
Notas sobre el género de abejas augochlorinas Chlerogas (Hymenoptera:
Halictidae)
MICHAEL S. ENGEL
Division of Entomology, Natural History Museum, and Department of Ecology
& Evolutionary Biology, 1501 Crestline Drive - Suite 140, University of
Kansas, Lawrence, Kansas 66049-2811, USA. msengel@ku.edu
ABSTRACT
Supplemental information to the earlier monograph of the rare bee genus Chlerogas Vachal (Halictinae: Augochlorini) is provided. The recently identified males of C. boliviensis Brooks & Engel and C. cyaneus Brooks & Engel are described and figured, including the first account of the eversible endophallic structure for the genus. In addition, C. cyaneus, previously known only from northern Ecuador, is newly recorded from north-central Colombia.
Key words. Andes, Anthophila, Apoidea, Halictinae, Bolivia, Colombia, taxonomy.
RESUMEN
Se presenta información adicional a la monografía del raro género de abejas Chlerogas Vachal (Halictinae: Augochlorini). Se describen por primera vez los machos de C. boliviensis Brook & Engel y C. cyaneus Brooks & Engel, incluyendo la primera descripción de la estructura endofálica reversible para el género. Además, C. cyaneus, una especie anteriormente conocida del norte de Ecuador, es registrada por primera vez para el norte-centro de Colombia.
Palabras clave. Andes, Anthophila, Apoidea, Halictinae, Bolivia, Colombia, taxonomía.
Recibido: 18/06/2009INTRODUCTION
The Neotropical bee genus Chlerogas Vachal is one of several bee genera in the tribe Augochlorini to possess a characteristically elongate head, in this case resulting from an elongate malar space, clypeus, and supraclypeal region (Figs. 1-3).
Unlike the other augochlorine genera with such elongate heads; namely Chlerogella Michener, Ischnomelissa Engel, Chlerogelloides Engel et al., and to a limited extent Rhynchochlora Engel; Chlerogas comprises consistently larger species with a reduced number of flagellomeres (nine in females, 10 in males), pectinate inner metatibial spur in males, and large hammer-like volsellae (Engel, 2000). In some respects Chlerogas is reminiscent of the similarly-sized and similarly-longheaded Old World genus Thrinchostoma Sausurre, although the latter is a member of the tribe Halictini (including Thrinchostomatini, sensu Engel, 2005; Michener, 2007). While Chlerogella, Ischnomelissa, and Chlerogelloides are all clearly related as evidenced by their pronotal, propodeal, and genitalic structure (Engel, 2000), Chlerogas and Rhynchochlora are not related to the aforementioned group (Engel, 2000, 2007). Unfortunately, although these genera can be readily identified, specimens of each are scarce and the species apparently of relatively low abundance. To date no real details of their biology have been documented outside of a few, largely anecdotal, floral records. In order to bring more attention to the diversity of these distinctive bees, studies to revise these genera have been on-going since the mid-1990s. Chlerogas, presently with 11 species (Table 1), was revised by Brooks & Engel (1999), with brief supplements provided by Engel et al. (2006) and Engel & Gonzalez (2009), while the other genera were synthesized by Engel et al. (1997), Engel & Brooks (1999, 2002), Brooks & Engel (1998), and Engel (2003a, 2003b, 2007). The final and most diverse long-headed genus to be revised is Chlerogella, a work now completed after more than a decade of investigation (Engel, in rev.).
Herein I provide supplemental notes on two species of Chlerogas, one newly recorded from Colombia and the first description of the male for this same species as well as for another from Bolivia. Morphological terminology follows that of Engel (2000, 2001), while the format for the description is generally based on that of Brooks & Engel (1999).
Chlerogas boliviensis Brooks & Engel
Figs. 1-3, 4-7
Chlerogas boliviensis Brooks & Engel, 1999: 468 [?].
Material. ?: Bolivia, Santa Cruz, Florida Prov., 11 km N.E. Achira, 1800 m, 3.xi.1999 [3 November 1999], C. Porter & L. Stange, cloud forest. Specimen deposited in the Division of Entomology, University of Kansas Natural History Museum, Lawrence, Kansas, USA.
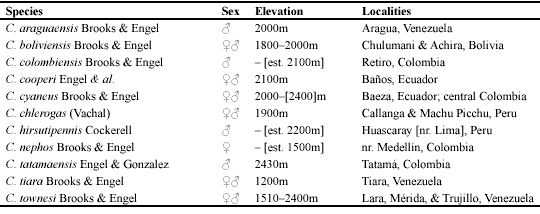
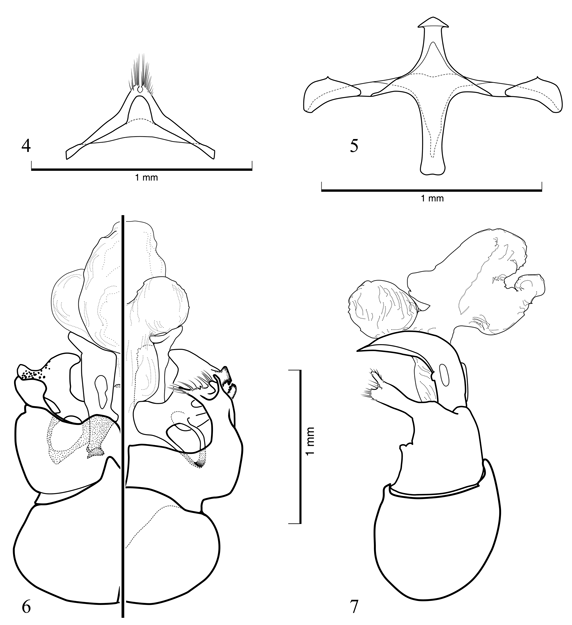
Diagnosis. Among uniformly metallic or dark metallic Chlerogas, the female of C. boliviensis is the only species with a non-striate basal area to the propodeum. The male can be readily distinguished from other Chlerogas by its uniformly black coloration, lacking significant metallic highlights, and by the structure of its terminalia (Figs. 4-7).
Description. ?: As described for the female (vide Brooks & Engel, 1999) with the following modifications: Total body length 13 mm; forewing length 8.6 mm. Head 1.56 times longer than wide, narrower than mesosoma. Gena nearly as broad as compound eye in profile. Base of clypeus below lower tangent of compound eyes by 0.27 mm. Head width 2.07 mm; length (to apex of clypeus) 3.23 mm; clypeal length 1.25 mm; malar space length 0.99 mm; lower interorbital distance 0.77 mm; upper interorbital distance 0.93 mm; intertorular distance 0.20 mm; torular-ocular distance 0.20 mm; torular-median ocellar distance 0.65 mm; distance between lateral ocelli 0.23 mm; distance between lateral ocellus and median ocellus 0.08 mm; ocellocular distance 0.22 mm. Scape not reaching to lateral ocellus, length 0.67 mm; pedicel as long as wide; F1 as long as wide, about as long as pedicel, length 0.18 mm; remaining flagellomeres longer than wide; F2 0.27 mm in length; F3 and F4 each 0.40 mm in length; F5-F7 each 0.43 mm in length; F8 and F9 each 0.47 mm in length; F10 0.67 mm in length. Intertegular distance 1.73 mm; mesoscutellum weakly bituberculate (paramedially). Basal area of propodeum shorter than mesoscutellum, slightly longer than metanotum; basal area strongly granular, without striae, although with exceedingly short, weak strigae at extreme base laterally. Forewing with basal vein distad cu-a by 1.5 times vein width; 1rs-m distad 1m-cu by twice vein width; 2rs-m distad 2m-cu by 7 times vein width; marginal cell length 2.3 mm, width 0.50 mm; first submarginal cell about as long as combined lengths of second and third submarginal cells (as measured along their posterior borders); second submarginal cell slightly narrowed anteriorly; anterior border of second submarginal cell along Rs shorter than anterior border of third submarginal cell; distal hamuli arranged 3-1-3. Inner metatibial spur with four teeth, not including apex. Metasomal terga and sterna imbricate, with scattered weak punctures; sternum IV apically with paramedial patches of dense golden setae; sternum V gently concave medially, with distinct, pale gold setae fringing apical borders except in medial concavity; sternum VI deeply concave medially, with dark gold to fuscous setae except medially along inner and proximal border of concavity; hidden sterna and genitalia as in figures 4-7.
Coloration of head (including scape and pedicel) and mesosoma (except as noted) black (Fig. 1); labrum, mouthparts, antennal flagellum, tegula, legs, and metasoma dark brown. Generally lacking metallic highlights except some scattered, exceptionally faint, dark metallic blue highlights on mesosoma and head dorsally; wings weakly infumate.
Comments. The female of C. boliviensis is a large (ca. 11 mm in total length), non-shining, dark metallic blue-green bee (Brooks & Engel, 1999). The male reported herein is of comparable size and agrees in most respects with the description of the female (except in the usual sex differences) but is largely black throughout with only faint highlights of dark metallic blue or blue-green coloration in some places. Such a slight difference in overall coloration between males and females is not uncommon among Augochlorini (Engel, pers. obs.) and thus this trivial difference does not presently contradict the assertion that the male and female are conspecific.
The endophallic structure of C. boliviensis is depicted in figures 6 and 7. This is the first account of this anatomical structure for the genus and, in fact, is one of the few endophallic morphologies known for any Augochlorini (and the only one figured for a member of the tribe). Previously, only four augochlorine species had been studied for endophallic characters: Augochlora (Augochlora) pura (Say), Augochlorella persimilis (Viereck), Augochlorella aurata (Smith) [as A. striata (Provancher)], and Augochloropsis (Paraugochloropsis) metallica (Fabricius) (Roig-Alsina, 1993). All four species possessed eversible sacs; however, while the eversible sac was simple for A. pura and A. metallica, both species of Augochlorella had secondary lobes, albeit not intricately developed. Overall the endophallus was relatively simple as is typical for halictids. The endophallus of C. boliviensis has an eversible sac and, like Augochlorella, possesses secondary lobes (Figs. 6, 7). Unlike Augochlorella, these lobes are rather dramatically developed (Figs. 6, 7) and the two ventral lobes are covered with minute spicules. As more information on the comparative morphology of augochlorine endophalli is amassed it will be interesting to see how such characters alter our understanding of phylogenetic relationships within the tribe, in addition to seeking correlations with particular mating repertoires of these bees.
Chlerogas cyaneus Brooks & Engel
Figs. 8-10
Chlerogas cyaneus Brooks & Engel, 1999: 469 [?].
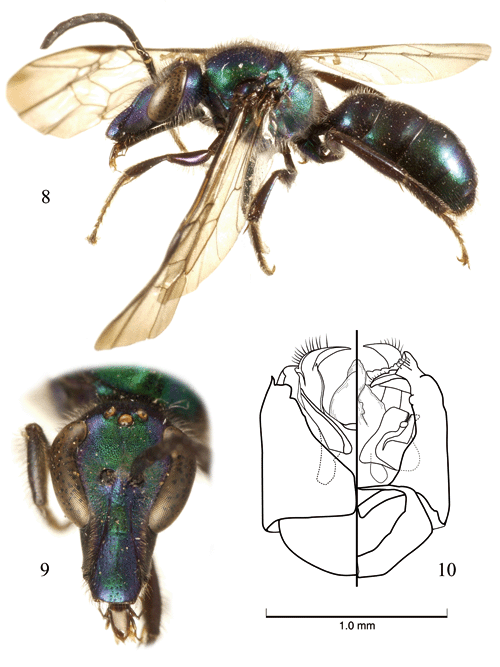
New records. ?: Colombia, Santander Virolin SFF Guanentá, 2400m, malaise [trap], 2.ii.98 [2 February 1998], J. Martinez; deposited in the Instituto Alexander von Humboldt, Bogotá, Colombia. ?: Colombia, Cundinamarca Mundo Nuevo, 1 Oct. [October] 1987, C. Sarmiento; deposited in the Division of Entomology, University of Kansas Natural History Museum, Lawrence, Kansas, USA. ?: Colombia, Cundinamarca, Choachí, 19 Mar. [March] 1988, C. Sarmiento (depicted in figures 8, 9); deposited in the Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá.
Diagnosis. This species most closely resembles C. chlerogas (Vachal), known from Peru, but can be recognized by the completely striate and medially rugulose basal area of the propodeum, the strongly imbricate integument between punctures ventrally on the mesepisternum, and the form of the male terminalia (Fig. 10).
Description. ?: As described for the female (vide Brooks & Engel, 1999) with the following modifications: Total body length 10.2 mm; forewing length 7.85 mm. Head 1.37 times longer than wide (Fig. 9), narrower than mesosoma. Gena nearly as broad as compound eye in profile. Base of clypeus below lower tangent of compound eyes by 0.04 mm. Head width 2.13 mm; length (to apex of clypeus) 2.92 mm; clypeal length 1.1 mm; malar space length 0.58 mm; lower interorbital distance 0.88 mm; upper interorbital distance 1.1 mm; intertorular distance 0.17 mm; torular-ocular distance 0.21 mm; torular-median ocellar distance 0.58 mm; distance between lateral ocelli 0.29 mm; distance between lateral ocellus and median ocellus 0.12 mm; ocellocular distance 0.29 mm. Scape not reaching to lateral ocellus, length 0.63 mm; pedicel as long as wide; F1 as long as wide, about as long as pedicel, length 0.16 mm; remaining flagellomeres longer than wide; F2 0.21 mm in length; F3-F6 each 0.36 mm in length; F7-F9 each 0.42 mm in length; F10 0.52 mm in length. Intertegular distance 1.88 mm; mesoscutellum not bituberculate. Basal area of propodeum about as long as mesoscutellum, about twice as long as metanotum; basal area distinctly striate, striae reaching apical margin, striae becoming slightly shorter medially and somewhat rugulose. Forewing with basal vein distad cu-a by 2 times vein width; 1rs-m confluent with 1m-cu; 2rs-m distad 2m-cu by 6 times vein width; marginal cell length 2.08 mm, width 0.46 mm; first submarginal cell about as long as combined lengths of second and third submarginal cells (as measured along their posterior borders); second submarginal cell slightly narrowed anteriorly; anterior border of second submarginal cell along Rs shorter than anterior border of third submarginal cell; distal hamuli arranged 3-1-3 on left hind wing and 4-1-2 on right hind wing. Inner metatibial spur with five short teeth (almost serrate in one specimen), not including apex. Metasomal terga and sterna imbricate, with scattered minute weak punctures; sternum IV with white setae more dense in narrow zone medially; sternum V with apical margin straight, with narrowly triangular, medioapical patch of lightly fuscouse setae; sternum VI as in C. chlerogas (vide Brooks & Engel, 1999); genitalia as in figure 10.
Coloration of head and mesosoma brilliant metallic green with strong purple and blue highlights, such purple highlights particularly strong in malar area, on clypeus, and bordering compound eyes (Figs. 8-9); such highlights weaker on mesosoma. Labrum and labiomaxillary complex dark brown; mandible dark brown with dark reddish brown apex; antenna black. Tegula dark brown with strong green and purple highlights. Pro- and metacoxae metallic green with blue and purple highlights, remainder of legs dark brown. Metasomal terga dark brown with strong metallic green overlay, particularly on more posterior terga, and with scattered purple highlights; sterna dark brown.
Comments. This species was previously known only from a female found in northern Ecuador (near Baeza: vide Brooks & Engel, 1999). Recently a female conspecific with the holotype and two males (reported herein), collected from north-central Colombia, were brought to my attention. This material expands the range of this species northward into Colombia and permits characterization of the male for the first time. Significant regions of suitable habitat exist between the type locality and the data points in Colombia suggesting numerous localities for potential collection of the species. Based on the material reported herein the species can be added now to the general list of the Colombian bee fauna summarized by Smith-Pardo (2003) and to the listing of those high elevation species summarized by Gonzalez & Engel (2004). Given the general rarity of individuals in the genus the best means for possible sampling of additional Chlerogas in these areas is through the use of traps (malaise and flight intercept traps). It is greatly hoped that exploration throughout central and southern Colombia as well as northern Ecuador may reveal additional material.
ACKNOWLEDGMENTS
I am grateful to J.R. Wiley, Florida State Collection of Arthropods, for sending the male of C. boliviensis to me and permitting its deposition in the University of Kansas Natural History Museum, and to F. Fernandez, Instituto Alexander von Humboldt, and V.H. Gonzalez, Utah State University, for permitting me to examine the specimens of C. cyaneus recovered from Colombia, donating one to the University of Kansas, and for translating my abstract and keywords into Spanish. Two reviewers provided gracious comments on the manuscript and I am thankful for their time and effort. Line illustrations of the terminalia were prepared by S. Taliaferro through support from the University of Kansas Engel Illustration Fund. This project is dedicated to A.H. Smith-Pardo, a new leading researcher in the systematics of Augochlorini. This is a contribution of the Division of Entomology, University of Kansas Natural History Museum.
LITERATURE CITED
1. BROOKS, R.W. & M.S. ENGEL. 1998. New bees of the genus Ischnomelissa
Engel, with a key to the species (Hymenoptera, Halictidae, Augochlorini). Deutsche
Entomologische Zeitschrift 45(2): 181-189.
2. BROOKS, R.W. & M.S. ENGEL. 1999. A revision of the augochlorine bee genus
Chlerogas Vachal (Hymenoptera: Halictidae). Zoological Journal of the Linnean
Society 125(4): 463-486.
3. ENGEL, M.S. 2000. Classification of the bee tribe Augochlorini (Hymenoptera:
Halictidae). Bulletin of the American Museum of Natural History 250: 1-89.
4. ENGEL, M.S. 2001. A monograph of the Baltic amber bees and evolution of the
Apoidea (Hymenoptera). Bulletin of the American Museum of Natural History 259:
1-192.
5. ENGEL, M.S. 2003a. A new species of the bee genus Chlerogella from Panama
(Hymenoptera: Halictidae). Zootaxa 286: 1-4.
6. ENGEL, M.S. 2003b. A new bee of the genus Chlerogella from Ecuador (Hymenoptera,
Halictidae). Pages 135-137 In: Melo, G.A.R. & Alves-dos-Santos, I. (eds.),
Apoidea Neotropica: Homenagem aos 90 Anos de Jesus Santiago Moure. Editora UNESC
[Universidade do Extremo Sul Catarinense], Criciúma.
7. ENGEL, M.S. 2005. Family-group names for bees (Hymenoptera: Apoidea). American
Museum Novitates 3476: 1-33.
8. ENGEL, M.S. 2007. Two new augochlorine bees from Ecuador (Hymenoptera: Halictidae).
Acta Entomologica Slovenica 15(1): 21-29.
9. ENGEL, M.S. & R.W. BROOKS. 1999. A new Chlerogelloides from French Guiana,
with comments on the genus (Hymenoptera: Halictidae). Journal of the Kansas
Entomological Society 72(2): 160-166.
10. ENGEL, M.S. & R.W. BROOKS. 2002. A new bee of the genus Ischnomelissa,
with a key to the known species (Hymenoptera: Halictidae). Entomological News
113(1): 1-5.
11. ENGEL, M.S. & V.H. GONZALEZ. 2009. A new species of Chlerogas from the
Andes of central Colombia (Hymenoptera: Halictidae). Caldasia 31(2): 441-447.
12. ENGEL, M.S., R.W. BROOKS & D. YANEGA. 1997. New genera and subgenera
of augochlorine bees (Hymenoptera: Halictidae). Scientific Papers, Natural History
Museum, University of Kansas 5: 1-21.
13. ENGEL, M.S., F.F. DE OLIVEIRA & A.H. SMITH-PARDO. 2006. A new species
of the bee genus Chlerogas Vachal from Ecuador (Hymenoptera: Halictidae). Entomologist's
Monthly Magazine 142(1703-1705): 103-106.
14. GONZALEZ, V.H. & M.S. ENGEL. 2004. The tropical Andean bee fauna (Insecta:
Hymenoptera: Apoidea), with examples from Colombia. Entomologische Abhandlungen
62(1): 65-75.
15. MICHENER, C.D. 2007. The Bees of the World [2nd Edition]. Johns Hopkins
University Press, Baltimore.
16. ROIG-ALSINA, A. 1993. The evolution of the apoid endophallus, its phylogenetic
implications, and functional significance of the genital capsule (Hymenoptera,
Apoidea). Bollettino di Zoologia 60(2): 169-183.
17. SMITH-PARDO, A.H. 2003. A preliminary account of the bees of Colombia (Hymenoptera:
Apoidea): Present knowledge and future directions. Journal of the Kansas Entomological
Society 76(2): 335-341.