



Published
E. coli Lipopolysaccharide Decreases the Expression of Proteins of Tight Junctions in the Jejunum of Weaning Piglets
Lipopolisacárido de E. coli Disminuye la Expresión de Proteínas de Uniones Apretadas en Yeyuno de Lechones Posdestete
DOI:
https://doi.org/10.15446/rfnam.v67n2.44172Keywords:
Diarrhea, fever, digestive functions, gastrointestinal pathophysiology. (en)Diarreas, fiebre, funciones digestivas, fisiopatología gastrointestinal. (es)
In order to assess the effect of the addition of E. coli lipopolysaccharide (LPS) on the expression of tight junction proteins in the jejunum of weaned piglets (the animals were weaned at 21-days-old), 64 piglets were slaughtered at 1, 5, 7 and 10 days postweanig and complete extraction of jejunum was carried out in order to evaluate the molecular expression of Claudine 3 (C3), Claudine 4 (C4), and Zonula Ocludens-1 (ZO-1). To induce intestinal inflammation, animals were fed a basal diet supplemented with four levels of E. coli LPS (0, 0.3, 0.5, and 1.0 mg mg-1 of feed). The statistical design consisted of randomized blocks in a 4X4 factorial arrangement (four LPS concentrations and four post-weaning periods). A decrease was observed (P<0.01) in the expression of C3, C4, and ZO-1 in the animals that were fed the diet with the highest LPS-inclusion level. LPS contributes to the appearance of anatomical and intestinal functional disorders which are denoted by a decrease in the molecular expression of proteins from the intestinal barrier. This, in turn, is associated with an indiscriminate paracellular transport of molecules, microorganisms, and toxic compounds into the circulatory system. This results in post-weaning diarrhea and a decrease in the productive efficiency of piglets.
E. Coli Lipopolysaccharide Decreases the Expression of Proteins of Tight Junctions in the Jejunum of Weaning Piglets
Lipopolisacárido de E. Coli Disminuye la Expresión de Proteínas de Uniones Apretadas en Yeyuno de Lechones Posdestete
Johana Andrea Ciro Galeano1; Albeiro López Herrera2 and Jaime Parra Suescún3
1 Auxiliar Professor. Universidad Autónoma de las Américas. Ph.D. student in Agricultural Sciences. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Grupo de Investigación BIOGEM. A.A. 1779, Medellín, Colombia. <johana.ciro@uam.edu.co>
2 Associate Professor. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Grupo de Investigación BIOGEM. A.A. 1779, Medellín, Colombia. <alherrera@unal.edu.co>
3 Assistant Professor. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agrarias - Grupo de Investigación BIOGEM. A.A. 1779, Medellín, Colombia. <jeparrasu@unal.edu.co>
Received: July 31, 2013; accepted: February 23, 2014.
DOI: https://doi.org/10.15446/rfnam.v67n2.44172
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.

Abstract. In order to assess the effect of the addition of E. coli lipopolysaccharide (LPS) on the expression of tight junction proteins in the jejunum of weaned piglets (the animals were weaned at 21-days-old), 64 piglets were slaughtered at 1, 5, 7 and 10 days postweanig and complete extraction of jejunum was carried out in order to evaluate the molecular expression of Claudine 3 (C3), Claudine 4 (C4), and Zonula Ocludens-1 (ZO-1). To induce intestinal inflammation, animals were fed a basal diet supplemented with four levels of E. coli LPS (0, 0.3, 0.5, and 1.0 mg mg-1 of feed). The statistical design consisted of randomized blocks in a 4X4 factorial arrangement (four LPS concentrations and four post-weaning periods). A decrease was observed (P<0.01) in the expression of C3, C4, and ZO-1 in the animals that were fed the diet with the highest LPS-inclusion level. LPS contributes to the appearance of anatomical and intestinal functional disorders which are denoted by a decrease in the molecular expression of proteins from the intestinal barrier. This, in turn, is associated with an indiscriminate paracellular transport of molecules, microorganisms, and toxic compounds into the circulatory system. This results in post-weaning diarrhea and a decrease in the productive efficiency of piglets.
Key words: Diarrhea, fever, digestive functions, gastrointestinal pathophysiology.
Resumen. Con el objetivo de evaluar el efecto de la adición de lipopolisacárido LPS de E. coli sobre la expresión de proteínas de las uniones apretadas en yeyuno de lechones destetos, se sacrificaron 64 lechones escalonadamente los días 1 (21 días de edad, día del destete), 5, 7 y 10 posdestete, y se les extrajo completamente el yeyuno para la evaluación de la expresión molecular de Claudina 3 (C3), Claudina 4 (C4), y Zonula Ocludens-1 (ZO-1). Para inducir la inflamación intestinal los animales fueron alimentados con una dieta basal, adicionada con cuatro niveles de LPS de E. coli (0; 0,3; 0,5 y 1,0 mg mg-1 de alimento). El diseño estadístico utilizado fue de bloques al azar en arreglo factorial de 4X4 (cuatro concentraciones de LPS en cuatro tiempos posdestete). Se observó la disminución (P<0,01) en la expresión de C3, C4 y ZO-1 en los animales que consumieron la dieta con mayor nivel de inclusión de LPS. El LPS contribuye a desordenes anatómicos y funcionales a nivel intestinal, representados por la disminución en la expresión molecular de proteínas de la barrera intestinal, lo que está asociado al transporte paracelular indiscriminado de moléculas, microorganismos y compuestos tóxicos hacia la circulación sistémica. Todo lo anterior conlleva la presentación de diarreas posdestete y a la disminución en la eficiencia productiva de los lechones.
Palabras clave: Diarreas, fiebre, funciones digestivas, fisiopatología gastrointestinal.
In the countries with the higher levels of pig production, piglets are weaned 14 to 28 days after being born (Marion et al., 2002). As a result, piglets are lighter and the digestive system is less developed during weaning, particularly the gut. This renders these animals more susceptible to digestive problems in the post-weaning period (Reis et al., 2007).
Besides its digestive functions, the gut of piglets forms a physical barrier that prevents toxic compounds and pathogens from entering the intestinal mucosa and the circulatory system. This barrier is mainly composed of tight junctions, which are multiprotein complexes that bind and seal the spaces between adjacent enterocytes and limit the paracellular movement of molecules and luminal bacteria within the mucosa of the tissues (Lamb-Rosteki et al., 2008). Junctions are formed by the proteins: Zonula Ocludens 1 (ZO-1) and two Claudines (C3 and C4), which are connected to the epithelial cells (Tlaskalová-Hogenová et al., 2004), act as a structure, and regulate the permeability of the tight junctions (McLaughlin et al., 2004).
Similarly, tight junctions participate in the preservation of cellular polarity and are considered to play a key role in intestinal diffusion mechanisms (Shen and Turner, 2005). However, diffusion mechanisms may be dramatically altered by the physiological processes triggered by weaning and the presence of microbial infections (Fan, 2002).
During the weaning period, there is a brief period of fasting resulting from the forced removal of milk and the subsequent provision of solid food. This may cause a decrease in nutrient absorption and an increase in the substrate available for enteropathogenic bacteria in the hindgut (Pluske et al., 2003). Consequently, weaning is associated with a dramatic change in the composition of the intestinal microbiota (Chow et al., 2010).
In modern production systems, early weaning causes the lactobacilli population to disappear from the gastrointestinal tract and favors an increase in the E. coli population, releasing proinflammatory substances such as lipopolysaccharides (LPS) from the cell walls (Amador et al., 2007). LPS causes physical damage in the gut by activating a coordinated series of cell signaling pathways which favor an inflammatory response, which in turn produces important changes in the structure and functional capacity of the gut (García et al., 2008). These intestinal changes are characterized by an alteration in the functions of the tight junctions as well as by an increase in the indiscriminate paracellular transport of molecules (Pitman and Blumberg, 2000; García et al., 2003).
The administration of E. coli LPS is one of the most used models to study acute infection processes because the LPS response is highly reproducible, without the side effects commonly associated with chronic infections. Therefore, the aim of this study was to determine the effect of E. coli LPS administration on the molecular expression of tight junction proteins in the jejunum of piglets at different post-weaning periods.
MATERIALS AND METHODS
Ethical considerations. All experimental procedures were conducted according to guidelines suggested by “The International Guiding Principles for Biomedical Research Involving Animals” (CIOMS, 1985), and approved by the Ethics Committee on Animal Experimentation of the Universidad Nacional de Colombia (Medellín; CEMED 001 from January 26, 2009).
Location. The fieldwork was conducted at the San Pablo Center of the Universidad Nacional de Colombia, located in Rionegro, Antioquia, at an altitude of 2,100 m, corresponding to a tropical lower-montane wet forest (LM-wf) zone, with average annual temperatures between 12 and 18 °C.
Animals. A total of 52 pigs were used. The pigs were group-penned in nursery crates with trough-type feeders in a temperature-controlled room (26 ± 3 °C).
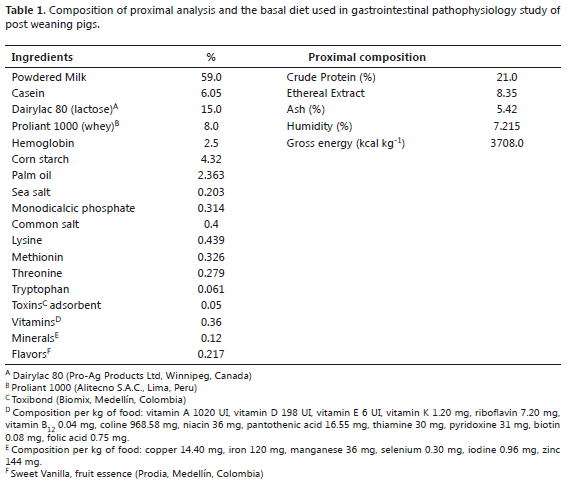
Diets. Four experimental diets were evaluated: a control diet (basal diet) and three others containing E. coli LPS, serotype 0111: B4 (Sigma-AldrichTM, St Louis, MO, USA), as follows:
- Basal Diet (BD) 0.0 mg LPS mg-1 feed.
- Diet 1 (D1): BD plus 0.3 mg LPS mg-1 feed.
- Diet 2 (D2): BD plus 0.5 mg LPS mg-1 feed.
- Diet 3 (D3): BD plus 1.0 mg LPS mg-1 feed.
The basal diet consisted of powdered milk and milk byproducts and was supplemented with vitamins, minerals, and HCL-lysine, meeting all minimum nutritional requirements proposed by the NRC (2012) (Table 1). The pigs were offered a minimum of 3000 g feed/day per crate, additional feed was provided when required. The diets were administered from days 1 to 10 after weaning. Water was provided ad-libitum throughout the experiment. No solid feed was offered to the piglets during lactation.
Small intestine sampling. The pigs were sequentially slaughtered as follows:
Four pigs were slaughtered on day 1 (weaning day), representing the control group. This group was used to assess the overall health status and to evaluate the macroscopic appearance of the organs before the pigs received LPS. Four pigs of each LPS-inclusion level (0, 0.3, 0.5, and 1.0 mg LPS mg-1 feed) were slaughtered on days five, seven, and 10 after weaning. All animals were slaughtered 2.5 hours after their last meal and were previously sedated by carbon dioxide inhalation for three minutes. The pigs were then slaughtered by exsanguination through a section of the jugular vein. After the slaughter, the small intestine was removed from the pyloric junction to the ileocecal valve (Segalés and Domingo, 2003). The intestine was lined up on a table, measured without any tension, divided into three equally sized sections (duodenum, jejunum, and ileum) and a 20 cm sample was taken from the center of the jejunum. The digesta contained in the samples was removed by washing it with cold saline infusion as previously described (Makkink et al., 1994; Reis et al., 2005). Subsamples (1 cm) were then obtained from each segment and were prepared for total RNA extraction. The samples were preserved in liquid nitrogen and the RNA was stored in a freezer at -70 oC.
Total RNA extraction and material quality check. Total RNA was extracted using an ULTRACLEANTM Tissue and Cells RNA Isolation Kit (MO BIO Laboratories Inc., San Diego, CA, USA). Samples (25 mg) were broken down in a mortar using liquid nitrogen. The RNA was stored at -80 °C and only aliquoted on ice for analysis. The integrity of the extracted material was assessed by running the samples on agarose gel with 1% ethidium bromide (100V for 50 min) and observing the rRNA bands. When not enough of the material or bands was clearly observed, the extraction was repeated under identical conditions (Freeman et al., 1999).
Synthesis of copy DNA (cDNA) from total RNA. A QuantiTectTM Reverse Transcription Kit (QIAGENTM) was used to synthesize the cDNA from the total RNA extracted. 1mg RNA samples were used for all reverse transcription reactions. Genomic DNA debris was removed and enzyme plus triphosphate nucleotides were added to synthesize the cDNA following the manufacturer's recommendations. The obtained cDNA was stored at -20 °C until it was used (Kubista et al., 2006).
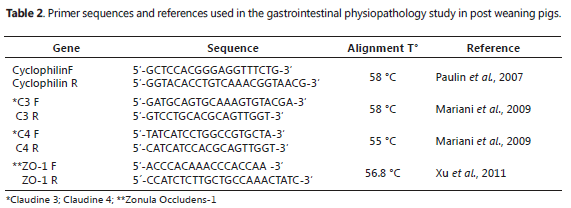
RT-PCR from cDNA for ZO-1, C3 and C4 and constitutive expression gene. The primers of the genes of interest and constitutive expression (Cyclophilin) are shown in Table 2. These primers were taken from the literature, verifying that their design had been performed on the intron/exon edges to avoid amplification of genomic DNA. Additionally, the primers were evaluated through bio-informatic analysis to verify their specificity with the studied mRNA and rRNA fragment and the absence of secondary structures, such as lenses or dimer formations, between them or with the other primers. The primers were chosen to amplify the less than 150 base pairs fragments, as recommended by some researchers (Bustin, 2000; Kubista et al., 2006). Standardization strategy, data processing, and relative quantification. To determine the expression level of each gene, a relative quantification method using a constitutively expressed gene (Cyclophilin) was used, following recommendations from several reports on gene choice and overall normalization strategy (Muller et al., 2002; Vandesompele et al., 2002; Huggett et al., 2005). The amount of material from which the RNA was extracted, the RNA amount in the reverse transcription reaction, and the amount of cDNA for PCR amplification were identical for all samples. The chosen constitutively expressed gene was validated in previous trials under similar experimental conditions (Paulin et al., 2007). The results were analyzed using the ImageJ software, version 1.43.
mRNA expression of intestinal ZO-1, C3 and C4 using RTPCR and quantification of PCR products. The mRNA amplification of C3, C4 and ZO-1 was conducted on individual samples of the middle intestine segment (Petersen et al., 2001). The general protocol for PCR was: Cyclophilin: 95 ºC x 1 min; 37 cycles 95 ºC x 30 seg; 58 ºC x 30 seg; 72 ºC x 40 seg; 72 ºC x 3 min; 10 ºC x ∞; C3: 95 ºC x 1 min; 39 cycles 95 ºC x 30 seg; 58 ºC x 30 seg; 72 ºC x 40 seg; 72 ºC x 3 min; 10 ºC x ∞; C4: 95 ºC x 1 min; 37 cycles 95 ºC x 30 seg; 55 ºC x 30 seg; 72 ºC x 40 seg; 72 ºC x 3 min; 10 ºC x ∞; ZO-1: 95 ºC x 1 min; 39 cycles 95 ºC x 30 seg; 56.8 ºC x 30 seg; 72 ºC x 40 seg; 72 ºC x 3 min; 10 ºC x ∞.
The density of the RT-PCR products from each protein was expressed in units relative to the band density of the constitutive expression gene (cyclophilin gene).
Statistical design: The experiment was performed as a randomized block design (2 blocks) in a 4x4 factorial arrangement (four experimental diets and four periods after weaning) (Steel and Torrie, 1980). The animals were blocked by initial weight. Each animal was assigned one of 16 treatments and each treatment had four repetitions. The General Linear Model procedure (GLM) of SASTM (2006) was used for the statistical analysis of the data. The Duncan test (P<0.05) was used to separate the treatment means.
RESULTS
Pigs fed the basal diet maintained normal body temperature, while animals that received LPS had rectal temperatures above 38 ºC throughout the experiment. However, pigs fed LPS did not show symptoms of disease, so none had to be slaughtered or excluded from the experiment. No food orts were observed during the experiment.
Data on the molecular expression of each of the genes for the studied proteins were obtained from the ratio between the expression of each protein's gene and the expression of the constitutive gene (Cyclophilin). Initially, the values for the molecular expression of the tight junction proteins in the basal diet were compared through the various post-weaning periods in order to determine the exclusive effect of weaning on the expression of these proteins. Table 3 shows the data obtained with the basal diet (BD) during the various experimental periods for the jejunum.

For C3, C4, and ZO-1, a statistically significant (P<0.01) decrease was observed in the molecular expression, starting from the first day after weaning (day 1). Similarly, on day 5 after weaning, the animals presented the lowest expression values in terms of cyclophilin (1.34, 1.18, and 0.90 respectively). All studied variables showed partial recovery from days five to ten. However, a statistical difference was observed between days 1 and 10 (P<0.01).
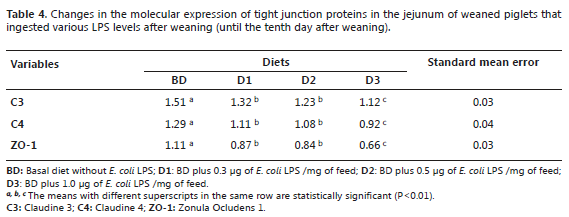
For tight junction proteins C3, C4, and ZO-1, significant decreases (P <0.01) were observed between the different diets (Table 4). The animals on the D3 diet had the lower molecular expression values (1.12, 0.92, and 0.66 respectively) with respect to BD (1.51, 1.29, and 1.11, respectively). However, it is worth mentioning that there were no statistically significant differences between D1 and D2 (P>0.01).
Tables 5 and 6 show the general average values of the molecular expression of the studied tight junction proteins for each treatment and for each exposure period. No statistical interaction was observed between the LPS concentrations and postweaning periods for any of the studied variables. Therefore, it was not necessary to analyze or break down said factors independently.
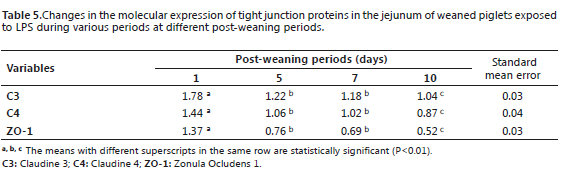
With regard to the molecular expression parameter for the C3, C4, and ZO-1 tight junctions during the LPS exposure periods, the overall evaluation of all the animals (DB, B1, D2, and D3; Table 5) showed a significant decrease (P<0.01) from day one after weaning. Additionally, the animals presented the lowest values at day 10 after weaning (1.04, 0.87, and 0.52 respectively).
DISCUSSION
The intestinal barrier is formed by many tight junctions where the proteins ZO-1, C3, and C4 are responsible for restricting the paracellular passage of luminal bacteria and molecules within the mucosa of the tissues (McLaughlin et al., 2004). When the intestinal microbiota has an optimal balance, it prevents pathogen or virulent microorganisms from multiplying and adhering to and invading the epithelial cells, as well as the circulatory system (Tlaskalová-Hogenová et al., 2004), which favors the integrity of the intestinal wall. However, under pathophysiological situations or stress, the MLCK pathway is activated, which increases intestinal permeability, solute cotransport, and the presence of proinflammatory cytokines and pathogenic bacteria.
Many cytokines, including TNF-a, are capable of increasing intestinal permeability because they can directly activate the MLCK (myosin regulatory light chain) pathway, thus shortening the actin filaments in the enterocytes, which ultimately breaks the tight union (Chin et al., 2002). A change in this conformation favors the development of pathogenic problems at the intestinal level (Saitou et al., 2000). Since tight junctions are important for maintaining the structural integrity of the epithelial barrier, they are the target of multiple infectious agents (Shen and Turner, 2005). Furthermore, these cytokines increase intestinal permeability by reducing the expression of the genes for proteins, such as ZO-1 and Claudines in the tight junctions, thus causing deep alterations in the normal physiology of enterocytes (Tsukita and Furuse, 2002; Bruewer et al., 2003). The results obtained in this study showed a reduction in the molecular expression of ZO-1, C3, and C4, which may be associated with the post-weaning physiological intestinal inflammation in the piglets fed with the basal diet.
The events in which the MLCK pathway is activated are associated with a decrease in transepithelial resistance (Berkes et al., 2003), an increase in indiscriminate paracellular transport of molecules, and a bacterial invasion of tissues (Shifflett et al., 2005). Thus, the modulation of MLCK activity represents a point of convergence of multiple signaling pathways regulating the function of the tight junctions, thus causing changes in their structure and, in turn, in the proteins that compose said junctions (Yu et al., 2010).
As mentioned above, the decrease in the molecular expression of the tight junction proteins observed in the animals that consumed the BD might be associated with the inflammatory and immune responses produced by different stress manifestations during weaning. These include: abrupt separation from the mother, relocation to new social groups, and changing to solid food (Kojima et al., 2007). In addition, physiological responses to the stress caused by weaning involve complex interactions and responses from the central nervous, endocrine, and immune systems, which influence the health and welfare of the animals in response to environmental and handling conditions (Yang and Glaser, 2000; Davis et al., 2006).
In this study, a decrease in the molecular expression of tight junction proteins was observed in the pigs that were fed with diets containing increasing concentrations of LPS. In the studies conducted by Pié et al. (2004), Amador et al. (2007), and Parra et al. (2013), the addition of LPS to the intestinal medium increased the production of various proinflammatory cytokines, mainly TNF-a, which may activate a wide range of signaling pathways that affect cell turnover and growth by stimulating apoptosis (Bruewer et al., 2003), thus favoring permeability to bacteria and compounds that might strengthen infectious and inflammatory processes (Berkes et al., 2003). In addition, McLaughlin et al. (2004) reported that TNF-a modifies the structure of claudines by weakening the multiprotein complex of the tight junctions, which is crucial for the maintenance of intestinal barrier properties. This is consistent with the results obtained in this study, which showed that LPS decreases the expression of ZO-1, C3, and C4, possibly due to an increase in the production of TNF-a, as reported by Parra et al. (2013).
Another alternate mechanism involved in the intestinal alterations resulting from the TNF-a production and from the inflammation caused by LPS (Hataya et al., 2003; Waseem et al., 2008, Parra et al., 2013) is the shortening of the actin filaments in the enterocytes, which makes the structure of the tight junctions permeable to bacteria and compounds that might strengthen infections and inflammations (Berkes et al., 2003). This favors the infiltration of leukocytes, the expression of the gene for proinflammatory cytokines, the overexpression of heat shock proteins, the modification of tissues caused by proteases, and, finally, the appearance of epithelial functional disorders related to absorption, mineral secretion, and intestinal permeability (Lallès et al., 2004).
Studies conducted by Lamb-Rosteki (2008) demonstrated that the integrity and function of the epithelial barrier is interrupted after one hour of exposure to C. jejuni in broilers. The interruption of such barrier is represented by the paracellular breaking and the granular accumulation of Claudines. This is consistent with this study's findings since, as the time of exposure to E. coli LPS increased, a decrease in the molecular expression of C3, C4, and ZO-1 was observed, as well as a reduction in the integrity and renewal rate of the epithelial barrier's multiprotein complex. On the other hand, exposure to some toxic secondary metabolites produced by fungi (deoxynivalenol) decreases C3 in caco-2 cells, as well as C3 and C4 in IPEC-1 cells, leading to the alteration of the structure of the tight junctions, which are responsible for the paracellular passage of molecules (McLaughlin et al., 2004; Pinton et al., 2009). Therefore, the decrease in the expression of proteins C3, C4, and ZO-1 observed in this study might cause the paracellular movement of compounds and a decrease in cell polarity, which would ultimately favor the appearance of diarrhea (Mush et al., 2006).
CONCLUSIONS
Weaning is associated with multiple environmental and nutritional factors that produce different manifestations of stress, which in turn cause the protein complexes forming tight junctions to deteriorate, thus decreasing intestinal barrier functioning.
E. coli LPS decreases the expression of tight junction proteins, which might contribute to the development of anatomical and functional disorders in the intestine. Such disorders are characterized by indiscriminate paracellular transport of molecules, microorganisms and toxic compounds into the circulatory system. This results in post-weaning diarrhea and a decrease in the productive efficiency of piglets.
Due to the large anatomical, physiological, and immunological similarities between human beings and pigs, the latter might be an excellent animal model for developing therapeutic strategies in human medicine. Likewise, these results could be a starting point in the search for therapies for Crohn's disease in human beings.
BIBLIOGRAPHY
Amador, P., J. García, M.C. Marca, J. de la Osada, S. Acín, M.A. Navarro, M.T. Salvador, M.P. Lostao and M.J. Rodríguez-Yoldi. 2007. Intestinal D-galactose transport in an endotoxemia model in the rabbit. The Journal of Membrane Biology 215(2-3): 125-133.
Berkes, J., V.K. Viswanathan, S.D. Savkovic and G. Hecht. 2003. Intestinal epithelial responses to enteric pathogens: effects on tight junction barrier, ion transport, and inflammation. Gut 52(3): 439-451.
Bruewer, M., A. Luegering, T. Kucharzik, C.A. Parkos, J.L. Madara, A.M. Hopkins and A. Nusrat. 2003. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. Journal of Immunology 171(11):6164-72.
Bustin, S.A. 2000. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. Journal of Molecular Endocrinology 25(2): 169-193.
Chin, A.C., D.A. Teoh, K.G. Scott, J.B. Meddings, W.K. Macnaughton and A.G. Buret. 2002. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infection and Immunity 70(7): 3673-3680.
Council for International Organizations of Medical Sciences. (CIOMS). 1985. International Guiding Principles for Biomedical Research Involving Animals. Geneva. 28 p.
Chow, J., S.M. Lee, Y. Shen, A. Khosravi and S.K. Mazmanian. 2010. Host-bacterial symbiosis in health and disease. Advances in Immunology 107: 243-274.
Davis, M.E., S.C. Sears, J.K. Apple, C.V. Maxwell and Z.B. Johnson. 2006. Effect of weaning age and commingling after the nursery phase of pigs in a wean-to-finish facility on growth, and humoral and behavioral indicators of well-being. Journal of Animal Science 84: 743-756.
Fan, M.Z. 2002. Growth and ontogeny of the gastrointestinal tract. pp. 31-60. In: Xu, R.J. and P. Cranwell (eds.). The neonatal pig: gastrointestinal physiology and nutrition. Nottingham University Press, Nottingham. 360 p.
Freeman, W.M., S.J. Walker and K.E. Vrana. 1999. Quantitative RT-PCR: pitfalls and potential. Biotechniques 26(1): 112-22.
García, J., B. Abad and M.J. Rodríguez-Yoldi. 2003. Effect of lipopolysaccharide on D-fructose transport across rabbit jejunum. Inflammation Research 52(4): 177-184.
García, J, M.C. Marca, E. Brot-Laroche, N. Guillén, S. Acin, M.A. Navarro, J. Osada and M.J. Rodríguez-Yoldi. 2008. Protein kinases, TNF-a, and proteasome contribute in the inhibition of fructose intestinal transport by sepsis in vivo. American Journal of Physiology. Gastrointestinal and Liver Physiology 294(1): G155-G164.
Hataya, Y., T. Akamizu, H. Hosoda, N. Kanamoto, K. Moriyama, K. Kangawa, K. Takaya and K. Nakao. 2003. Alterations of plasma ghrelin levels in rats with lipopolysaccharide-induced wasting syndrome and effects of ghrelin treatment on the syndrome. Endocrinology 144: 5365-5371.
Huggett, J., K. Dheda, S. Bustin and A. Zumla. 2005. Real-time RT-PCR normalisation; strategies and considerations. Genes and Immunity 6(4): 279-84.
Kojima, C.J., J.A. Carroll, R.L. Matteri, K.J. Touchette and G.L. Allee. 2007. Effects of weaning and weaning weight on neuroendocrine regulators of feed intake in pigs. Journal of Animal Science 85(9): 2133-2139.
Kubista, M., J.M. Andrade, M. Bengtsson, A. Forootan, J. Jonák, K. Lind, R, Sindelka, R. Sjöback, B. Sjögreen, L. Strömbom, A. Ståhlberg and N. Zoric. 2006. The real-time polymerase chain reaction. Molecular Aspects of Medicine 27(2-3): 95-125.
Lallès, J.P., G. Boudry, C. Favier, N. Le Floc'h, S. Pié, C. Piel and B. Sève, 2004. Gut function and dysfunction in young pigs: physiology. Animal Research 53: 301- 316.
Lamb-Rosteki, J., L. Kalischuk, G. Inglis and A. Buret. 2008. Epidermal growth factor inhibits Campylobacter jejuni-induced Claudin-4 disruption, loss of epithelial barrier funtion, and Escherichia coli translocation. Infection and Immunity 76(8): 3390-3398.
Makkink, C.A., P.J.M. Berntsen, B.M.L. op den Kamp, B. Kemp and W.A. Verstegen. 1994. Gastric protein breakdown and pancreatic enzyme activities in response to two different dietary protein sources in newly weaned pigs. Journal of Animal Science 72(11): 2843-2850.
Mariani, V., S. Palermo, S. Fiorentini, A. Lanubile and E. Giuffra. 2009. Gene expression study of two widely used pig intestinal epithelial cell lines: IPEC-J2 and IPI-2I. Veterinary Immunology and Immunopathology 131(3-4): 278-284.
Marion, J., M. Biernat, F. Thomas, G. Savary, Y. Le Breton, R. Zabielski, I. Le Huërou-Luron and J. Le Dividich, 2002. Small intestine growth and morphometry in piglets weaned at 7 days of age. Effects of level of energy intake. Reproduction, Nutrition, Development 42(4): 339-354.
McLaughlin, J., P. Padfield, J. Burt and C. O´Neil. 2004. Ochratoxin a increases permeability through tight juntions by removal of specific claudin isoforms. American Journal of Physiology. Cell physiology 287(5): C1412-C1417.
Muller, P.Y., H. Janovjak, A.R. Miserez and Z. Dobbie. 2002. Processing of gene expression data generated by quantitative real time RT-PCR. Biotechniques 32(6): 1372-1374, 1376, 1378-1379.
Musch, M.W., M.M. Walsh-Reitz and E.B. Chang. 2006. Roles of ZO-1, occludin, and actin in oxidant-induced barrier disruption. American Journal of Physiology and Gastrointestinal Liver Physiology 290: G222-G23.
National Research Council (NRC). 2012. The nutrient requirements of swine. Eleventh Revised Edition. The National Academy Press, Washington. 400 p.
Parra, S.J.E., T.J. Agudelo, P.D. Sanín, DJ. Forero, L.C. Muskus and H.A. López. 2013. Intestinal expression of pro-inðAammatory cytokines induced by oral intake of lipopolysaccharide (LPS) from E. coli in weaned pigs. Revista Colombiana de Ciencias Pecuarias 26: 108-118.
Paulin, S.M., A. Jagannathan, J. Campbell, T.S. Wallis and M.P. Stevens. 2007. Net replication of Salmonella enterica serovars Typhimurium and Choleraesuis in porcine intestinal mucosa and nodes is associated with their differential virulence. Infection and Immunity 75(8): 3950-3960.
Petersen, Y.M., D.G. Burrin and P.T. Sangild. 2001. GLP-2 has differential effects on small intestine growth and function in fetal and neonatal pigs. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology 281(6): 1986-1993.
Pié, S., S.P. Lallès, F. Blazy, J. Laffitte, B. Sève and I.P. Oswald. 2004. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. Journal of Nutrition 134: 641-647.
Pinton, P., J. Nougayréde, J. Del Rio, C. Moreno, D. Marin, L. Ferrier, A. Bracarence, M. Kolf-Clauw and I. Oswald. 2009. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicology and Applied Pharmacology 237(1): 41-48.
Pitman, R.S. and R.S. Blumberg. 2000. First line of defense: the role of the intestinal epithelium as an active component of the mucosal immune system. Journal of Gastroenterology 35(11): 805-814.
Pluske, J.R., D.E. Hopwood y D.J. Hampson. 2003. Relación entre la microbiótica intestinal, el pienso y la incidencia de diarreas, y su influencia sobre la salud del lechón tras el destete. pp. 94-108. En: XIX Curso de Especialización FEDNA, Madrid.
Reis, S.T.C., C.M.J. Guerrero, B.A. Aguilera y L.G. Mariscal. 2005. Efecto de diferentes cereales sobre la morfología intestinal de lechones recién destetados. Técnica Pecuaria en México 43(3): 309-321.
Reis, S.T.C., L.G. Mariscal, B.A. Aguilera y J.G.H. Cervantes. 2007. Digestibilidad de la proteína y energía en dietas para lechones complementadas con tres diferentes tipos de suero de leche deshidratado. Revista Veterinaria México 38(2): 141-151.
SASTM. 2006. SAS/STAT User's Guide. Institute Inc. Statistical Analysis Systems Institute. Version 9.1th edition. Cary, NC: SAS Institute Inc.
Saitou, M., M. Furuse, H. Sasaki, J.D. Schulzke, M. Fromm, H. Takano, T. Noda and S. Tsukita. 2000. Complex phenotype of mice lacking occludin, a component of tight junction strands. Molecular Biology of the Cell 11(12): 4131-4142.
Segalés, J. y M. Domingo. 2003. La necropsia en el ganado porcino, diagnóstico anatomopatológico y toma de muestras. Boehringer Ingelheim, Madrid. 43 p.
Shen, L. and J.R. Turner. 2005. Actin depolymerization disrupts tight junctions via caveolae-mediated endocytosis. Molecular Biology of the Cell 16(9): 3919-3936.
Shifflett, D.E., D.R. Clayburgh, A. Koutsouris, J.R. Turner and G.A. Hecht. 2005. Enteropathogenic E. coli disrupts tight junction barrier function and structure in vivo. Laboratory Investigation; a Journal of Technical Methods and Pathology 85(10): 1308-1324.
Steel, R.G. and J.H. Torrie. 1980. Principles and procedures of statistics: a biometrical approach. Second edition. McGraw-Hill, New York. 633 p.
Tlaskalová-Hogenová, H., R. Stepánková, T. Hudcovic, L. Tucková, B. Cukrowska, R. Lodinová-Zádníková, H. Kozáková, P. Rossmann, J. Bártová, D. Sokol, D.P. Funda, D. Borovská, Z. Reháková, J. Sinkora, J. Hofman, P. Drastich and A. Kokesová. 2004. Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunology Letters 93(2-3): 97-108.
Tsukita, S. and M. Furuse. 2002. Claudin-based barrier in simple and stratified cellular sheets. Current Opinion in Cell Biology 14(5):531-536
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe and F. Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biology 3(7): Research 0034.
Waseem, T., M. Duxbury, H. Ito and M. Robinson. 2008. Exogenous ghrelin modulates release of pro- and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Surgery 143: 334-342.
Xu, S., J. Lee and M. Miyake. 2011. Expression of ZO-1 and occludin at mRNA and protein level during preimplantation development of the pig parthenogenetic diploids. Zygote 20(2): 147-158.
Yang, E.V., and R. Glaser. 2000. Stress-induced immunomodulation: Impact on immune defenses against infectious disease. Biomedicine and Pharmacotherapy 54: 245-250.
Yu, D., A.M. Marchiando, C.R. Weber, D.R. Raleigh, Y. Wang, L. Shen and J.R. Turner. 2010. MLCK-dependent exchange and actin binding region-dependent anchoring of ZO-1 regulate tight junction barrier function. Proceedings of the National Academy of Sciences of the United States of America 107(18): 8237-8241.
References
Amador, P., J. García, M.C. Marca, J. de la Osada, S. Acín, M.A. Navarro, M.T. Salvador, M.P. Lostao and M.J. Rodríguez-Yoldi. 2007. Intestinal D-galactose transport in an endotoxemia model in the rabbit. The Journal of Membrane Biology 215(2-3): 125-133.
Berkes, J., V.K. Viswanathan, S.D. Savkovic and G. Hecht. 2003. Intestinal epithelial responses to enteric pathogens: effects on tight junction barrier, ion transport, and inflammation. Gut 52(3): 439-451.
Bruewer, M., A. Luegering, T. Kucharzik, C.A. Parkos, J.L. Madara, A.M. Hopkins and A. Nusrat. 2003. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. Journal of Immunology 171(11):6164-72.
Bustin, S.A. 2000. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. Journal of Molecular Endocrinology 25(2): 169-193.
Chin, A.C., D.A. Teoh, K.G. Scott, J.B. Meddings, W.K. Macnaughton and A.G. Buret. 2002. Strain-dependent induction of enterocyte apoptosis by Giardia lamblia disrupts epithelial barrier function in a caspase-3-dependent manner. Infection and Immunity 70(7): 3673-3680.
Council for International Organizations of Medical Sciences. (CIOMS). 1985. International Guiding Principles for Biomedical Research Involving Animals. Geneva. 28 p.
Chow, J., S.M. Lee, Y. Shen, A. Khosravi and S.K. Mazmanian. 2010. Host-bacterial symbiosis in health and disease. Advances in Immunology 107: 243-274.
Davis, M.E., S.C. Sears, J.K. Apple, C.V. Maxwell and Z.B. Johnson. 2006. Effect of weaning age and commingling after the nursery phase of pigs in a wean-to-finish facility on growth, and humoral and behavioral indicators of well-being. Journal of Animal Science 84: 743-756.
Fan, M.Z. 2002. Growth and ontogeny of the gastrointestinal tract. pp. 31-60. In: Xu, R.J. and P. Cranwell (eds.). The neonatal pig: gastrointestinal physiology and nutrition. Nottingham University Press, Nottingham. 360 p.
Freeman, W.M., S.J. Walker and K.E. Vrana. 1999. Quantitative RT-PCR: pitfalls and potential. Biotechniques 26(1): 112-22.
García, J., B. Abad and M.J. Rodríguez-Yoldi. 2003. Effect of lipopolysaccharide on D-fructose transport across rabbit jejunum. Inflammation Research 52(4): 177-184.
García, J, M.C. Marca, E. Brot-Laroche, N. Guillén, S. Acin, M.A. Navarro, J. Osada and M.J. Rodríguez-Yoldi. 2008. Protein kinases, TNF-a, and proteasome contribute in the inhibition of fructose intestinal transport by sepsis in vivo. American Journal of Physiology. Gastrointestinal and Liver Physiology 294(1): G155-G164.
Hataya, Y., T. Akamizu, H. Hosoda, N. Kanamoto, K. Moriyama, K. Kangawa, K. Takaya and K. Nakao. 2003. Alterations of plasma ghrelin levels in rats with lipopolysaccharide-induced wasting syndrome and effects of ghrelin treatment on the syndrome. Endocrinology 144: 5365-5371.
Huggett, J., K. Dheda, S. Bustin and A. Zumla. 2005. Real-time RT-PCR normalisation; strategies and considerations. Genes and Immunity 6(4): 279-84.
Kojima, C.J., J.A. Carroll, R.L. Matteri, K.J. Touchette and G.L. Allee. 2007. Effects of weaning and weaning weight on neuroendocrine regulators of feed intake in pigs. Journal of Animal Science 85(9): 2133-2139.
Kubista, M., J.M. Andrade, M. Bengtsson, A. Forootan, J. Jonák, K. Lind, R, Sindelka, R. Sjöback, B. Sjögreen, L. Strömbom, A. Ståhlberg and N. Zoric. 2006. The real-time polymerase chain reaction. Molecular Aspects of Medicine 27(2-3): 95-125.
Lallès, J.P., G. Boudry, C. Favier, N. Le Floc'h, S. Pié, C. Piel and B. Sève, 2004. Gut function and dysfunction in young pigs: physiology. Animal Research 53: 301- 316.
Lamb-Rosteki, J., L. Kalischuk, G. Inglis and A. Buret. 2008. Epidermal growth factor inhibits Campylobacter jejuni-induced Claudin-4 disruption, loss of epithelial barrier funtion, and Escherichia coli translocation. Infection and Immunity 76(8): 3390-3398.
Makkink, C.A., P.J.M. Berntsen, B.M.L. op den Kamp, B. Kemp and W.A. Verstegen. 1994. Gastric protein breakdown and pancreatic enzyme activities in response to two different dietary protein sources in newly weaned pigs. Journal of Animal Science 72(11): 2843-2850.
Mariani, V., S. Palermo, S. Fiorentini, A. Lanubile and E. Giuffra. 2009. Gene expression study of two widely used pig intestinal epithelial cell lines: IPEC-J2 and IPI-2I. Veterinary Immunology and Immunopathology 131(3-4): 278-284.
Marion, J., M. Biernat, F. Thomas, G. Savary, Y. Le Breton, R. Zabielski, I. Le Huërou-Luron and J. Le Dividich, 2002. Small intestine growth and morphometry in piglets weaned at 7 days of age. Effects of level of energy intake. Reproduction, Nutrition, Development 42(4): 339-354.
McLaughlin, J., P. Padfield, J. Burt and C. O´Neil. 2004. Ochratoxin a increases permeability through tight juntions by removal of specific claudin isoforms. American Journal of Physiology. Cell physiology 287(5): C1412-C1417.
Muller, P.Y., H. Janovjak, A.R. Miserez and Z. Dobbie. 2002. Processing of gene expression data generated by quantitative real time RT-PCR. Biotechniques 32(6): 1372-1374, 1376, 1378-1379.
Musch, M.W., M.M. Walsh-Reitz and E.B. Chang. 2006. Roles of ZO-1, occludin, and actin in oxidant-induced barrier disruption. American Journal of Physiology and Gastrointestinal Liver Physiology 290: G222-G23.
National Research Council (NRC). 2012. The nutrient requirements of swine. Eleventh Revised Edition. The National Academy Press, Washington. 400 p.
Parra, S.J.E., T.J. Agudelo, P.D. Sanín, DJ. Forero, L.C. Muskus and H.A. López. 2013. Intestinal expression of pro-inðAammatory cytokines induced by oral intake of lipopolysaccharide (LPS) from E. coli in weaned pigs. Revista Colombiana de Ciencias Pecuarias 26: 108-118.
Paulin, S.M., A. Jagannathan, J. Campbell, T.S. Wallis and M.P. Stevens. 2007. Net replication of Salmonella enterica serovars Typhimurium and Choleraesuis in porcine intestinal mucosa and nodes is associated with their differential virulence. Infection and Immunity 75(8): 3950-3960.
Petersen, Y.M., D.G. Burrin and P.T. Sangild. 2001. GLP-2 has differential effects on small intestine growth and function in fetal and neonatal pigs. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology 281(6): 1986-1993.
Pié, S., S.P. Lallès, F. Blazy, J. Laffitte, B. Sève and I.P. Oswald. 2004. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. Journal of Nutrition 134: 641-647.
Pinton, P., J. Nougayréde, J. Del Rio, C. Moreno, D. Marin, L. Ferrier, A. Bracarence, M. Kolf-Clauw and I. Oswald. 2009. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicology and Applied Pharmacology 237(1): 41-48.
Pitman, R.S. and R.S. Blumberg. 2000. First line of defense: the role of the intestinal epithelium as an active component of the mucosal immune system. Journal of Gastroenterology 35(11): 805-814.
Pluske, J.R., D.E. Hopwood y D.J. Hampson. 2003. Relación entre la microbiótica intestinal, el pienso y la incidencia de diarreas, y su influencia sobre la salud del lechón tras el destete. pp. 94-108. En: XIX Curso de Especialización FEDNA, Madrid.
Reis, S.T.C., C.M.J. Guerrero, B.A. Aguilera y L.G. Mariscal. 2005. Efecto de diferentes cereales sobre la morfología intestinal de lechones recién destetados. Técnica Pecuaria en México 43(3): 309-321.
Reis, S.T.C., L.G. Mariscal, B.A. Aguilera y J.G.H. Cervantes. 2007. Digestibilidad de la proteína y energía en dietas para lechones complementadas con tres diferentes tipos de suero de leche deshidratado. Revista Veterinaria México 38(2): 141-151.
SASTM. 2006. SAS/STAT User's Guide. Institute Inc. Statistical Analysis Systems Institute. Version 9.1th edition. Cary, NC: SAS Institute Inc.
Saitou, M., M. Furuse, H. Sasaki, J.D. Schulzke, M. Fromm, H. Takano, T. Noda and S. Tsukita. 2000. Complex phenotype of mice lacking occludin, a component of tight junction strands. Molecular Biology of the Cell 11(12): 4131-4142.
Segalés, J. y M. Domingo. 2003. La necropsia en el ganado porcino, diagnóstico anatomopatológico y toma de muestras. Boehringer Ingelheim, Madrid. 43 p.
Shen, L. and J.R. Turner. 2005. Actin depolymerization disrupts tight junctions via caveolae-mediated endocytosis. Molecular Biology of the Cell 16(9): 3919-3936.
Shifflett, D.E., D.R. Clayburgh, A. Koutsouris, J.R. Turner and G.A. Hecht. 2005. Enteropathogenic E. coli disrupts tight junction barrier function and structure in vivo. Laboratory Investigation; a Journal of Technical Methods and Pathology 85(10): 1308-1324.
Steel, R.G. and J.H. Torrie. 1980. Principles and procedures of statistics: a biometrical approach. Second edition. McGraw-Hill, New York. 633 p.
Tlaskalová-Hogenová, H., R. Stepánková, T. Hudcovic, L. Tucková, B. Cukrowska, R. Lodinová-Zádníková, H. Kozáková, P. Rossmann, J. Bártová, D. Sokol, D.P. Funda, D. Borovská, Z. Reháková, J. Sinkora, J. Hofman, P. Drastich and A. Kokesová. 2004. Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunology Letters 93(2-3): 97-108.
Tsukita, S. and M. Furuse. 2002. Claudin-based barrier in simple and stratified cellular sheets. Current Opinion in Cell Biology 14(5):531-536
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe and F. Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biology 3(7): Research 0034.
Waseem, T., M. Duxbury, H. Ito and M. Robinson. 2008. Exogenous ghrelin modulates release of pro- and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Surgery 143: 334-342.
Xu, S., J. Lee and M. Miyake. 2011. Expression of ZO-1 and occludin at mRNA and protein level during preimplantation development of the pig parthenogenetic diploids. Zygote 20(2): 147-158.
Yang, E.V., and R. Glaser. 2000. Stress-induced immunomodulation: Impact on immune defenses against infectious disease. Biomedicine and Pharmacotherapy 54: 245-250.
Yu, D., A.M. Marchiando, C.R. Weber, D.R. Raleigh, Y. Wang, L. Shen and J.R. Turner. 2010. MLCK-dependent exchange and actin binding region-dependent anchoring of ZO-1 regulate tight junction barrier function. Proceedings of the National Academy of Sciences of the United States of America 107(18): 8237-8241.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
CrossRef Cited-by
1. F. Correa, D. Luise, L. Amatucci, F. Palumbo, S. Virdis, C. Negrini, P. Clavenzani, M. Vecchi, M. Mazzoni, P. Bosi, P. Trevisi. (2022). Effect of an Escherichia coli F4/F18 bivalent oral live vaccine on gut health and performance of healthy weaned pigs. animal, 16(11), p.100654. https://doi.org/10.1016/j.animal.2022.100654.
2. T. Skrzypek, S. Szymańczyk, K. Ferenc, W. Kazimierczak, K. Szczepaniak, R. Zabielski. (2018). The contribution of vacuolated foetal-type enterocytes in the process of maturation of the small intestine in piglets. Invited review. Journal of Animal and Feed Sciences, 27(3), p.187. https://doi.org/10.22358/jafs/94167/2018.
Dimensions
PlumX
- Citations
- CrossRef - Citation Indexes: 2
- Usage
- SciELO - Full Text Views: 1878
- SciELO - Abstract Views: 1637
- Captures
- Mendeley - Readers: 12
Article abstract page views
Downloads
License
Copyright (c) 2015 Revista Facultad Nacional de Agronomía Medellín

This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
The journal allows the author(s) to maintain the exploitation rights (copyright) of their articles without restrictions. The author(s) accept the distribution of their articles on the web and in paper support (25 copies per issue) under open access at local, regional, and international levels. The full paper will be included and disseminated through the Portal of Journals and Institutional Repository of the Universidad Nacional de Colombia, and in all the specialized databases that the journal considers pertinent for its indexation, to provide visibility and positioning to the article. All articles must comply with Colombian and international legislation, related to copyright.
Author Commitments
The author(s) undertake to assign the rights of printing and reprinting of the material published to the journal Revista Facultad Nacional de Agronomía Medellín. Any quotation of the articles published in the journal should be made given the respective credits to the journal and its content. In case content duplication of the journal or its partial or total publication in another language, there must be written permission of the Director.
Content Responsibility
The Faculty of Agricultural Sciences and the journal are not necessarily responsible or in solidarity with the concepts issued in the published articles, whose responsibility will be entirely the author or the authors.


















