



Publicado
Evidencias de influencia marina durante el Mioceno en el Sudoeste de la Amazonia (Brasil)
Evidence for marine influx during the Miocene in southwestern Amazonia, Brazil
Palabras clave:
Marine Miocene, Brazilian Amazonia, ostracods, Solimões Formation, microfossiles (es)Probables incursiones marinas en la región Amazoníca durante el Mioceno han sido objeto de debates durante mucho tiempo. La base de datos general, impide una exacta localización de las ingresiones marinas propuestas. Sin embargo, la presencia de una mezcla de grupos marinos y de transición o no marinos (foraminíferos, moluscos marinos marginales, dientes de tiburónes, rayas, pólen de los manglares, ostrácodos y percebes) han sido registradas en estudios previos. Esto indica que la influencia marina puede haber jugado, al menos esporádicamente, algún papel en los depósitos neógenos del Amazona, a pesar que las llanuras aluviales fueron los ambientes predominantes (Monsch, 1998; Lundberg et al, 2010;. Hoorn et al, 2010; Hovikoski et al., 2010). Recientes estudios micropaleontológicos con muestras del núcleo (1AS-31-AM, profundidad total 302 m; 05°18'S / 71°02'O; río itui) del suroeste del estado Amazonas corroboran esta evidencia marina. Las muestras entre 170,90 y 174,40 metros de profundidad, contiénen esporádicamente una mezcla de organismos marinos (basados principalmente en la presencia de foraminíferos y briozoarios), otra fauna de transición y también no marinos (principalmente de foraminíferos y ostrácodos). Algunos de estos organismos son resistentes a condiciones de estrés (como el Cyprideis ostrácodos y algunos foraminíferos aglutinados). Aparte de estos elementos de la fauna, también la presencia de Cyprideis nodulares por debajo y por encima de los niveles mezcla de fauna, indica fluctuaciones de los niveles de salinidad y cambios paleoambientales abruptos. Estos fósiles y as características morfológicas particulares de loes fosiles pueden estar relacionadas con una ingresión marina de corta duración. Esta influencia marina que llegó a la parte central de la Amazonia, probablemente puede tener ocurrido durante el Mioceno Inferior a Medio.
The possibility of marine incursions into the Amazonia during the Miocene has been the subject of discussion for a long time, while incomplete geological and paleontological data impedes the exact tracing of the proposed marine ingressions. However, previous records of marine taxa mixed with transitional and non-marine fossils, including foraminifers, marginal marine mollusks, shark teeth, rays, mangrove pollen, ostracods and barnacles, indicate that marine influence was present, at least episodically, in Amazonia during the Neogene although fluvial and lacustrine environments predominated.
Here we present micropaleontological analyses of core samples (core 1AS - 31-AM) from the southwest of Amazonas State, Brazil of that corroborate marine influence during the Miocene. The samples, from between 170.90 and 174.40 m depth yielded a mixture of marine (mainly based on planktonic foraminifers and bryozoans), transitional and non-marine faunal elements. Some of these organisms are resilient by stress conditions, such as the ostracod Cyprideis, some mollusks and agglutinated foraminifers. The radiation of Cyprideis and the presence of noded Cyprideis-valves below and above the mixed faunal levels point to abrupt paleoenvironmental change related to salinity fluctuations (from marine normal to non-marine). These fossils and the particular morphologic (ecophenotypic traits) features can be associated with a short-lived marine incursion. The presence of planktonic foraminifers indicates that marine influence must have been much more profound than previously reported. The marine influence reached the central part of Amazonia, probably during the Early Middle Miocene age.
Evidence for marine influx during the Miocene in southwestern Amazonia, Brazil
Evidencias de influencia marina durante el Mioceno en el Sudoeste de la Amazonia (Brasil)
Ana Paula Linhares 1, María Inês Feijó Ramos 1, Martín Gross 2 & Werner E. Piller3
1 Coordenação de Ciências da Terra e Ecologia, Museu Paraense Emílio Goeldi, Avenida Perimetral, s/n Terra Firme, Belém-PA 66077-830, Brazil; e-mail: mramos@museu-goeldi.br
2 Department for Geology and Palaeontology, Universalmuseum Joanneum, Weinzöttlstrasse 16, 8045 Graz, Austria; e-mail:martin.gross@museum-joanneum.at telephone: +43-316-8017-9733; fax: +43-316-8017-9671
3 Institute for Earth Sciences, Karl-Franzens-University, Heinrichstrasse 26, 8010 Graz, Austria; e-mail: werner.piller@uni-graz.at
Correspondent author: mramos@museu-goeldi.br
Manuscrito recibido: 5 de junio 2011; aceptado:15 de agosto 2011
Abstract
The possibility of marine incursions into the Amazonia during the Miocene has been the subject of discussion for a long time, while incomplete geological and paleontological data impedes the exact tracing of the proposed marine ingressions. However, previous records of marine taxa mixed with transitional and non-marine fossils, including foraminifers, marginal marine mollusks, shark teeth, rays, mangrove pollen, ostracods and barnacles, indicate that marine influence was present, at least episodically, in Amazonia during the Neogene although fluvial and lacustrine environments predominated.
Here we present micropaleontological analyses of core samples (core 1AS - 31-AM) from the southwest of Amazonas State, Brazil of that corroborate marine influence during the Miocene. The samples, from between 170.90 and 174.40 m depth yielded a mixture of marine (mainly based on planktonic foraminifers and bryozoans), transitional and non-marine faunal elements. Some of these organisms are resilient by stress conditions, such as the ostracod Cyprideis, some mollusks and agglutinated foraminifers. The radiation of Cyprideis and the presence of noded Cyprideis-valves below and above the mixed faunal levels point to abrupt paleoenvironmental change related to salinity fluctuations (from marine normal to non-marine). These fossils and the particular morphologic (ecophenotypic traits) features can be associated with a short-lived marine incursion. The presence of planktonic foraminifers indicates that marine influence must have been much more profound than previously reported. The marine influence reached the central part of Amazonia, probably during the Early Middle Miocene age.
Key-words: Marine Miocene, Brazilian Amazonia, ostracods, Solimões Formation.
Resumen
Probables incursiones marinas en la región Amazónica durante el Mioceno han sido objeto de debates durante mucho tiempo. La base de datos general, impide una exacta localización de las ingresiones marinas propuestas. Sin embargo, una mezcla de grupos marinos y de transición o no marinos como foraminíferos, moluscos marinos marginales, dientes de tiburónes, rayas, polen de los manglares, ostrácodos y percebes han sido registradas en estudios previos. Esto indica que la influencia marina puede haber jugado, al menos esporádicamente, algún papel en los depósitos neógenos de la Amazonia, a pesar que los ambientes fluviales y lacustres fueron los predominantes. Recientes estudios micropaleontológicos con muestras de testigo (1AS-31-AM, profundidad de 302 m; 05°18‘S / 71°02‘O; río Itui) del suroeste del estado Amazonas corroboran esta evidencia marina. Las muestras entre 170,90 y 174,40 metros de profundidad, contienen esporádicamente una mezcla de organismos marinos (principalmente basado en la presencia de foraminíferos y briozoarios), otra fauna de transición y también no marinos (principalmente de foraminíferos y ostrácodos). Aparte de estos elementos de la fauna, también la presencia de Cyprideis nodulares por debajo y por encima de los niveles de mezcla de fauna, indican fluctuaciones de los niveles de salinidad y cambios paleoambientales abruptos. Estos microfósiles y las características morfológicas particulares de los fosiles pueden estar relacionados con una ingresión marina de corta duración. Esta influencia marina que llegó a la parte central de la Amazonia, probablemente puede haber ocurrido durante el inicio del Mioceno Medio.
Palabras clave: Mioceno marino, Amazonia brasileña, ostrácodos, Formación Solimões.
INTRODUCTION
Neogene deposits are widespread in Amazonia. In Brazil, these deposits are represented by the Solimões Formation that reflects part of the orogenetic history of Western Amazonia during the Neogene. Considerable environmental changes took place during this period when the uplift of the Andes Mountains and sea-level fluctuations affected the hydrographical, ecological and climate systems of this region (Shephard et al. 2010).
Previous studies recorded the presence of marine, transitional and non-marine fossil taxa in the Solimões Formation and others contemporaneous units, which indicates that marine conditions, at least sporadically, influenced Amazonia during the Neogene (Shepard & Bate 1980; Hoorn 1993, 1994 a, b; Räsänen et al. 1995; Monsch 1998; Muñoz-Torres et al. 1998; Wesselingh et al. 2002, 2006; Gingras et al.. 2002; Hovikoski et al. 2010). However, a complex of fluvial and lacustrine environments generally predominated (Wesselingh et al. 2006; Ramos 2006; Lundberg et al. 2010; Hoorn 1996; Hoorn et al. 2010; Latrubesse et al. 2010; Gross et al. 2011), and the biostratigraphical correlation of different localities within the Solimões Formation remains problematic (see Hoorn & Wesselingh 2010).
Most investigations are restricted to only a few localities that cover relatively short stratigraphical range. The availability of core material from petroleum companies is usually difficult due to their policy of restrictive access, and is mostly limited to the western fringe of Amazonia where hydrocarbon resources are being explored. Most of the cores that have been analysed in detail originate from western Brazilian Amazonia where they were collected in the 1970‘s by the Geological Survey of Brazil (CPRM) to investigate the presence of lignite. The studies on these cores began with Purper (1979, employing ostracods), and was followed by Hoorn (1994a, 1994b, utilising pollen). The recent re-examination of these cores has focused mainly on palynological taxonomy, biostratigraphy and paleoecology (Leite 2006; Silva-Caminha et al. 2010; Latrubesse et al. 2007, 2010). The present paper presents a new micropaleontological analysis of samples from core 1AS-31-AM (05°18‘S/71°02‘W, 302 m depth), located near the Ituí River, southwest of Amazonas state, Brazil (Figura 1). The main aim is to reconstruct the paleoenvironmental evolution of this area during the Miocene and to attempt correlations with previous studies.
METHODOLOGY
The study material comprise 93 samples from core 1AS-31-AM (Figura 2), drilled on the margin of the Ituí river (Maia et al. 1977). The core reached a depth of 302.05 m; five cm of sediment was taken from each cored sampled interval. Samples were processed following conventional micropaleontological techniques (250g of dry sediment sieved through a ≥ 250µm size fractions). Microfossils were extracted from the sieve residual using a stereo microscope and then identified. Photographs were obtained by using LEO 1450VP SEM (Museu Paraense Emilio Goeldi, Belem, Para). The core samples were loaned by Departamento Nacional de Produção Mineral (8˚ Distrito, Manaus).

RESULTS AND DISCUSSIONS
The genus Cyprideis comprises 90% of the ostracod fauna from the Solimões Formation with high diversity and abundance (Muñoz-Torres et al. 1998, 2006). However, it also occurs with few marine, marginal- marine and freshwater taxa (e.g. Paracypris, Pontocypris, Perissocytheridea, Cytheridella, Cypria, Darwinula, Heterocypris) at different localities in the Neogene of Amazonia (Purper 1979; Shepard & Bate 1980; Whatley et al. 1998; Ramos 2006; Wesselingh & Ramos 2010). Ecophenotypes of Cyprideis can tolerate lower to higher salinities (1.5‰ to 60‰ Athersuch et al. 1989 fide Keyser et al. 2004) thriving in environments that can be physiologically stressful such as lagoons and inland ephemeral saltwater lakes (e.g., Whatley et al. 1998; Aladin & Potts 1996).
Cyprideis can develop nodes as a response to salinity fluctuation, generally linked to lower (<5‰) saline conditions (Keyser & Aladin 2002; Keyser 2005). According to Keyser (2005) the main cause of the nodding in low salinity waters is a failure in osmoregulation capacities during molting, which also can occurs in others genera like Heterocypris (see Keyser & Aladin 2002; Keyser 2005). Although Cyprideis is more usually associated with brackish water it also occurs in freshwater, especially the African lakes (Martens et al. 1994; Wouters and Martens 1992, 2001).
The Solimoes and Pannonian Basin of SW Europe are two important examples of adaptive radiations of Cyprideis during the Miocene (Whatley et al. 1998; Gross & Piller 2009). According to Whatley et al. (1998) the radiation of Cyprideis in Solimões Formation, occurred in a brackish closed hydrological system where extra-basinal species migration was limited. There appears to be no correlation between enhanced evolutionary activity and named extrinsic events.
As with other records, the microfauna of 1AS-31-AM is dominated Cyprideis, with 18 species, 11 recorded previously from others localities and seven new species to science (Figura 3, Figura 4 and Figura 5).



Cyprideis species occur from 297.50 m to 20.0 m depth. The highest diversity as well as the presence of noded Cyprideis-valves are found in samples below and just above of the marine incursion interval (at 175 and 170.80 m respectively, Figura 5). Besides the ostracodes, the microfossil assemblages analyses throughout the core allowed identifying three temporally variable salinity intervals (Figura 2): Lacustrine Phase (Interval I); Transitional to marine Phase (Interval II); and Fluvio- lacustrine Phase (Interval III).
Lacustrine Phase: Interval I (230.0-175.0 m) is dominated by the Cyprideis assemblage that consists of nine species (C. amazonica, C. caraionae, C. graciosa, C. machadoi, C. olivencai, C. pebasae, C. sulcosigmoidalis, C. sp. 6 and C. sp. 7) and 93 specimens that reduce abruptly by 180.9 m. Rare mollusks and fish remains occur associated to the ostracods.
The first occurrence of foraminifera (Amphistegina sp) is at 181 m. In the end of the Interval I, at 175.0 m, besides noded Cyprideis-valves the Cyprideis assemblage re- emerges with two new species, and a total of 11 species from 220 specimens. Also, at this depth, an otolith of typical marine fish was found (Aguilera et al. in press). It could indicate the first signals of the marine influence.
The Transitional to Marine Phase: Interval II (174.90- 170.90 m) is represented by a mixture of marine, transitional and non-marine taxa. This interval begins with the presence of marine mollusk Melongena woodwardii alongside a lower diversity (five species) and abundance of Cyprideis (C. amazonica, C. aulakos, C. graciosa, C. machadoi and C. pebasae). At 174.65 m and 174.40 m charophyte oogonia were found with a few specimens of Amphistegina spp., bryozoans and Cyprideis (C. aulakos, C. machadoi and C. pebasae). At 172.40 m, a more diverse assemblage occurs: benthic (Amphistegina spp. and Quinqueloculina) and agglutinated (Textularia) foraminifers, bryozoans, decapods, mollusks, fishes remains, rare non-marine ostracods (Candona and Cytheridella) and some species of Cyprideis (C. inversa, C. Olivencai, C. pebasae, C. sulcosigmoidalis, C. sp. 1, C. sp. 3 and C. sp. 5).
The Interval II, at 170.90m depth, finish with a typical marine microfauna dominated by planktonic (Globigerinoides, Globorotalia and Globigerina) and benthic foraminifers (Amphistegina, Planorbulina and Quinqueloculina), totalizing 21 specimens, as well as bryozoans (Figura 6). The presence of planktonic foraminifers at this depth indicates the peak of marine conditions within this sequence. At this depth, the others taxa (mollusks, fish remains, etc.) mentioned before and the ostracods disappear with only one remaining specimen of C. aulakos.
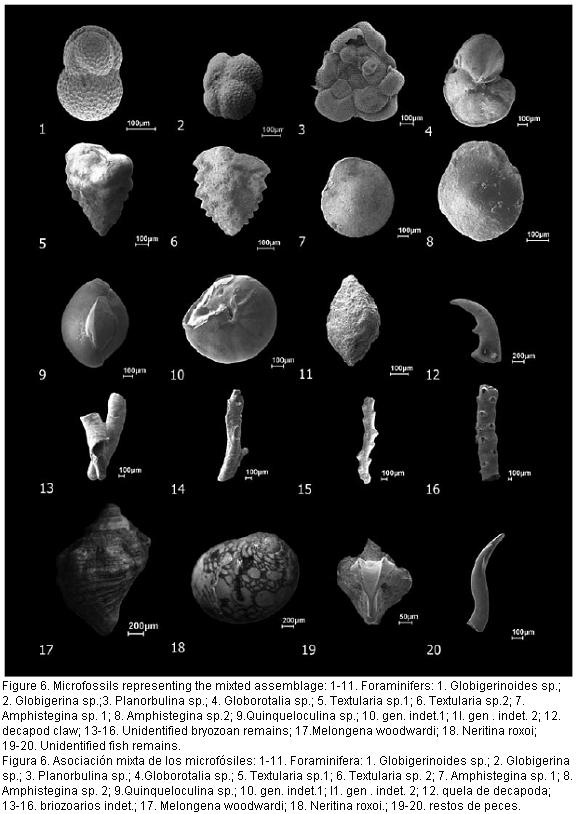
Fluvio-lacustrine Phase: Interval III (170.80-20 m) begin with the disappearance of marine taxa and the return of high Cyprideis diversity (C. amazonica, C. aulakos, C. caraionae, C. cyrtoma, C. graciosa, C. machadoi, C. olivencai, C. pebasae, C. sulcosigmoidales, C. sp. 1 and C. sp. 7); the presence of noded Cyprideis (Figura 4; 9-10) as well as agglutinated foraminifers and mollusks (e.g. Neritina roxoi) which are able to tolerant fluctuating salinities as well as fresh-water fishes remains suggests abrupt decrease of salinity water. From 157.25 m to 138.20 m the diversity of Cyprideis reduces and new fresh water ostracodes genera, such as Cypria and Darwinula arise indicating a freshening of waters. To the uppermost of this interval the ostracodes almost disappear, remaining only few freshwater species (Cypria aqualica, Darwinulla fragilis and Cytheridella sp.) and few Cyprideis (C. amazonica, C. graciosa and C. machadoi). This fresh water ostracod assemblage (Cypria, Cytheridella and Darwinulla) has been recorded in the Late Miocene (Solimões Formation) outcrops around Eirunepé city, in environments associated with a complex fluviolacustrine environment (Ramos, 2006; Gross et al. 2011). Recently, living Cypria and Cytheridella has been found in fresh-water natural floodplain lake (abandoned channels fill) in the Amazonia, closed to Eirunepé city (M.I.F.R. pers. comun.). Probably the endemic Cyprideis assemblage from Solimões Fm. was extinct at the end of Late Miocene.
The radiation of Cyprideis in this study is an excellent biological marker for abrupt salinity change. The number of Cyprideis species increase before and after the Transitional to Marine Phase (Interval II). When marine waters invade the lacustrine environment for the first time (at 174.90 m), the adaptation of Cyprideis begin rapidly resulting carapace noding. Noding of Cyprideis also occurred during the offstage, as region reverted to continental conditions. Whatley et al. (1998) suggest that the high diversity of Cyprideis, throughout the Miocene in Amazonia, occurred within a stable, isolated body of saline water; however, in the study core, radiation is directly associated with the salinities change caused by marine incursions. It represents that the radiation of Cyprideis do not occurred in a closed system.
Similar to the Cyprideis signature, Wesselingh et al. (2010) show an important mollusk turnover linked to marine incursions in Peru and Colombia. At the time the endemic bivalve Pachydon obliquus became replaced by P. trigonalis as the dominant bivalve species. Furthermore, several previously common gastropod species, such as Dyris tricarinatus disappeared and various novel species appeared. It is thus possible that the marine incursion played an important role in reshaping the invertebrate fauna of Amazonian lakes and wetlands.
Until recently, most authors suggested only marginal marine influence in the Miocene of western Amazonia (e.g., Hoorn 1995; Vonhof et al.. 1998; Vermeij & Wesselingh 2002; Gingras et al.. 2002; Wesselingh & Ramos 2010). Geochemical and faunal paleosalinities were estimated at most 5 psu (Vonhof et al.. 1998, 2003). Hovikoski et al. (2007, 2010) recorded within the Pebas Formation a low-gradient depositional system, influenced by recurring, shallow and restricted marine incursions.
The presence of planktonic foraminifera shows, however, that episodically paleosalinities must have been near normal marine and that the (probably short-lived) marine peak influx in Amazonia may have been much more profound than previously recognized.
The Transicional to Marine Phase (Interval II) is not younger than Early Middle Miocene based on the stratigraphical distribution of the C. caraione. According to the ostracod biozonation by Muñoz-Torres et al. (2006), the C. caraione occur since the Cyprideis aulakos zone (Late Early to Early Middle Miocene) to C. caraione zone (Early Middle Miocene) in the Langhian.
Although marine incursions was previously attributed to Late Middle Miocene (Grimsdalea zone of Hoorn,1995) based on the occurrence of marine mollusks (including Melongena woodwardi), barnacles and bryozoans in samples in localities Buenos Aires, Porvenir, Nuevo Horizonte from Colombia and Peru (Wesselingh et al.. 2010; Vonhof et al.. 1998), the estimated Early Middle Miocene age (~ 16 to 11.3 Ma) to the marine incursion in the present study is in agreement with the "Pebas Phase" by Hoorn et al. (2010).
This palaeoenvironmental reconstruction offers new insight into the evolution of the Amazonia depositional system. Micro and macro-faunal evidence clearly indicates a transition from continental (lacustrine) environments at the base to a short duration marine influenced and the re-establishment of fluvial conditions at the top of the sequence. The effect of this environmental change, mainly the abrupt marine incursion is probably related to tectonic activity and/or sea level rise rate change. Probably events of great magnitude that could allow the sea water reach so long distance into the continent. The radiation of Cyprideis is most likely triggered by these sudden environmental changes (mainly related to salinity change). These fossils and the particular morphologic (ecophenotypic traits) features infer that a short-lived marine incursions reached the central part of Amazonia, probably in the Early Middle Miocene.
The origin of the marine incursion has a good chance to be from Caribbean region, based on the fossil content, although a more detailed taxonomic study of taxa, is necessary in subsequent publications.
CONCLUSIONS
The microfossils from 93 samples of the 1-AS-31-AM core (southwestern Amazonas state, Brazil) allowed the reconstruction of paleoenvironmental conditions in southwestern Amazonia, Brazil during the Miocene. Three temporally variable salinity intervals are recognized: Lacustrine Phase (Interval I); Transitional to marine Phase (Interval II); and Fluvio-lacustrine Phase (Interval III). The succession began with lacustrine environments and graded into a short- live marine incursion and ending with the establishment of a fluviolacustrine system. The peak of the marine incursions at 170.90 m is evidenced by the occurrence of planktonic foraminifers, and the disappearance of brackish and non- marine taxa. The extent of the marine influence is much more profound than previously assumed. The short- lived marine incursion caused rapid changes in water salinities resulting in the radiation of the ostracod genus Cyprideis. The presence of nodes in Cyprideis valves as well as the presence of eurytopic microfossils is related to abrupt salinity change. This brief but profound marine incursion was probably caused by tectonic events of great magnitude that most likely took place during the Early Middle Miocene.
Acknowledgments
The authors thanks to CNPq for the financial support; DNPM (8˚ Distrito, Manaus) for the permission to access and loan the core samples and especially to Gert Woeltje for his assistance during the samples collecting; to Adrian Wood for the comments and the English review and finally to Frank Wesselingh and an other anonymous reviewer for the fruitful suggestions and corrections.
REFERENCES
Aguilera, O.S., Ramos, M.I.F., Linhares, A.P., Ohe, F. (in press): Miocene Marine episodes in Western Amazonia: An interpretation of Fossil Ariidae catfishes occurrence and their palaeobiogeographic significance. Palaeogeography, Palaeoclimatology, Palaeoecology.
Aladin, N. V. & Potts W.T.W. (1996): The osmoregulatory capacity of the Ostracoda. Journal of Comparative Physiology, 166: 215-222.
Gingras, M.K., Rasanen, M., Pemberton, G.S., Romero L.P. (2002): Ichnology and sedimentology reveal depositional characteristics of bay-margin parasequenses in the Miocene Amazonian foreland Basin. 2002. Journal of Sedimentary Research, 72 (6) 871-883.
Gross, M., Piller, W.E., Ramos, M.I.F., Paz, J.D.S. (2011). Late Miocene sedimentary environments in south-western Amazonia (Solimões Formation; Brazil). Journal of South American Earth Sciences,32: 169-181.
Hoorn, C. (1993). Marine incursions and the influence of Andean tectonics on the Miocene depositional history of northwestern Amazonia: results of a palynostratigraphic study. Palaeogeography, Palaeoclimatology, Palaeoecology, 105: 267-309.
Hoorn, C. (1994a): Fluvial palaeoenvironmental in the intracratonic Amazonas Basin (Early Miocene- Early Middle Miocene) Colombia. Palaeogeography, Palaeoclimatology, Palaeoecology, 109: I-54.
Hoorn, C. (1994b): An environmental reconstruction of the palaeo-Amazon River System (Middle- Late Miocene), NW Amazonia. Palaeogeography, Palaeoclimatology, Palaeoecology, 112: 187-238.
Hoorn, C. (1996): Miocene Deposits in the Amazonian Foreland Basin. Science, 273: 122-123.
Hoorn C. & Wesselingh F. (2010): Amazonia: Landscape and species evolution, a look into the past 447 p. Wiley-Blackwell publications, UK.
Hoorn, C., Wesselingh, F.P., Hovikoski, J., Guerrero, J. (2010): The development of the Amazonian mega-wetland (Miocene; Brazil, Colombia, Peru, Bolívia). In: Hoorn C. & Wesselingh F. (eds) Amazonia: Landscape and species evolution, a look into the past, Chapter 8: 123-142.Wiley-Blackwell publications, UK.
Hovikoski, J., Räsänen, M., Gingras, M., Lopez, S. Ranzi, A., Melo, J. (2007). Palaeogeographical implications of the Miocene Quendeque Formation (Bolívia) and tidally-influenced strata in the southwestern Amazonia. Palaeogeography, Palaeoclimatology, Palaeoecology, 243: 23-41.
Hovikoski, J., Wesselingh, F.P., Räsänen, M., Gingras, M., Vonhof, H.B. (2010): Marine influence in Amazonia: evidence from the geological record. In: Hoorn C. & Wesselingh F. (eds) Amazonia: Landscape and species evolution, a look into the past, Chapter 9: 143-161. Wiley-Blackwell publications, UK.
Keyser, D. & Aladin, N. (2002): Noding in Cyprideis torosa and its causes. Studia Quarternaria, 21: 19-24.
Keyser, D., Aladin, N., Plotnikov, I. (2004): Ostracods and their adjustment to saline environments. Mitt. Hamb. Zool. Mus. Inst. 101: 157-167.
Keyser, D. (2005): Histological peculiarities of the noding process in Cyprideis torosa (Jones) (Crustacea, Ostracoda). Hydrobiologia, 538: 95-106.
Latrubesse, E.M., Silva, S.A.F. da, Cozzuol, M., Absy, M.L. (2007): Late Miocene continental sedimentation in southwestern Amazonia and its regional significance: Biotic and geological evidence. Journal of South American Earth Sciences, 23: 61-80.
Latrubesse, E.M., Cozzuol, M., Silva-Caminha, S.A.F., Rigsby, C.A., Absy, M.L., Jaramillo, C.A. (2010): The Late Miocene paleogeography of the Amazon Basin and the evolution of the Amazon river system. Earth-Science Reviews, 99: 99-124.
Leite, F.P.R. (2006): Palinologia da Formação Solimões, Neógeno da Bacia do Solimões, Estado do Amazonas, Brasil: Implicações paleoambientais e bioestratigráficas (Tese de Doutorado). 128 p. Universidade de Brasília, Instituto de Geociências, Brasília.
Lundberg, J.G., Pérez, M.H.S., Dahdul, W.M., Aguilera, O.A.(2010): The Amazonian Neogene fish fauna. In: Amazonia: Landscape and Species Evolution (Hoorn, C. and Wesseling, F eds.), Chapter 17: 281-301.
Maia, R.G.N., Godoy, H. de O., Yamaguti, H.S., Moura, P.A., Costa, F.S.F. da, Holanda, M.A., Costa, J.A. (1977): Projeto Carvão no Alto Solimões; Relatório Final. Manaus: CPRM; DNPM, 1: 142 p.
Martens, K., Coulter, G. & Goddeeris, B. (1994): Speciation in ancient lakes-40 years after. Advances in Limnology, 44: 75-96.
Monsch, K.A. (1998): Miocene fish faunas from the northwestern Amazonia Basin (Colombia, Peru, Brazil) with evidence of marine incursions. Palaeogeography, Palaeoclimatology, Palaeoecology, 143: 31-50.
Muñoz-Torres, F., Whatley, R.C & Harten, D. van. (1998): The endemic non-marine Miocene ostracod fauna of the Upper Amazon Basin. Revista Espanola de Micropaleontologia, 30 (3): 89-105.
Muñoz-Torres, F., Whatley, R.C & Harten, D. van. (2006): Miocene ostracod (Crustacea) biostratigraphy of the Upper Amazon Basin and evolution of the genus Cyprideis. Journal of South American Earth Sciences, 21: 75-86.
Purper, I. (1979): Cenozoic Ostracods of the Upper Amazon Basin, Brazil. Pesquisas, 12: 209-281.
Radambrasil. (1977): Folha SB.19 Juruá: geologia, geomorfologia, pedologia, vegetação e uso potencial da terra. DNPM, Brasil, 436 p.
Ramos, M.I.F. (2006): Ostracods from the Neogene Solimoes Formation (Amazonas, Brazil). Journal of South American Earth Sciences, 21: 87-95.
Räsänen, M.E., Linna, A.M., Santos, J.C.R., Negri, F.R. (1995): Late Miocene Tidal Deposits in the Amazonian Foreland Basin. Science, 269: 386-390.
Shepard, L.M. & Bate, R.H. (1980): Plio-Pleistocene Ostracods from the Upper Amazon of Colombia and Peru. Palaeontology, 23: 97-124.
Shephard, G.E., Müller, R.D.& Gurnis, M. (2010): Miocene drainage reversal of the Amazon River driven by plate–mantle interaction. Nature Geoscience, 3, 870-875: 1-15.
Silva-Caminha, S.A.F., Jaramillo, C.A. & Absy, M.L. (2010): Neogene palynology of the Solimoes Basin, Brazilian Amazonia. Palaeontographica, Abteilung B: Palaeobotany – Palaeophytology, 283, 1-3: 1-67.
Vermeij, G.J. & Wesseling, F.P. (2002): Neo-Gastropod mollusks from the Miocene of Western Amazonia, with commnents on marine to fresh water transition on mollusks. J. Paleontol. 76: 265-270.
Vonhof, H. B., Wesselingh, F.B., Kaandorp, RJ., Davies, G.R, van Hinte, J.E., Guerrero, l., Rãsãnen, M, Romero-Pitman, L., Ranzi, A. (2003): Palaeogeog- raphy of Miocene Western Amazonia: isotopic com- position of molluscan shells constrain the influence marine incursions. Geological Society of American Bulletin, 20: 983-993.
Vonhof, H. B., F. P. Wesselingh & G.M. Gringas (1998): Reconstruction of the Miocene western Amazonian aquatic system using molluscan isotopic signatures. Palaeogeography, Palaeoclimatology, Palaeoecology 141: 85-93.
Wesselingh, F.B., Rãsãnen, M.E., Irion, G., Vonhof, H.B., Kaandorp, RJ., Renema, W., Romero-Pitman, L., Gingras, M. (2002): Lake Pebas: a palaeocological reconstruction of a Miocene, long-lived lake complex in western Amazonia. Cainozoic Research 1: 35-81.
Wesselingh, F.P., Hoorn, M.C., Guerrero, J., Räsänen, M. E., Romero Pittmann, L., Salo, J. (2006): The stratigraphy and regional structure of Miocene deposits in western Amazonia (Peru, Colombia and Brazil), with implications for Late Neogene landscape evolution. Scripta Geologica, 133: 291-322.
Wesselingh, F.P. & Ramos M.I. F. (2010): Amazonian aquatic invertebrate faunas (Mollusca, Ostracoda) in their development over the past 30 million years. In: Amazonia: Landscape and Species Evolution (Hoorn, C. and Wesseling, F. eds.), Chapter 18:302-316.
Whatley, R., Muñoz-Torres, F. & Harten, D. van. (1998): The Ostracoda of an isolated Neogene saline lake in the Western Amazon Basin. In: S. Crasquin- Soleau, Braccini, E. and Lethiers, F. (ed.). What About Ostracoda! Bulletin Centre Recherches Elf Exploration Production Mémoires, 20: 231-245.
Wouters, K. & Martens, K. (1992): Contribution to the knowledge of the tanganyikan cytheraceans, with the description of Mesocyprideis nom. nov., (Crus- tacea, Ostracoda). Bulletin Koninklijk Belgisch Instituut voor Natuurwetenschappen, Biologie, 62: 159-166.
Wouters, K. & Martens, K. (1994): Contribution to the knowledge of Cyprideis species flock (Crustacea, Ostracoda) of Tanganyika, with the description of three new species. Bulletin Koninklijk Belgisch Instituut voor Natuurwetenschappen, Biologie, 64:111-128.
Wouters, K. & Martens, K. (2001): On the Cyprideis species flock (Crustacea, Ostracoda) in Lake Tanganyika, with the description of four new species. Hydrobiologia, 450: 111–127.









