron 186 y 97 individuos deforestados en las sub-parcela E y U respectivamente.
Análisis de datos. Se calcularon las medidas de tendencia central (media y mediana) y de dispersión (desviación estándar, coeficiente de variación y error estándar) de los datos. Mediante el coeficiente de correlación de Pearson se evaluó la correlación entre las distintas estimaciones de BA generadas con los siete modelos. Así mismo, se evaluaron los supuestos de normalidad y homogeneidad de varianzas con los estadísticos de Bartlett's y Hartley's; cuando los datos fueron normales y se cumplieron los supuestos se realizó un análisis de varianza ANAVA (Hoshmand, 1998). Cuando no se cumplieron los supuestos del ANAVA se empleó el estadístico de Kruskal Wallis para evaluar la existencia de diferencias significativas entre las medianas de la BA en función de los períodos de medición (1998, 2005 y 2008), sub-parcelas, cuadrantes, y grupo ecológico de especies. Mediante la prueba de rangos múltiples de Bonferroni se evaluó entre cuáles grupos se presentaron diferencias significativas de BA. Dado que en el proceso de depuración de la base de datos de campo se encontró que cerca del 30% de la información del DAP de 1998 en la sub-parcela "U" presentaba inconsistencias con relación con las mediciones de 2005 y 2008, se optó por excluir dichos datos del análisis general de la BA.
Para probar la hipótesis de la disminución de la PPN de los bosques tropicales con el incremento en la precipitación (Schuur, 2003), se estimó la productividad primaria neta aérea (PPNA) de la siguiente forma: PPNA= ?s + PH. Donde ?s es el incremento en BA de árboles sobrevivientes y PH es la producción de hojarasca. La producción de hojarasca (7,2 t·ha-1·año-1) de Salero empleada, fue la publicada por Quinto et al. (2007). En la estimación de la PPNA no fueron incluidas las pérdidas por consumidores y de compuestos orgánicos volátiles que hacen parte de la PPN porque representan proporciones poco significativas de la PPN total y generalmente no son medidas (Clark et al. 2001a).
Con la información de la base de datos de Clark et al. (2001b) incluidos los datos de Salero y el Bajo Calima, se establecieron rangos de precipitación promedio anual (de 500-2.000, 2.000-4.000, 4.000-6.000, 6.000-8.000 y de 8.000-10.000 mm anuales) con los cuales se realizó una prueba de Kruskal-Wallis para evaluar las diferencias significativas entre las medianas de la PPNA de cada rango de precipitación, y mediante el estadístico de rangos múltiples de Bonferroni se evaluaron los posibles grupos al nivel de los rangos de precipitación con relación a la PPNA (Guttman, 1982). Todos los análisis de los datos se realizaron con el programa Statgraphics Centurion XVI (Statistical Graphics Corporation, 2002).
RESULTADOS
Modelación de las alturas de los árboles en el bosque pluvial tropical de Salero, Chocó. Los modelos lineales generados para estimar la altura total y comercial en función del DAP, de las especies arbóreas climáxicas y pioneras del bosque pluvial de Salero (Chocó) se detallan en la Tabla 2. Todos los modelos fueron significativos y variaron en el tamaño de muestra y en el coeficiente de determinación (Tabla 2).
Tabla 2. Modelos lineales de la altura total y comercial de las especies arbóreas pioneras y climáxicas del bosque de Salero (Chocó, Colombia). En los modelos Ht es la altura total en m. Hc es la altura comercial en m. DAP es el diámetro a la altura del pecho en cm. E.E. es el Error estándar de los coeficientes. r2 es el Coeficiente de determinación. N. árboles muestreados. E.A.P. Error absoluto promedio.
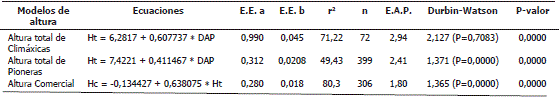
Estimaciones de biomasa aérea en el bosque pluvial tropical de Salero, Chocó. Las estimaciones de BA de árboles realizadas con los siete modelos empleados presentaron diferencias estadísticamente significativas entre ellos (F= 42,98; P<0,05). Mediante la prueba de rangos múltiples de Bonferroni se encontró que los valores de los modelos 4, 5, 6 y 7 fueron estadísticamente similares. Las mayores estimaciones de BA se presentaron con el modelo 3, con 293,25 t·ha-1 promedio; mientras que con el modelo 1 se presentaron las menores estimaciones de BA, con 157,27 t·ha-1 promedio (Tabla 3).
Tabla 3. Estimaciones de biomasa aérea en la parcela permanente de investigación de Salero (Chocó, Colombia), con los siete modelos de estimación. El modelo 1 fue publicado por Rodríguez (1989), el modelo 2 por Brown (1997), el modelo 3 por Chambers et al. (2001), el modelo 4 por Baker et al. (2004) y los modelos 5, 6 y 7 por Chave et al. (2005).
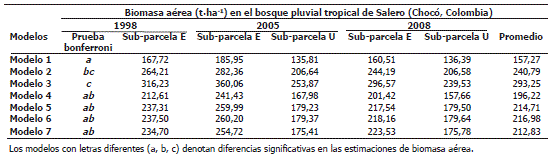
Las estimaciones de BA de los modelos fueron diferentes con relación a los diámetros de los árboles. Dichas diferencias fueron menores en los árboles pequeños y se hicieron cada vez mayores en los árboles más grandes, especialmente en los modelos que emplean la densidad de la madera y la altura total de los árboles en sus cálculos (Figura 3). Sin embargo, las variaciones en la BA entre individuos influyeron de forma significativa sobre las estimaciones de BA total, a pesar de que dichas variaciones se compensaron entre sí en algunas de las estimaciones de BA al nivel de bosque (Tabla 3).
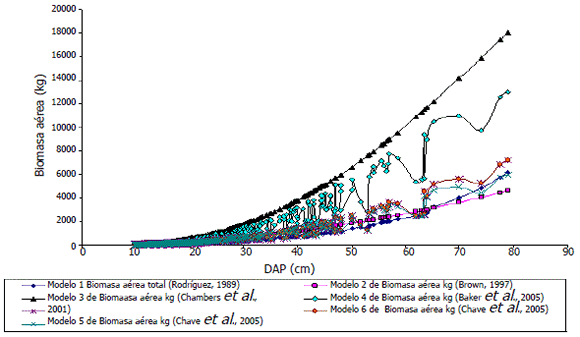
Figura 3. Estimación de la biomasa aérea de árboles en función del DAP para los siete modelos empleados. Estimación realizada con los datos del año 2005 de la sub-parcela "E" del bosque pluvial tropical de Salero (Chocó, Colombia).
Se registraron correlaciones significativas entre la BA estimada con los distintos modelos. Los modelos 5 y 6 presentaron las mayores correlaciones (0,984%), debido a esta alta correlación se optó por elegir el modelo 5 para realizar las evaluaciones de BA de los árboles. Por otro lado, el modelo 1 mostró la menor correlación (0,965%) (Tabla 4).
Tabla 4. Matriz de correlación de Pearson de los siete modelos de biomasa aérea empleados. Las correlaciones se realizaron con la BA de los individuos registrados en el año 2005 en la sub-parcela "E" en Salero (Chocó, Colombia). La última fila muestra el promedio de correlación de cada modelo. 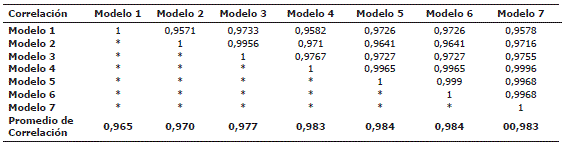
Distribución de la BA en las sub-parcelas de Salero, Chocó. En el año 1998 en la sub-parcela "E" se registraron en total 709 individuos. La BA estimada a partir de los cuadrantes de 20x20 m fue en promedio (± desviación estándar) de 237,31 ±101,61 t·ha-1. Así mismo, la BA en la sub-parcela "E" fue de 259,99 ± 109,25 t·ha-1, y de 179,23 ± 89,33 t·ha-1 en la sub-parcela "U" en el año 2005. En dicha medición se determinaron diferencias significativas (T= 7,43; P= 0,006) entre las medianas al nivel de las sub-parcelas. =5,28; P=0,021) en la BA total entre sub-parcelas en la medición del año 2008, cuando fueron incluidos los valores de BA de los árboles deforestados (Tabla 5).
Tabla 5. Estimaciones de biomasa aérea en el bosque pluvial de Salero (Chocó, Colombia). Las pruebas de hipótesis de varianzas fueron realizadas con la prueba Kruskal-Wallis. D.E. Desviación estándar.
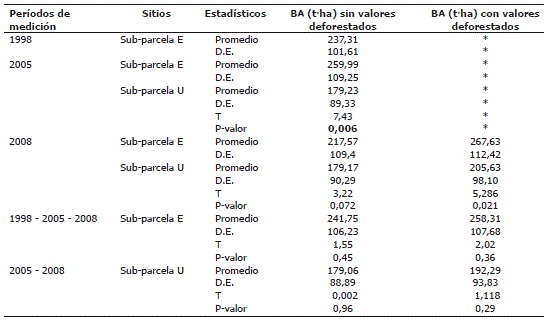
Las estimaciones de BA total de los años 1998, 2005 y 2008 de la sub-parcela E, no mostraron diferencias significativas en sus medianas. Así mismo, la BA total de los años 2005 y 2008 de la sub-parcela U, no presentó diferencias significativas. No obstante, las estimaciones de BA total expresaron una tendencia al incremento con el paso del tiempo cuando fueron incluidos los valores de los árboles deforestados; aunque, dichas evaluaciones no evidenciaron diferencias significativas en sus medianas (Tabla 5).
En las especies, se encontró que Pouteria sp1, Eschweilera pittieri, Eschweilera sp, Brosimum utile, Qualea sp1 y Tetragastris panamensis fueron las que registraron mayor BA en las sub-parcelas en todos los períodos de medición. En todas las evaluacciones la especie más abundante fue Pouteria sp1 (Tabla 6). La distribución de la BA a nivel de los géneros mostró que Pouteria, Eschweilera, Licania, Brosimum, Tetragastris y Qualea fueron los que mayor BA registraron (Tabla 7). Por su parte, las familias Sapotaceae, Lecythidaceae, Chrysobalanaceae, Burseraceae, Moraceae, Myristicaceae y Vochysiaceae fueron las de mayor BA (Tabla 8). Cabe mencionar que Euphorbiaceae y Arecaceae, dos familias abundantes en los bosques estudiados, con 58 y 59 individuos respectivamente, presentaron baja BA (6,7 y 5,7 t·ha-1 respectivamente). La distribución de la BA en los niveles de especie, género y familias botánicas fue similar en todas las mediciones (Tablas 6, 7 y 8).
Tabla 6. Estimaciones de la biomasa aérea al nivel de especies en la parcela permanente de investigación de Salero (Chocó, Colombia). n es el número de individuos registrados.
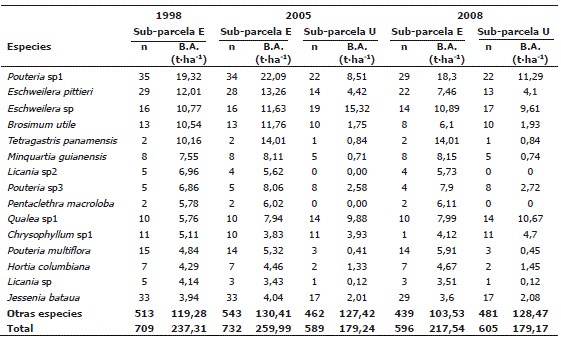
Tabla 7. Estimaciones de la biomasa aérea al nivel de géneros en la parcela permanente de investigación de Salero (Chocó, Colombia). n es el número de individuos registrados.
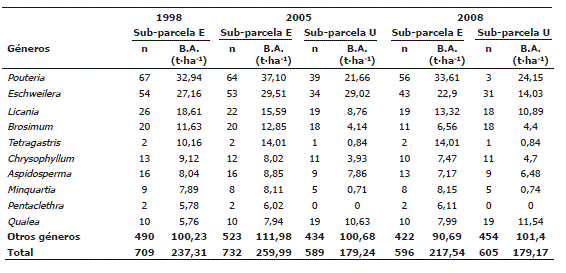
Tabla 8. Estimaciones de la biomasa aérea al nivel de familias botánicas en la parcela permanente de investigación de Salero (Chocó, Colombia). n es el número de individuos registrados.
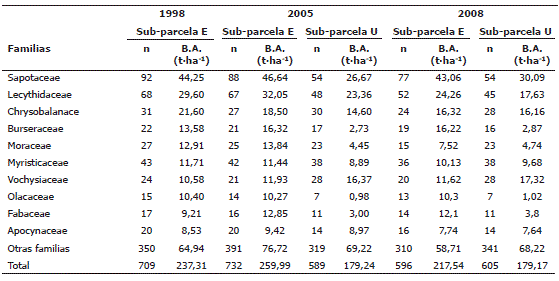
Al nivel de los cuadrantes, en la sub-parcela "E" en los años 2005 y 2008 el intervalo de clase modal (cerca del 32% en el 2005 y del 45% en el 2008) indicó una BA de entre 170 t·ha-1 y 270 t·ha-1. Mientras que en la sub-parcela "U" el intervalo modal (el 36% de los cuadrantes) presentó una BA entre 65 t·ha-1 y 137,5 t·ha-1 en el año 2005, y de entre 125 t·ha-1 y 200 t·ha-1 (el 36% de los cuadrantes) en el año 2008 (Figura 4).
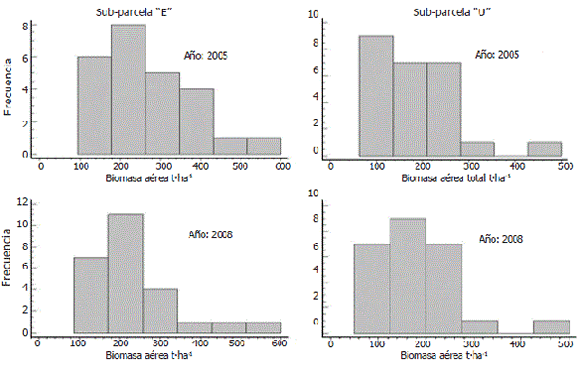
Figura 4. Histograma de frecuencia de la biomasa aérea al nivel de cuadrantes en las sub-parcelas "E" y "U" en los años 2005 y 2008 en el bosque pluvial tropical de Salero (Chocó, Colombia).
En los grupos ecológicos de especies, la mayor BA y cantidad de individuos se manifestó en las especies climáxicas en todos los períodos de medición. En el año 2005, la BA de las pioneras fue mayor en la sub-parcela "E" que en la sub-parcela U, a pesar de exhibir una cantidad similar de individuos (156 en la "E" y 152 en la "U") (Tabla 9).
Tabla 9. Distribución de la biomasa aérea en grupos ecológicos de especies en las sub-parcelas "E" y "U" en los años 1998, 2005 y 2008 en el bosque pluvial tropical de Salero (Chocó, Colombia).
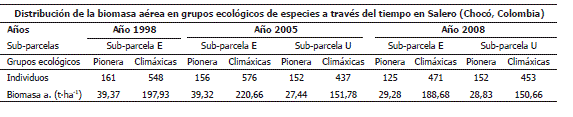
Dinámica de la BA en el bosque pluvial tropical en Salero, Chocó. En el período comprendido entre los años 1998 y 2005 se presentó el mayor incremento en la BA de los árboles sobrevivientes (4,42 t·ha-1·año-1) en la sub-parcela E, en comparación con los incrementos del período entre los años 2005 y 2008 de 1,69 t·ha-1·año-1 y 3,18 t·ha-1·año-1 en las sub-parcelas E y U respectivamente. Por otro lado, la TINBA fue positiva en el período entre los años 1998 y 2005 (con 2,62 t·ha-1·año), y en el período entre los años 2005 y 2008 fue negativa en ambas sub-parcelas (con -14,86 t·ha-1·año y -5,64 t·ha-1·año) (Tabla 10). Estos valores negativos se debieron a la alta producción de necromasa a causa de la intervención antrópica (extracción selectiva de madera). Aparte de esto, el reclutamiento en términos de BA fue evidente solo en el período entre 1998 y 2005. En consecuencia, el bosque incrementó significativamente su BA neta solamente en el período entre 1998 y 2005 (Tabla 10).
Tabla 10. Dinámica de la BA en el bosque pluvial tropical de Salero (Chocó, Colombia).
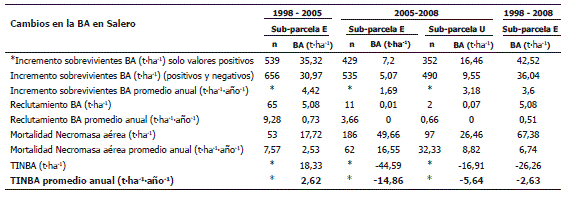
PPNA y precipitación promedia anual en bosques tropicales. En el bosque pluvial tropical de Salero, se registró una PPNA promedio (incremento en BA de sobrevivientes + producción de hojarasca) de carbono 5,21 t·ha-1·año-1. Los valores de PPNA de los rangos de precipitación (de 500 - 2.000; 2.000 - 4.000; 4.000 - 6.000; 6.000 - 8.000 y 8.000 - 10.000 mm anuales) presentaron un promedio general de carbono de 5,25 t·ha-1·año-1; así mismo, en el rango de precipitación de entre 500-2.000 mm anuales se registró la mayor PPNA con un promedio de carbono de 5,88 t·ha-1·año-1, mientras que en el rango de precipitación de entre 4.000 -6.000 mm anuales se observó la menor PPNA con un promedio de carbono de 2,43 t·ha-1·año-1 (Figura 5).
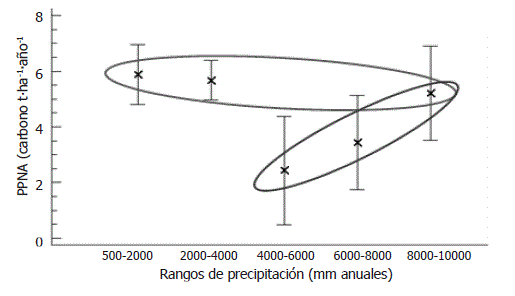
Figura 5. Análisis de varianza de la productividad primaria neta aérea con relación a los rangos de precipitación promedia anual en bosques tropicales. Las X son los promedios de PPNA y las líneas horizontales son los intervalos de confianza de los datos. Los valores dentro de los circulos azules y rojos muestran los grupos formados mediante la prueba de Bonferroni. Datos tomados de Clark et al. (2001) e incluidos los valores de Salero (Chocó, Colombia) y Bajo Calima (Valle del Cauca, Colombia).
Mediante el estadístico de Kruskal-Wallis se detectaron diferencias significativas entre las medianas de la PPNA (estadístico = 12,99; P = 0,011) de los rangos de precipitación evaluados; y mediante el estadístico de rangos múltiples de Bonferroni se identificaron dos grupos de rangos de precipitación con relación a la PPNA, el primer grupo estuvo formado por los rangos de 500-2.000, 2.000-4.000 y 8.000-10.000 mm anuales, mientras que el segundo estuvo conformado por los rangos 4.000-6.000, 6.000-8.000 y 8.000-10.000 mm anuales (Figura 5). El hecho de que el rango de entre 8.000-10.000 mm anuales (correspondiente a Salero) hiciera parte de ambos grupos estadísticos, mostró la poca influencia de la precipitación sobre la PPNA.
DISCUSIÓN
Exactitud de la estimación de la biomasa aérea con los modelos utilizados. Los modelos 1 y 3 empleados para estimar la BA de árboles en Salero (Chocó), generaron valores estadísticamente diferentes a los demás modelos (Tabla 2), lo cual también ha sido reportado por diversos investigadores, quienes han documentado que distintas ecuaciones generan estimaciones diferentes de BA (Araujo et al., 1999; Clark et al., 2001; Keller et al., 2001; Sarmiento et al., 2005). Se ha sugerido que los modelos que solo emplean una variable como el DAP en la estimación de la BA, podrían dar origen a mayores errores, pues se ignoran las variaciones en arquitectura y densidad de la madera de las especies (Clark et al., 2001; Chave et al., 2003), especialmente en bosques con alta diversidad de especies como los del Chocó (Faber y Gentry, 1991). Sin embargo, en el presente estudio se observó que cuando se analizan los valores de BA a nivel de bosque (BA total), las estimaciones tienden a ser muy similares, debido probablemente a que los valores se compensan entre sí (Tabla 3), por lo cual el margen de error es técnicamente aceptable, a pesar de que el margen de error de los inventarios estén alrededor del 20%; así ha sido documentado también en bosques de la Amazonia brasilera por investigadores como Baker et al. (2004a). Por otro lado, las variaciones en la BA al nivel de los cuadrantes de las sub-parcelas E y U, están parcialmente explicadas por las variaciones en las características edáficas de la zona, que han sido analizadas con mayor detalle en otros estudios, como Quinto y Moreno (2010) y Quinto y Moreno (sin publicar).
Comparación de la BA de Salero (Chocó) con otras estimaciones en bosques tropicales. La BA en el bosque pluvial tropical de Salero tuvo un rango de entre 179,2 y 259,9 t·ha-1, con un promedio general de 216,8 t·ha-1. Dicha BA fue ligeramente superior a los informes de BA realizadas por Mosquera y Sánchez (2005), Faber y Gentry (1991) y Rodríguez (1989) de 176 t·ha-1, 169-193 t·ha-1 y 190 t·ha-1 respectivamente, realizadas con anterioridad en distintos bosques del Chocó Biogeográfico. En general, los valores de BA encontrados en Salero (Chocó) de 216,8 t·ha-1, se encuentran en la porción inferior del rango de BA de árboles (entre 167 y 419 t·ha-1) referido por Sarmiento et al. (2005) para selvas neotropicales de baja altitud. Estos resultados de BA de Salero (Chocó) también se encuentran dentro del rango (entre 45-649 t·ha-1, con 275,16 t·ha-1 promedio) expresado por Clark et al. (2001b) para bosques tropicales de distintas zonas del mundo.
Incremento de la BA del bosque pluvial tropical de Salero, Chocó. La BA del bosque pluvial no cambió significativamente en el tiempo (Tabla 5), debido a esto, los resultados en términos de la BA total, no apoyan la hipótesis del incremento en BA en bosques tropicales planteada por Phillips et al. (1998), Malhi et al. (1998, 2004) y Baker et al. (2004b). En el bosque pluvia tropical de Salero, Chocó, se encontró un incremento promedio anual de BA de árboles sobrevivientes de entre 1,69 y 4,42 t·ha-1·año-1 (Tabla 10), el cual está dentro del rango de incremento en BA de árboles reportado por Clark et al. (2001b) de entre 0,6 a 7,6 t·ha-1·año-1, para diversos bosques tropicales. Por otro lado, solo la TINBA de la sub-parcela "E" entre 1998 y 2005, presentó un promedio de 2,61 t·ha-1·año-1 (Tabla 10), que no incluyó el cero en sus intervalos de confianza (Tabla 11). Por tal razón, el bosque presentó un incremento positivo de BA, sólo en dicho período, superior a los incrementos reportados por Chave et al. (2008) para diferentes bosques tropicales (Tabla 11). Este hecho (TINBA de 2,61 t·ha-1·año-1), está acorde con lo planteado por la hipótesis del incremento en la BA en los bosques húmedos tropicales registrado en las ultimas décadas (Phillips et al., 1998; Malhi et al., 1998, 2004; Baker et al., 2004b).
Tabla 11. Valores de BA, incremento, reclutamiento, mortalidad y cambio neto de la biomasa aérea de bosques primarios tropicales incluidos los datos de Salero (Chocó, Colombia), modificado de Chave et al. 2008. BA es biomasa aérea (t·ha-1). Los datos son de la sub-parcela "E" (años 1998-2005).
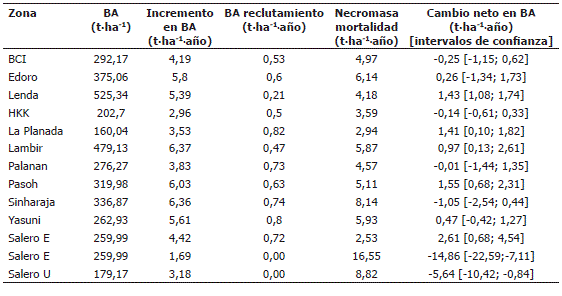
Al analizar las variaciones a través del tiempo de la BA en términos de la TINBA fue posible detectar los cambios en las capturas y emisiones de carbono de los bosques, en comparación con los análisis en términos de la BA total, debido a que los cambios en la BA total a través del tiempo no apoyaron la hipótesis del incremento en la captura de carbono de los bosques tropicales (Phillips et al., 1998, Baker et al., 2004b). Así mismo, los valores negativos en las TINBA del periodo entre 2005 y 2008 (Tabla 11) fueron producto de las deforestaciones; frente a lo cual, solo se puede aseverar que en la parcela debido a la intervención se incrementó la emisión de carbono a la atmosfera, la fragmentación y la masa de detritos, como se ha documentado en otros bosques tropicales (Laurance et al., 1998; Gerwing, 2002; Broadbent et al., 2008).
PPNA y precipitación promedia anual en bosques tropicales. Recientemente, Schuur (2003) manifestó que en los bosques tropicales con precipitación promedia anual (PPA) superior a 5.000 mm, la PPNA tiende a disminuir debido a la reducción de la tasa de descomposición y de la disponibilidad de nutrientes; además, por el incremento en la lixiviación y la nubosidad que merman la PPN. Sin embargo, en este estudio, mediante estadístico de rangos múltiples no se encontraron diferencias significativas entre los promedios de PPNA de los bosques tropicales con precipitaciones entre 500-4.000 y de 8.000-10.000 mm anuales (Figura 5); con lo cual, los resultados obtenidos, contradicen la hipótesis de Schuur (2003), ya que no fue evidente una disminución en la PPNA con el incremento en la precipitación (Figura 5).
El hecho de que la PPNA registrada fuese alta, a pesar de alta precipitación (contrario a Schuur (2003), se debió posiblemente a las siguientes razones: 1) No hay limitación de oxígeno para la descomposición en el suelo por exceso de agua, debido a que en estos ecosistemas a pesar de las altas precipitaciones los suelos tienen buen drenaje. 2) las tasas de descomposición de hojarasca de las zonas en las que se presentan anegamientos temporales, posiblemente son similares a las de las áreas bien drenadas. Puesto que en los suelos en los que el nivel freático oscila unos pocos centímetros con las inundaciones, la hojarasca se encuentra sometida a períodos alternativos de anegamiento y aireación, dependiendo de las lluvias, esto produce pulsos de oxidación de la hojarasca haciendo que se descomponga, en las áreas inundadas, a tasas similares a las registradas en suelos sin inundación (Del Valle, 2003). 3) Los niveles de precipitación en el bosque pluvial tropical cercano a Salero (Chocó), varían significativamente a lo largo del día. A tal punto que se presentan lluvias con mayor frecuencia e intensidad en las horas de la noche (Murillo et al. 2005). Esto implica que, en las horas de mayor radiación solar, entre las 10:00 y las 15:00 horas, la fotosíntesis no es inhibida por la precipitación, y por ende la PPN puede registrar valores altos.
Para Baker et al. (2003), en bosques tropicales no estacionales, en los que la disponibilidad de agua no es una limitante, la radiación solar es la variable ambiental más influyente sobre las tasas de crecimiento. De manera similar, en el bosque pluvial tropical de Salero (Chocó), a pesar de presentar una alta PPA, las lluvias no constituyeron una limitante para la PPN, debido probablemente a que en la zona se registra una alta radiación solar, los suelos son bien drenados, las tasas de descomposición de la hojarasca son altas, como se han documentado en bosques del municipio de Lloró (Chocó) muy cercanos al área de estudio (Quinto y Ramos, 2009), y hay una buena disponibilidad de nutrientes para las plantas (Quinto y Moreno, 2010). Por otro lado, cuando se supera el margen de los 10.000 mm en PPA en bosques tropicales, el panorama para la PPN es aún incierto, pues no ha sido muestreado.
En síntesis, los valores de PPNA obtenidos en este estudio, contradicen la hipótesis de la disminución de la PPNA de los bosques tropicales cuando la precipitación es muy alta, propuesta por Schuur (2003). Además, se evidenció que el bosque en condiciones imperturbadas captó de forma significativa carbono atmosférico, acorde con la hipótesis del incremento en BA de los bosques tropicales de Phillips et al., 1998; Malhi et al., 1998, 2004; Baker et al., 2004a. Por último, queda probado el efecto negativo de las intervenciones antrópicas, cuando son realizadas sin planes de manejo sostenible del recurso, sobre las existencias y tasas de incremento neto de la BA, porque cuando se deforesta masivamente los bosques las emisiones de carbono superan las tasas de captura, de ahí la importancia de la conservación y el manejo sostenible de estos complejos ecosistemas, especialmente en regiones tropicales como el Chocó Biogeográfico.
AGRADECIMIENTOS
Este trabajo contó con el apoyo del proyecto "CRECIMIENTO Y PRODUCTIVIDAD DEL BOSQUE EN UN GRADIENTE ALTITUDINAL: Efectos Potenciales en el Funcionamiento del Sistema de Transmisión de Energía de Colombia", cofinanciado por la Empresa de Interconexión Eléctrica S. A. (ISA) y la Universidad Nacional de Colombia, Sede Medellín, mediante el programa de becas del Fondo de Investigaciones I.S.A. - Universidad Nacional.
BIBLIOGRAFÍA
Angelsen, A., S. Brown, C. Loisel, L. Peskett, Ch. Streck y D. Zarin. 2009. Reducing emissions from deforestation and forest degradation (REDD): an options assessment report. Prepared for the government of Norway.Meridian Institute, http://www.REDD-OAR.org. 100 p.; consulta: febrero 2009.
Araujo, T.M., N. Higuchiand and J.A. Carvalho. 1999. Comparison of formulae for biomass content determination in a tropical rain forest site in the state of Para, Brazil. Forest Ecology and Management 117(1-3): 43-52.
Asprilla, A., C. Mosquera, H. Valoyes, H. Cuesta y F. García. 2003. Composición florística de un bosque pluvial tropical (bp-T) en la parcela permanente de investigación en biodiversidad (PPIB) en Salero, Unión Panamericana, Chocó. pp. 39-44. En: García, F., Y. Ramos, J. Palacios, J.E. Arroyo, A. Mena y M. González (eds.). Salero diversidad biológica de un bosque pluvial tropical (bp-T). Editora Guadalupe Ltda., Bogotá. 207 p.
Baker, T.R., M.D. Swaine, y D.F. Burslem. 2003. Variation in tropical forest growth rates: combined effects of functional group composition and resource availability. Perspectives in Plant Ecology, Evolution and Systematics 6(1-2): 21-36.
Baker, T.R., O.L. Phillips, Y. Malhi, S. Almeida, L. Arroyo, A. Di Fiore, T. Erwin, T.J. Killeen, S.G. Laurance, W.F. Laurance, S.L. Lewis, J. Lloyd, A. Monteagudo, D.A. Neill, S. Patiño, N.A. Pitman, J.N. Silva and R. Vásquez. 2004a. Variation in wood density determines spatial patterns in Amazonian forest biomass. Global Change Biology 10: 545-562.
Baker, T. R., O.L. Phillips, Y. Malhi, S. Almeida, L. Arroyo, A. Di Fiore, T Erwin, N Higuchi, T. Killeen, S.G. Laurance, W.F. Laurance, S.L. Lewis, A. Monteagudo, D.A. Neill, P.N Vargas, N.C.A. Pitman, N. Silva y R. Vásquez. 2004b. Increasing biomass in Amazonian forest plots. Philosophical Transactions of the Royal Society. 359:353-365.
Beers, T.W. 1962. Components of forest growth. Journal of Forestry 60: 245-248.
Bormann, F.H. and G.E. Likens. 1979. Catastrophic disturbance and the steady state in northern hardwood forests. American Scientist 67(6): 660-669.
Broadbent, E.N., G. Asner, M. Keller, D. Knapp, P.J. Oliveira and J.N. Silva. 2008. Forest fragmentation and edge effects from deforestation and selective logging in the Brazilian Amazon. Biological Conservation 141(7): 1745-1757.
Brown, S. and A.E. Lugo. 1982. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 14: 161-187.
Brown, S. 1997. Estimating biomass and biomass change of tropical forests: A primer. Food and Agriculture Organization (FAO), Rome. 57 p. (Forestry Paper No. 134).
Chambers, J.Q., J. Dos Santos, R.J. Ribeiro and N. Higuchi. 2001. Tree damage, allometric relationships, and above-ground net primary production in central Amazon forest. Forest Ecology and Management 152: 73-84.
Chave, J., R. Condit, S. Lao, J.P. Caspersen, R.B. Foster and S.P. Hubbell. 2003. Spatial and temporal variation of biomass in a tropical forest: results from a large census in Panamá. Journal of Ecology 91(2): 240-252.
Chave, J., C. Andalo, S. Brown, M. Cairns, J. Chambers, D. Eamus, H. Folster, F. Fromard, N. Higuchi, T. Kira, J. Lescure, B. Nelson, H. Ogawa, H. Puig, B. Riera and T. Yamakura. 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145: 87-99.
Chave, J., R. Condit, H. Muller, S. Thomas, P. Ashton, S. Bunyavejchewin, L. Co, H. Dattaraja, S. Davies, S. Esufali, C. Ewango, K. Feeley, R. Foster, N. Gunatilleke, S. Gunatilleke, P. Hall, T. Hart, C. Hernández, S. Hubbell, A. Itoh, S. Kiratiprayoon, J. La Frankie, S. Loo de Lao, J. Makana, M. Noor, A. Kassim, C. Samper, R. Sukumar, H. Suresh, S. Tan, J. Thompson, M. Tongco, R. Valencia, M. Vallejo, G. Villa, T. Yamakura, J. Zimmerman and E. Losos. 2008. Assessing evidence for a pervasive alteration in tropical tree communities. Plos Biology 6(3): 1-19.
Chazdon, R. y R. Montgomery. 2002. La adquisición de carbono en las plantas. pp. 167-192. En: Guariguata M. y G. Kattan (eds). Ecología y conservación de bosques neotropicales. Libro Universitario Regional, Cartago, Costa Rica. 691 p.
Clark, D.A. 2002. Are tropical forests an important carbon sink? Reanalysis of the long-term plot data. Ecological Applications 12(1): 3-7.
Clark, D.A. y D.B. Clark. 1994. Climate-induced annual variation in canopy tree growth in a Costa Rican tropical rain forest. Journal of Ecology 82(4): 865-872.
Clark, D.B. and D.A. Clark. 2000. Landscape-scale variation in forest structure and biomass in a tropical rain forest. Forest Ecology and Management 137: 185-198.
Clark, D.A., S. Brown, D. Kicklighter, J. Chambers, J. Thomlinson and J. Ni. 2001a. Measuring net primary production in forest: concepts and field methods. Ecological Aplications 11(2): 356-370.
Clark, D.A., S. Brown, D. Kicklighter, J. Chambers, J. Thomlinson, E. Holland and J. Ni. 2001b. Net primary production in forest: an evaluation and sinthesis of existing field data. Ecological Aplications 11(2): 356- 370.
Corporación Autónoma Regional del Centro de Antioquia (CORANTIOQUIA). 2006. Diversidad y productividad de los bosques de Antioquia. Proyecto "Manejo y conservación de la flora". Informe final. Primera fase. CORANTIOQUIA - Universidad Nacional de Colombia, Medellín. 87 p.
Córdoba, J.A., D. González, Y. Ramos, D. Serna, y N. Panesso. 2005. Regeneración natural en claros de un bosque pluvial tropical en Pacurita, Chocó - Colombia. Revista Institucional. Universidad Tecnológica del Chocó D.L.C. 23: 11-19.
Cuartas, C. 1993. Estimadores cuantitativos de la biomasa aérea de especies dominantes en un bosque inundable de la Amazonia colombiana. Trabajo de grado de Ingeniería Forestal. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 124 p.
Del Valle, J.I. 2003. Descomposición de la hojarasca fina en bosques pantanosos del Pacifico colombiano. Interciencia 28(3):148- 153.
Del Valle, J.I., F. Moreno y S. Orrego. 2003. Los bosques tropicales y su contribución a la mitigación del cambio climático. pp. 3-43. En: Orrego S.A., J.I. Del Valle y F.H. Moreno (eds.). Medición de la captura de C en ecosistemas forestales tropicales de Colombia. Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia, Medellín. 314 p.
Dixon, R.K., S. Brown, R. Houghton, A. Solomon, M. Trexler and J. Wisniewski. 1994. Carbon pools and flux of global forest ecosystems. Science 263: 185-190.
Faber, D. and A.H. Gentry. 1991. The structure and diversity of rain forests at bajo Calima, Chocó region, western Colombia. Biotropica 23(1): 2-11.
Feeley, K.J., S. Wright, M. Supardi, A. Kassim and S. Davies. 2007. Decelerating growth in tropical forest trees. Ecology Letters 10: 461-469.
Forero, E. y A. Gentry. 1989. Lista anotada de plantas del departamento del Chocó, Colombia. Instituto de Ciencias Naturales - Universidad Nacional de Colombia. Museo de Historia Natural. Editorial Guadalupe Ltda., Bogotá. 142 p.
García, F., Y. Ramos, J. Palacios, J. Arroyo, E. Mena y M. González. 2003. Salero diversidad biológica de un bosque pluvial tropical (bp-T). Editorial Guadalupe Ltda., Bogotá, Colombia. 207 p.
Gentry, A. 1993. A field guide to the families and genera of woody plants of Northwest South America (Colombia, Ecuador, Perú). University of Chicago. Chicago. 895 p.
Gerwing, J. 2002. Degradation of forests through logging and fire in the eastern Brazilian Amazon. Forest Ecology and Management 157: 131-141.
Guttman, I. 1982. Linear models: an introduction. First edition. Wiley, John and Sons Incorporated, USA. 358 p.
Hogan, K.P. y J.L. Machado. 2002. La luz solar: consecuencias biológicas y medición. pp. 119-143. En: Guariguata, M. y G. Kattan (eds). Ecología y conservación de bosques neotropicales. Libro Universitario Regional. LUR. Costa Rica. 691 p.
Holdridge, L.P. 1982. Ecología basada en las zonas de vida. Instituto Interamericano para la Agricultura. San José, Costa Rica. 216 p.
Hoshmand, A.R. 1998. Statistical methods for environmental and agricultural sciences. Second edition. CRC Press LLC, New York, USA. 439 p.
Houghton, R.A. 2005. Aboveground forest biomass and the global carbon balance. Global Change Biology 11:945-958.
Intergovernmental Panel on Climate Change (IPCC). 2001. Summary for policymakers. A report of working group I of the Intergovernmental Panel on Climate Change. Wembley, United Kingdom. 34 p.
Keller, M., M. Palace and G. Hurtt. 2001. Biomass estimation in the Tapajos National Forest, Brazil. Examination of sampling and allometric uncertainties. Forest Ecology and Management 154(3): 371-382.
Laurance, W., L.V. Ferreira, J.M. Rankinand S.G. Laurance. 1998. Rain forest fragmentation and the dynamics of amazonian tree communities. Ecology 79(6): 2032-2040.
Luyssaert, I., M. Inglima, A. Jung, D. Richardson, M. Reichstein, D. Papale, S.L. Piao, E.D. Schulze, L. Wingate, G. Matteucci, L. Aragao, M. Aubinet, C. Beer, C. Bernhofer, K. Black, D. Bonal, J. Bonnefond, J. Chambers, P. Ciais, B. Cook, K. Davis, A.J. Dolman, B. GiElen, M. Goulden, J. Grace, A. Granier, A. Grelle, T. Griffis, T. Grunwald, G. Guidolotti, P.J. Hanson, R. Harding, D.Y. Hollinger, L.R. Hutyra, P. Kolari, B. Kruijt, W. Kutsch, F. Lagergren, T. Laurila, B.E. Law, G. Le Maire, A. Lindroth, D. Loustau, Y. Malhi, J. Mateus, M. Migliavacca, L. Misson, L. Montagnani, J. Moncrieff, E. Moors, J. Munger, E. Nikinmaa, S. Ollinger, G. Pita, C. Rebmann, O. Roupsard, N. Saigusa, M. Sanz, G. Seufert, C. Sierra, M. Smith, J. Tang, R. Valentini, T. Vesala and I. Janssens. 2007. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Global Change Biology 13: 2509-2537.
Mahecha, G. 1997. Fundamentos y metodologías para la identificación de plantas. Proyecto Biopacífico, Ministerio del Medio Ambiente, GEF - PNUD. Lerner Ltda., Bogotá. 282 p.
Malhi, Y., A. Nobre, J. Grace, B. Kruijt, M. Pereira, A. Culf, and S. Scott. 1998. Carbon dioxide transfer over a Central Amazonian rain forest. Journal of Geophysical Research 103: 31593-31612.
Malhi, Y., T. Baker, O. Phillips, S. Almeida, E. Alvarez, L. Arroyo, J. Chave, C. Czimczik, A. Difiore, N. Higuchi, T. Killeen, S. Laurance, W. Laurance, S. Lewis, L. Mercado, A. Monteagudo, D. Neill, P. Nuñez, S. Patiño, N. Pitman, C. Quesada, R. Salomao, J. Silva, A. Lezama, R. Martínez, J. Terborgh, B. Vinceti and J. Lloyd. 2004. The above-ground coarse wood productivity of 104 neotropical forest plots. Global Change Biology 10: 563-591.
Melo, O. y R. Vargas. 2003. Evaluación ecológica y silvicultural de ecosistemas boscosos. Universidad del Tolima, Impresiones CONDE, Ibagué. 235 p.
Mosquera, Y. y C. Sánchez. 2005. Propuesta para la venta de servicios ambientales con participación comunitaria en el Medio Atrato. Chocó - Colombia. Trabajo de grado. Facultad de Ingeniería. Programa de Ingeniería Ambiental. Universidad Tecnológica del Chocó "Diego Luís Córdoba". Quibdó, Chocó, Colombia. 135 p.
Mostacedo, B. y T. Fredericksen. 2000. Manual de Métodos Básicos de Muestreo y Análisis en Ecología Vegetal. Proyecto Bolfor. Editora El País, Santa Cruz, Bolivia. 82 p.
Murillo, W., R. Palomino, S. Córdoba, G. Aragón y E. Banguero. 2005. El régimen diario de la precipitación en el municipio de Quibdó (Colombia). Revista de Climatología 5: 1-7.
Odum, E. 1969. The strategy of ecosystem development. An understanding of ecological succession provides a basis for resolving mans conflict with nature. Science, New Series 164(3877): 262-270.
Orrego, S.A. y J.I. Del Valle. 2003. Existencias y tasas de incremento neto de la biomasa y del C en bosques primarios intervenidos y secundarios. pp. 215-242. En: Orrego, S.A., J.I. Del Valle y F.H. Moreno (eds.). Medición de la captura de C en ecosistemas forestales de Colombia: Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia, Sede Medellín, Departamento de Ciencias Forestales y Centro Andino para la Economía en el Medio Ambiente, Bogotá. 314 p.
Orrego, S.A., J.I. Del Valle y F.H. Moreno. 2003. Medición de la captura de C en ecosistemas forestales de Colombia: Contribuciones para la mitigación del cambio climático. Universidad Nacional de Colombia, Sede Medellín, Departamento de Ciencias Forestales y Centro Andino para la Economía en el Medio Ambiente, Bogotá. 314 p.
Palacios, J., Y. Ramos, A. Mosquera, F.L. Castro, F. García, J. Arroyo y A. Cogollo. 2003b. Estructura de un bosque pluvial tropical (bp-T) en Salero, Unión Panamericana, Chocó. pp. 45-61. En: García, F., Y. Ramos, J. Palacios, J.E. Arroyo, A. Mena y M. González. Salero diversidad biológica de un bosque pluvial tropical (bp-T). Editorial Guadalupe Ltda, Bogotá. 207 p.
Phillips, O.L., Y. Malhi, N. Higuchi, W. Laurance, P. Núñez, M. Vásquez, S. Laurance, L. Ferreira, M. Stern, S. Brown and J. Grace. 1998. Changes in the carbon balance of tropical forests: Evidence from long-term plots. Science 282: 439-442.
Quinto, H., Y. Ramos y D. Abadía. 2007. Cuantificación de la caída de hojarasca como medida de la productividad primaria neta en un bosque pluvial tropical en Salero, Unión Panamericana, Chocó, Colombia. Revista Institucional Universidad Tecnológica del Chocó D.L.C. 26: 28-41.
Quinto, H. y Y. Ramos. 2009. Descomposición de hojarasca en un bosque pluvial tropical intervenido en el municipio de Lloró, Chocó, Colombia. Revista Institucional Universidad Tecnológica del Chocó D.L.C. 28: 32-43.
Quinto, H. y F.H. Moreno. 2010. Dinámica de la biomasa aérea en bosques primarios de Colombia y su relación con la precipitación y la altitud. Tesis de Posgrado en Bosques y Conservación Ambiental. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 75 p.
Rodríguez, J.C. 1989. Consideraciones sobre la biomasa, composición química y dinámica del bosque pluvial tropical de colinas bajas. Bajo Calima. Buenaventura, Colombia. Corporación Nacional de Investigación y Fomento Forestal - CONIF, Bogotá, D.E. 36 p. (Serie Documental No. 16).
Sarmiento, G., M. Pinillos and I. Garay. 2005. Biomass variability in tropical american lowland rainforests. Ecotropicos 18(1): 1-20.
Schuur, E.A.G. 2003. Productivity and global climate revisited: the sensitivity of tropical forest growth to precipitation. Ecology 84(5): 1165-1170.
Sierra, C.A., J.I. Del Valle, S.A. Orrego, F.H. Moreno, M.E. Harmon, M. Zapata, G.J.Colorado, M.A. Herrera, W. Lara, D.E. Restrepo, L.M. Berrouet, L.M. Loaiza, and J.F. Benjumea. 2007. Total carbon stocks in a tropical forest landscape of the Porce region, Colombia. Forest Ecology and Management 243(2-3): 299-309.
Statistical Graphics Corporation. 2002. Statgraphics Plus Version 5.1, www.statgraphics.com.
Swaine M.D. and J.B. Hall. 1983. Early succession on cleared forest land in Ghana. Journal of Ecology 71(2): 601-627.
Turner, I.M. 2001. The ecology of trees in the tropical rain forest. First publisher. Cambridge University Press, Cambridge. 298 p.
Universidad Nacional de Colombia, Sede Medellín. (UNALMED). 2001. Evaluación de las plantaciones de Acacia mangium Willd en áreas degradadas del Bajo Cauca. Informe Técnico. Universidad Nacional de Colombia - CORANTIOQUIA, Medellín. 325 p.
Vallejo, M., A.C. Londoño, R. López, G. Galeano, E. Álvarez, y W. Dévia. 2005. Establecimiento de parcelas permanentes en bosques de Colombia (métodos para estudios ecológicos largo plazo). Instituto de Investigaciones de Recursos Biológicos Alexander von Humboldt, Bogotá D.C. 310 p.
Whitmore, T.C. 1975. Tropical rain forest of the far east. Oxford University Press. Scotland. Reino Unido. 260 p.
Whitmore, T.C. 1998 Anintroduction to tropical rain forests. Second edition. Oxford University Press, Oxford. 226 p.
Yepes, A. 2008. Dinámica de un bosque primario premontano de los Andes Colombianos. Tesis de Posgrado en Bosques y Conservación Ambiental. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 75 p.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
Article abstract page views
Downloads
License
Copyright (c) 2015 Revista Facultad Nacional de Agronomía

This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
The journal allows the author(s) to maintain the exploitation rights (copyright) of their articles without restrictions. The author(s) accept the distribution of their articles on the web and in paper support (25 copies per issue) under open access at local, regional, and international levels. The full paper will be included and disseminated through the Portal of Journals and Institutional Repository of the Universidad Nacional de Colombia, and in all the specialized databases that the journal considers pertinent for its indexation, to provide visibility and positioning to the article. All articles must comply with Colombian and international legislation, related to copyright.
Author Commitments
The author(s) undertake to assign the rights of printing and reprinting of the material published to the journal Revista Facultad Nacional de Agronomía Medellín. Any quotation of the articles published in the journal should be made given the respective credits to the journal and its content. In case content duplication of the journal or its partial or total publication in another language, there must be written permission of the Director.
Content Responsibility
The Faculty of Agricultural Sciences and the journal are not necessarily responsible or in solidarity with the concepts issued in the published articles, whose responsibility will be entirely the author or the authors.


















