



Publicado
ESTRUCTURA, RIQUEZA Y COMPOSICIÓN DE PLANTAS ARBORESENTES EN UN BOSQUE DE NIEBLA ENTRESACADO DEL TOLIMA (COLOMBIA)
Palabras clave:
Bosque de niebla, vegetación andina, estructura de la vegetación, extracción de maderas. (es)RESUMEN
Se presenta el estudio de la estructura, la riqueza y la composición florística del componente arborescente de un bosque entresacado, en la vereda Dantas, municipio de Ibagué, Tolima, Colombia, con base en el muestreo de los individuos con DAP ≥ 2,5 cm en 0,1 ha. El bosque se caracteriza por la escasez de lianas y de hemiepífitas, ausencia de familias típicas de bosques de niebla colombianos entre los 2000 y 2500 msnm (Ericaceae, Myrtaceae, Meliaceae y Aquifoliaceae) y por un incremento en la riqueza de familias como Sabiaceae y Euphorbiaceae. Comparado con otros bosques de niebla de los Andes colombianos y del neotrópico, éste presenta una de las más bajas densidades de tallos (237 individuos con DAP ≥ 2,5 cm por 0,1 ha) y una proporción de árboles grandes muy alta (39,7% individuos con DAP ≥ 10 cm por 0,1 ha).
Aparentemente, los efectos de la extracción selectiva de maderas sobre la riqueza, la estructura y la composición, fueron la disminución de la riqueza florística, del número de tallos del bosque, y de la densidad y la riqueza de lianas, así como un aumento en la densidad de especies secundarias como Hedyosmum goudotianum Slms-Laubach var. goudatianum, Miconia resima Naud. y Palicourea calophlebia Standl.
ESTRUCTURA, RIQUEZA Y COMPOSICIÓN DE PLANTASARBORESENTES EN UN BOSQUE DE NIEBLA ENTRESACADODEL TOLIMA (COLOMBIA).
Structure, Richness and Composition of Arboreal Plants in a Cloud Thinning Forest of Tolima (Colombia).
JUAN MANUEL CAMPO KURMEN1, Biólogo. 1 Universidad Nacional de Colombia, Sede Bogotá. Facultad de Ciencias, Departamento de Biología-Instituto de Ciencias Naturales. Bogotá, Colombia. alburacolombia@yahoo.es
Presentado 14 de octubre de 2010, aceptado 4 de junio de 2010, correcciones 18 de agosto de 2010.
RESUMEN
Se presenta el estudio de la estructura, la riqueza y la composición florística del componente arborescente de un bosque entresacado, en la vereda Dantas, municipio de Ibagué, Tolima, Colombia, con base en el muestreo de los individuos con DAP ≥2,5 cm en 0,1 ha. El bosque se caracteriza por la escasez de lianas y de hemiepífitas, ausencia de familias típicas de bosques de niebla colombianos entre los 2.000 y 2.500 msnm (Ericaceae, Myrtaceae, Meliaceae y Aquifoliaceae) y por un incremento en la riqueza de familias como Sabiaceae y Euphorbiaceae. Comparado con otros bosques de niebla de los Andes colombianos y del neotrópico, éste presenta una de las más bajas densidades de tallos (237 individuos con DAP ≥2,5 cm por 0,1 ha) y una proporción de árboles grandes muy alta (39,7% individuos con DAP ≥ 10 cm por 0,1 ha). Aparentemente, los efectos de la extracción selectiva de maderas sobre la riqueza, la estructura y la composición, fueron la disminución de la riqueza florística, del número de tallos del bosque, y de la densidad y la riqueza de lianas, así como un aumento en la densidad de especies secundarias como Hedyosmum goudotianum Slms-Laubach var. goudatianum, Miconia resima Naud. y Palicourea calophlebia Standl.
Palabras clave: bosque de niebla, vegetación andina, estructura de la vegetación, extrac-ción de maderas.
ABSTRACT
Structure, richness, and floristic composition of the woody elements of the selective logging forest of the Vereda Dantas, (Ibagué, Tolima, Colombia), where studied in a 0.1 ha plot sampled for all individuals ≥2.5 cm dbh. The forest is characterized by scarcity of lianas and hemiepiphytic, absence of typical families of the Colombian cloud forests between 2000 and 2500 m (Araceae, Ericaceae, Myrtaceae, Meliaceae and Aquifoliaceae), and richness increment of the Sabiaceae and Euphorbiaceae. Compared to others cloud forest from the Colombian Andes and the Neotropic, it has, fewer individuals (237 individuals ≥2,5 cm dbh per 0.1 ha) and more large trees (39,7% of individuals ≥10 cm dbh per 0,1 ha). The forest has a lower woody species richness (75 species ≥2,5 cm dbh per 0,1 ha). Apparently, the effects of -selective- timber extraction on structure, richness, and floristic composition are decrease floristic richness and density of individuals, decrease of lianas density and richness, and more individuals of secondary species, like: Hedyosmum goudotianum Slms-Laubach var. goudatianum, Miconia resima Naud. y Palicourea calophlebia Standl.
Key words: Cloud forest, Andes vegetation, vegetation structure, selective logging.
INTRODUCCIÓN
Los bosques neotropicales montanos están entre los menos conocidos y más amenazados de todas las formaciones vegetales tropicales (Van der Hammen y Cleef, 1983; Stadmuller, 1987; Gentry, 1993; Gentry, 2001). Sin embargo, los bosques de niebla tropicales han sido reconocidos como uno de los principales centros de especiación del mundo y una de las regiones más diversas del planeta (Gentry, 1982; Churchill et al., 1995). Henderson et al. (Henderson et al., 1991) estiman que en Colombia la cobertura remanente de los bosques andinos originales es de menos del 10%, mientras que Carrizosa-U, 1990, estima que queda menos del 5%. Muchos de estos bosques que hoy se conservan han sufrido explotaciones selectivas de maderas preciosas, principalmente de lauráceas, familia que Gentry, 1995, ha considerado como la más rica en especies en casi todos los bosques andinos entre 1.500 y 2.900 m de elevación; esta sobreexplotación ha traído como consecuencia que las lauráceas y otras especies maderables sean escasas o localmente extintas en muchos de los relictos de los bosques andinos colombianos.
El efecto de la tala selectiva en el bosque andino hasta el punto de casi desaparecer una familia entera, no tiene equivalente en los disturbios que ocurren naturalmente en la dinámica de estos bosques. Las implicaciones de este cambio son difíciles de elucidar, pero es evidente que la extracción selectiva de estas maderas no solo amenaza la conservación de estas especies sino que alterará la dinámica del bosque. El estudio de estos bosques entresacados, en lo que respecta a la modificación de su estructura, su riqueza y su composición ha sido pobre pese a su importancia. La presente investigación pretende estudiar estos cambios, haciendo un análisis comparativo con algunas investigaciones que se han realizado en bosques andinos primarios, desde el punto de vista de los cambios en la estructura, la riqueza y la composición florística del componente leñoso, en una muestra de 0,1 ha.
MÉTODOS
ÁREA DE ESTUDIO
La zona de estudio se encuentra ubicada en la vertiente oriental de la Cordillera Central de los Andes colombianos, en el departamento del Tolima, municipio de Ibagué, Vereda Dantas en la finca -La Barquereña-, cuyo rango altitudinal es de 2.260-2.500 msnm para un altura media de 2.340 msnm (Mediciones realizadas con un altímetro barómetro Konus®). El promedio de precipitación anual es de 1.635 mm y a lo largo del año se presentan dos picos de precipitación: de marzo a junio y de septiembre a octubre, mientras que el periodo menos lluvioso es de diciembre a enero (Promedio de 17 años, IDEAM, 2005). La pendiente promedio de los diez transectos es de 27,5º (Mediciones realizadas con clinómetro Sunnto®).
La zona ha sido sometida a diferentes grados de tala selectiva, de acuerdo con la dificultad para transportar la madera. Se realizaron entrevistas no estructuradas con cinco campesinos y se estableció con confiabilidad que la tala fue efectuada en dos periodos, hace 30 (la más intensa) y hace nueve años, y se extrajeron solo las especies de mayor valor en el mercado; Laureles (Lauraceae), Pino Romerón (aparentemente Prumnopytis sp.), Cedros (Cedrela sp.) y Nogales (Juglans neotropica). A pesar de la tala selectiva que sufrieron los bosques de la zona en estudio, la región constituye un importante relicto de los bosques andinos de la vertiente oriental de la Cordillera Central.
MÉTODO DE MUESTREO
Se establecieron diez transectos en bosques primarios que han sufrido tala selectiva, de 50 x 2 m (0,1 ha) siguiendo la metodología del inventario rápido (Gentry, 1995). Los sitios para los transectos fueron escogidos evitando la presencia de grandes claros y áreas de perturbación evidente como potreros, cultivos abandonados o caminos y tratando de incluir el rango de variación topográfica de la zona. Se contaron y se recolectaron todos los individuos con un diámetro igual o mayor a 2,5 cm a la altura del pecho (1,3 m por encima del suelo) y se registraron datos de altura de la primera ramificación, altura total, DAP, presencia de exudados, características de la corteza, número de tallos por debajo de 1,3 m (en el caso de hallar tallos múltiples estos se midieron separadamente) y forma de crecimiento. Los individuos ubicados en los límites del transecto, solo se contaron si más de la mitad de su tronco se encontraba dentro de la parcela. Adicionalmente se midió la pendiente promedio de cada transecto y su altura sobre el nivel del mar. Para la delimitación de las familias se siguió el sistema de clasificación de Cronquist, 1981; la determinación de las especies se realizó en el Herbario Nacional Colombiano (COL), (Universidad Nacional de Colombia, sede Bogotá), donde fueron depositados los especimenes bajo la serie de colección del autor.
ANÁLISIS DE DATOS
El área basal se calculó siguiendo a Caín y Castro, 1959. Se calculó el índice de valor de importancia de cada especie (IVI) como la sumatoria de la densidad (DeR), la frecuencia (FR) y la dominancia relativa (DoR). Se calculó el índice de valor de importancia para familias (VIF) para evaluar la importancia ecológica éstas, como la sumatoria de la densidad, la dominancia y la riqueza relativas de cada familia, según lo propuesto por Mueller-Dombois y Ellenberg (Mueller-Dombois y Ellenberg, 1974) en donde DeR = (# de individuos por especie/# total de individuos en la comunidad) x 100; FR = (# de subtransectos en los que aparece la especie/∑de las frecuencias de todas las especies) x 100; DoR = (∑Área basal de todos los individuos de la especie/∑Área basal de toda la comunidad) x 100. Se estimó el número de especies esperadas para los sitios utilizando la ecuación de regresión de Gentry, 1995, para los bosques andinos: # de especies = (260,1-0,073 x altitud) y los índices de diversidad de Shannon-Wiener, según la ecuación: H’= -∑pi ln pi ; en donde pi es la proporción de individuos encontrada en cada una de las especies (Magurran, 1988) y el índice de Simpson calculado de la ecuación: D = ∑(ni (ni-1)/N(N-1)); en donde ni=número de individuos en cada especie y N = el total de individuos muestreados (Magurran, 1988).
RESULTADOS
Se encontraron 237 individuos, 75 especies, 58 géneros y 34 familias. En promedio se presentaron 2,1 especies por familia, 1,4 especies por género y 3,2 individuos por especie, en las tablas Tabla. 1 y Tabla. 2 se presenta la distribución de las especies y los individuos por hábito.
ESTRUCTURA
La cantidad de tallos fue inferior en comparación a la reportada para otros bosques andinos colombianos de altitud similar (Tabla. 2). La estructura vertical del bosque estuvo caracterizada en general por un dosel bastante elevado, 39 individuos superaron los 20 m, 18 superaron los 25 m y diez los 30 m; el promedio en altura fue de 12 m para todos los individuos y de 14 m para los árboles en particular. Por otra parte, se encontró que el 46% del área basal se concentra en las diez especies más comunes, mientras que si se toman las diez especies más importantes (con base en el IVI) el área basal repre-senta el 64% del total. En cuanto al porte de los individuos, el 39,2 % de los individuos alcanzó un DAP≥10 cm representando 33 especies de 22 familias y el 92% del área basal, mientras el 2,5% de los individuos registraron un DAP>40 cm representado seis individuos de seis especies y cinco familias.
La distribución de individuos, especies y familias por clases diamétricas mostró una curva en forma de J invertida. Por el contrario, la distribución del área basal manifestó patrones irregulares y tuvo su mayor expresión en las dos últimas clases diamétricas (Fig. 1).
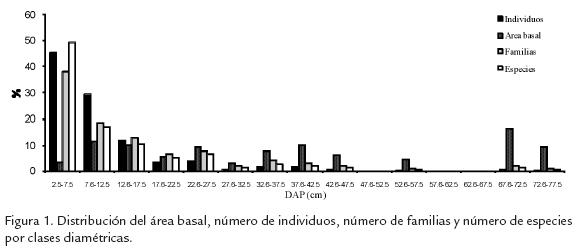
0,84% de los individuos, aunque en términos de área basal representaron lo mismo que las lianas (0,17%). Las hemiepífitas estuvieron ausentes en toda la muestra.
RIQUEZA Y COMPOSICIÓN
El número de especies esperadas para los sitios según la ecuación de regresión de Gentry, 1995, para el bosque en estudio fue de 89, aunque este bosque presentó catorce especies menos de las esperadas.
Las cinco especies más abundantes fueron Cyathea sp., Miconia cf. orescia L. Uribe, Palicourea calophlebia Standl., Weinmannia pubescens Kunth y Hedyosmum goudotianum Slms-Laubach var. goudatianum. Estas especies a pesar de ser el 7% de las especies totales, representan el 41% de todos los individuos y el 19% del área basal.
Al comparar la composición a nivel de familia con la de tres bosques de la vertiente oriental de la cordillera central de los Andes colombianos entre los 2.000 y 2.500 m, familias comunes en estos bosques, como Araceae, Aquifoliaceae, Meliaceae y Ericaceae no se presentaron en La Barquereña y otras como Euphorbiaceae y Sabiaceae superaron el promedio notoriamente, a su vez se encontraron muy pocas especies de lauráceas (Fig. 2). En el anexo 1, se presenta un listado detallado de las especies.

Valor de importancia de familia (VIF). Las cinco familias más importantes fueron Melastomataceae, Rubiaceae, Cyatheaceae, Theaceae y Moraceae con el 51% de los individuos y el 58% del área basal. Si se toman las diez familias más importantes éstas representan el 76% de los individuos y el 80% del área basal total. La familia Lauraceae ocupó el puesto 16 en importancia siendo incluso superada por familias como Sabiaceae y Piperaceae. Al considerar el VIF de los individuos con un DAP≥2,5 cm en comparación al VIF de los individuos con un DAP≥10 cm, familias típicas de bosques maduros como Sabiaceae e Hippocastanaceae, incrementan su importancia notoriamente. Las familias Rubiaceae y Myrsinaceae por el contrario, disminuyen su importancia. Algunas familias dejan de estar entre las 20 más importantes como Piperaceae, Monimiaceae y Apocynaceae, siendo remplazadas en importancia por Juglandaceae, Arecaceae y Sapindaceae.
Iacute;ndice de valor de importancia (IVI). Las cinco especies más importantes fueron Cyathea sp., Frezieracanescens Bonpl. Miconia cf. orescia, Tibouchinalepidota y Weinmanniapubescens, representando el 35% de los individuos y el 42% del área basal total. Si se toman las diez especies más importantes, éstas representan el 50% de los individuos y el 64% del área basal.
DIVERSIDAD
El resultado obtenido para el índice de diversidad de Shannon-Wiener fue de 3,7 y para el de Simpson fue de 21,7.
DISCUSIÓN
ESTRUCTURA
El bosque presenta varias particularidades estructurales, que lo destacan entre muchos de los bosques de niebla colombianos y neotropicales. El número de tallos encontrado es particularmente bajo, incluso es más bajo que el de las 19 localidades en Colombia, nueve en Perú, tres en Ecuador y tres en Bolivia reportadas por Gentry, 1995, para bosques de niebla con muestras de 0,1 ha con un DAP≥2,5 cm. Esto puede estar relacionado con la densidad registrada de árboles grandes (39,2 % de los individuos con más de 10 cm de DAP), 16,4% más que el promedio para seis bosques nublados de alturas y pluviosidad similares; mientras que el promedio de individuos con DAP ≥2,5≤10 cm fue la mitad que el promedio para lo bosques comparados, la densidad de individuos con DAP≥10 cm fue prácticamente la misma (véase Tabla. 2).
Teniendo en cuenta únicamente los individuos con un DAP>10 cm, este bosque presentó un promedio de 20 cm de DAP, mientras que en estudios realizados en siete bosques primarios ecuatorianos entre los 1.825 m y los 2.425 m (Bussmann et al., 2005), se obtuvo un promedio de 14,2 cm con un pico máximo de 15,4 cm. La altura del dosel fue también característica, dado que presentó 23 m de altura y árboles emergentes hasta de 33 m, mientras que según estudios de Rangel y Garzón, 1994, solo en tres de cuatro levantamientos en bosques de la Cordillera Central entre los 2.140 m y los 2.480 msnm se presentaron árboles por encima de los 25 m. A nivel de la altura total de los individuos de DAP≥10 cm se encontró un promedio13,2 m mientras que en el estudio de los bosques ecuatorianos citados, el promedio más elevado (9,4 m) y el área basal prácticamente duplica la de cualquiera de estos bosques. Además, comparando el área basal de los individuos con un DAP≥10 cm con el promedio de siete bosques nublados entre los 2.000 y 2.500 m del Perú (Antón y Reynel, 2004), el área basal de La Barquereña la duplica a pesar de que solo posee dos terceras partes de los individuos que poseen dichos bosques. En resumen el bosque se caracteriza por un dosel alto y una gran cantidad de árboles robustos con la correspondiente reducción del número de tallos de los estratos inferiores por competencia lumínica.
RIQUEZA Y COMPOSICIÓN
La riqueza de especies en el bosque de La Barquereña fue un 16% más baja que la esperada según la ecuación de regresión de Gentry (Gentry, 1995), además, debe tenerse en cuenta que para los bosques maduros de la vertiente oriental de la Cordillera Central, la diversidad supera a la esperada, de hecho, en los bosques de cuatro localidades de la Cordillera Central a alturas similares y con muestras de 0,1 ha la riqueza era en promedio 17% superior a la esperada (Tabla. 2); asimismo, la Cordillera Central, específicamente en el PNN Los Nevados, ha sido considerada como uno de los centros de concentración de especies más importantes en Colombia (Rangel, 1995). Por todo lo anterior, podría pensarse que la riqueza encontrada puede ser incluso más baja de lo que revela la ecuación de regresión según Gentry, 1995.
Para los bosques en general se ha encontrado que la riqueza de especies está asociada con factores físicos y bióticos, dentro de los cuales cabe destacar la correlación positiva entre riqueza y pluviosidad (Gentry, 1995), como también entre numero de tallos y riqueza (Lieberman et al., 1995; Denslow, 1995; Condit et al., 1996); así como una correlación negativa entre riqueza y aumento del DAP promedio (Whitmore, 1975). En el caso del bosque de La Barquereña, estos tres factores podrían estar cumpliendo un papel determinante en la disminución de la riqueza de especies, por presentar características particulares, no encontradas en otros bosques andinos entre los 2.000 y 3.000 msnm en Colombia; estas particularidades son: la precipitación promedio anual para la zona es 31% inferior a la registrada en otros estudios, una baja densidad de tallos (46% por debajo del promedio) y un alto porcentaje de árboles con DAP>10 cm (39,2%), mientras que el promedio para los otros bosqueses de 28% y ninguno de ellos sobrepasa el30% .Adicionalmente se conoce que la extracción maderera ocurrida hace 30 años fue dirigida especialmente a la extracción de amarillos (Lauraceas); siendo lauraceae la familia con mayor número de especies para este tipo de bosques (Fig. 2), es esperable una repercusión directa en la disminución de la riqueza de los bosques de la Barquereña. De esta manera la baja riqueza se explica principalmente por un bajo promedio de precipitación anual, bajo densidad de tallos y un alto porcentaje de individuos de gran porte, así como por la extracción selectiva de amarillos. Por otra parte, no se puede desconocer que el factor edáfico también puede influir de manera relevante en dicha riqueza, pero no se tienen análisis de suelos para la zona de estudio, que nos permitan dilucidar el comportamiento del bosque con respecto a este factor.
Otra particularidad florística del bosque de la Barquereña es que la riqueza de la familia Sabiaceae es más alta en comparación con la de otros bosques similares (Fig. 2); es inusual encontrar más de una especie de Meliosma en una muestra de estas dimensiones (Gentry, 1993). Las especies de Meliosma son árboles de madera dura, lento crecimiento y características de bosques maduros, por lo que puede estar presentándose un remplazamiento ecológico de las lauráceas que fueron extraídas. Lo mismo podría estar ocurriendo con la familia Hippocastanaceae que está entre las 20 familias más importantes, en contraste con otros bosques.
Valor de importancia de familia (IVF). Las familias Melastomataceae, Rubiaceae, Cyatheaceae y Theaceae son las más importantes del bosque en estudio, las tres primeras reflejan un aumento en las especies heliófitas después de la entresaca (Fig. 1), mientras que Theaceae representada por una sola especie: Freziera aff. canescens Bonpl. registra el segundo IVI más alto con 10 individuos, esto explica el elevado IVF de esta familia dada la tendencia de esta especie por formar bosques monoespecíficos.
Contrario a lo esperado y acorde con la extracción maderera de la zona, las lauráceas ocupan el puesto 16 a nivel de IVF. La familia Chloranthaceae también obtuvo un elevado IVF que está relacionada con la abundancia de Hedyosmum goudotianum, un importante componente de la Cordillera Central, siendo Hediosmo-Nectandrion la principal alianza para los bosques subandinos de la Cordillera Central (Rangel, 1995); por otro lado está abundancia de especies del genero Hedyosmun está relacionada con un pasado de disturbios (Gentry, 1993).
Teniendo en cuenta la alta representatividad de los individuos con DAP≥10 cm, se calculó el IVF separadamente con estos individuos, encontrando que familias típicas de bosques maduros como Sabiaceae e Hippocastanaceae incrementan su importancia notoriamente en relación al IVF general (DAP≥2,5 cm.), las familias Rubiaceae y Myrsinaceae por el contrario, disminuyen su importancia dado que en general las especies encontradas son arbolitos de sotobosque o arbustos; familias como Piperaceae, Monimiaceae y Apocynaceae dejan de aparecer dentro de las 20 más importantes, debido a que las especies encontradas son de tipo arbustivo o arbolitos y son remplazadas en importancia por Juglandaceae, Arecaceae y Sapindaceae; La familia Arecaceae aparece dentro de las 20 más importantes al considerar solo los individuos con un DAP≥10cm, pese a que si bien esta familia estuvo pobremente representada por una sola especie, dicha familia era bastante común. Además, debe mencionarse que durante la fase de campo se observaron por lo menos seis especies de palmas en la zona de estudio, aunque solo una se presentó en los transectos.
DIVERSIDAD
La diversidad medida con el índice de Shannon-Wiener fue mayor en el presente estudio que la encontrada por Kappelle, 2001; en la cordillera de Talamanca en Costa Rica en un bosque de niebla maduro de altura máxima de dosel de 35 m y a 2.300 msnm (3,7 y 3,48 respectivamente). Además, cuando se midió la diversidad con el índice de Simpson, esta fue mayor en La Barquereña que en Costa Rica (21,7 y 8, respectivamente), sugiriendo que las especies están distribuidas más equitativamente en cuanto a densidad en el bosque de La Barquereña.
A pesar de que tal equitatividad de especies parece decrecer por efecto de la tala selectiva (por lo menos en tierras bajas, véase Yockteng y Cavalier, 1998), se requieren de más datos sobre el índice de Simpson de otros bosques de niebla similares, dado que la dominancia de unas pocas especies es una característica frecuente en bosques primarios de tierras altas, posiblemente como consecuencia de la influencia de componentes florísticos holárticos y australes como Fagaceae, Betulaceae, Theaceae, Cunoniaceae, y los consiguientes bosques dominados por estas especies: Robledales, Alisales, Cerezales y Bosques de encenillo, respectivamente.
ECOLOGÍA DE LAS ESPECIES
Con base en la información reportada por Vargas y Palacios (Vargas, 2004; Palacios, 2004) para 63 de las especies de las 75 especies encontradas, se encontró que nueve de las especies son exclusivas de bosques maduros, 32 de bosques secundarios y 22 se encuentran en ambos tipos de bosques (Fig. 3), esta clasificación está relacionada principalmente con la tolerancia lumínica de las especies en los estadios iníciales de desarrollo y establecimiento; aunque este tipo de clasificación se presta para muchas discusiones (véase Asquith 2002), es útil para entender los cambios en la dinámica del bosque.
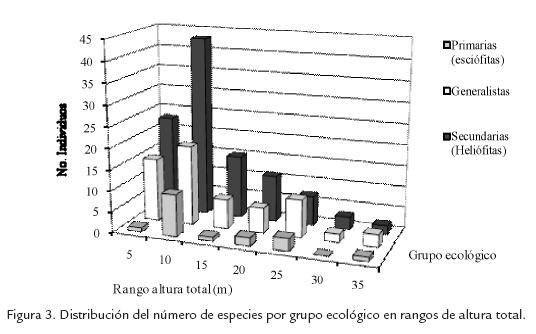
Podría pensarse entonces, que el bosque sufrió una importante disminución del numero de especies tolerantes a la sombra o especies climáxicas, pero parece haber poca claridad sobre el porcentaje esperado de estas especies en un bosque primario, teniendo en cuenta que según Hartshorm, 1980, más de las dos terceras partes de las especies encontradas en un bosque primario neotropical de tierras bajas resultaron ser intolerantes a la sombra (esciófitas), mientras que en estudios más recientes en Barro Colorado y La Selva se encontró que el 85% y 90% de las especies eran generalistas, en cada estación (Welden et al., 1991 y Lieberman et al., 1995). Independientemente de esto, las plantas esciófilas encontradas que representan el 14% del total de las especies, solo representan el 8% y el 9% de los individuos y del área basal respectivamente, haciendo que su importancia en densidad y dominancia sea mucho menor que en riqueza; más aún si se tiene en cuenta que el dosel está dominado por heliófitas y generalistas. Adicionalmente, las especies típicamente heliófilas se encontró que éstas representaban el 50% de las especies y el 60 % de los individuos, lo que revela que sí existe un marcado aumento de las especies heliófilas como consecuencia de la extracción maderera, coincidiendo con las observaciones de Wagner, 1997; quien además reportó que este cambio, se da incluso con cantidades de extracción relativamente bajas.
CONSIDERACIONES SOBRE EL EFECTO DE LA TALA SELECTIVA
Después de 30 años de haber ocurrido la extracción maderera, se encontró que las especies heliófilas y generalistas de mayor porte han cerrado el dosel y el estrato medio (el rango de altura total 5-10 m, presenta el 30% de los individuos (Fig. 3) de forma que la escasez de luz del sotobosque puede estar reduciendo las especies secundarias de corta vida, de hecho las especies típicamente secundarias encontradas en un bosque nublado de la serranía de Mérida (Schneider, 2003), la tercera parte era de tipo arbustiva, mientras en la Barquereña solo una octava parte de éste tipo de especies eran arbustos; por consiguiente, y aunque la disminución en la densidad de tallos está claramente relacionada con el aumento en área basal en relación al número de individuos, la tala selectiva puede estar implicada en la disminución de la densidad de tallos y está evidentemente afectando la riqueza total.
Algunas especies esciófitas como Hyeronima huilensis Cuatr. y Billia rosea Planch. y Linden ex Triana y Planch parecen iniciar una sucesión en aras de remplazar estructuralmente a las localmente desaparecidas lauráceas, pero dado su lento crecimiento no han alcanzado representatividad en el dosel (Fig. 3).
Desde el punto de vista de la escasez de lianas encontrada, muchas debieron ser arrancadas durante el proceso de extracción maderera y dada la longevidad de muchas de sus especies (Romero-Saltos, 1999) su recuperación es muy lenta, además como consecuencia de que las lianas reproductivamente maduras generalmente están asociadas a los árboles de mayor porte, la oferta de semillas debió ser fuertemente disminuida. Por otro lado, la alta densidad de helechos arborescentes asociada a la extracción maderera, disminuye la posibilidad de que las lianas trepen hasta el dosel como consecuencia de sus frondas caducifolias (Putz, 1980; Page y Brownsey, 1986), esto sumado a una baja densidad de tallos puede ser determinante en la escasez de lianas, puesto que la disponibilidad de soportes es quizás uno de los factores más determinantes en la distribución y abundancia de las lianas en un bosque (Putz, 1984a; Hara, 1988).
En resumen parece ser que la extracción maderera efectuada hace 30 años haya traído como consecuencia disminución de la riqueza de especies, del número de tallos del bosque, y de la densidad y la riqueza de lianas así como un aumento en la densidad de especies típicamente secundarias de mediana y larga vida, así como ocurre un remplazamiento ecológico y estructural de las lauráceas como principales elementos florísticos y estructurales del clímax de este tipo de bosques.
RECOMENDACIONES
Se recomienda realizar investigaciones puntuales sobre la historia de vida de las especies con énfasis en las que dominan el dosel de los bosques entresacados y que remplazan estructuralmente a las especies climáxicas, en aras de predecir las consecuencias de la extracción selectiva de maderas en el largo plazo. Se recomienda acompañar de cartografía espacial los estudios florísticos de este tipo de bosques dado que por sus características funcionales y estructurales estos bosques suelen tomarse como bosques primarios a pesar de los importantes cambios florísticos que se dan al interior de la comunidad después de la extracción de maderas; así como se recomienda hacer una investigación similar en el municipio de Dantas en un bosque lo suficientemente conservado para realizar comparaciones puntuales en los cambios producidos por esta extracción.
AGRADECIMIENTOS
A la Universidad Nacional de Colombia, sede Bogotá, especialmente al Departamento de Biología y al Instituto de Ciencias Naturales, por el apoyo brindado en la fase de laboratorio. A Idea Wild por la financiación de los equipos de campo. A Mabel Suescún, por su colaboración, durante la fase de campo y de laboratorio. Al Herbario Nacional Colombiano y a todos los botánicos que colaboraron en el proceso de determinación de los especímenes (Z. Cordero, D. Giraldo-Cañas, J. L. Fernández, G. Galeano, J. Betancurt, J.C. Granados, C. I. Orozco, J. Murillo y O. Rivera). Por ultimo, D. Giraldo-Cañas y G. Galeano por su asesoría académica y orientación durante el desarrollo de este estudio. A todos los campesinos que colaboraron con información acerca de la zona y su historia de uso, especialmente a Dider Cediel y Nubia Cediel.
BIBLIOGRAFÍA
ANDRADE GI. Paisaje y biodiversidad en las selvas de los Andes. En: G. I. Andrade (ed.), Carpanta, Ecología y conservación de un ecosistema altoandino. Bogotá: Editorial Presencia; 1993: p. 31-48.
ANTÓN D, REYNEL (eds.). Relictos de bosques de excepcional diversidad en los Andes Centrales del Perú. Lima: Universidad Nacional Agraria La Molina; 2004.
ASQUITH NM. La dinámica del bosque y la diversidad arbórea. En: Guariguata MR, Kattan GH, eds. Ecología y conservación de bosques neotropicales. Cartago: Ediciones LUR; 2002. p. 377-406
BUSSMAN RW, GÁLVEZ JR, ORDOÑEZ OR. Forest structure in an undisturbed tropical mountain forest in southern Ecuador. Descargado el 2005 de: http://www.groovybits.com
CARRIZOSA-U J. La selva andina. Selva y Futuro. En: Carrizosa-U J, Hernández-C J, eds. Selva y Futuro. Bogotá: El Sello Editorial; 1990. p. 151-184.
CHURCHILL SP, BALSLEV H, FORERO E, LUTEYN JL, eds. Biodiversity and Conservation of Neotropical Montane Forests. Nueva York: The New York Botanical Garden; 1995.
CRONQUIST A. An integrated System of Classification of Flowering Plants. Nueva York: Columbia University Press; 1981.
GENTRY AH. Neotropical floristic diversity: phytogeographical connections between Central and South America, Pleistoscene climatic fluctations, or an accident of the Andean orogeny? Ann Mo Bot Gard. 1982;69:557-593.
GENTRY AH. Vistazo general a los bosques andinos y a la flora de Carpanta. En: Andrade GI, ed. Paisaje y biodiversidad en las selvas de los Andes. Carpanta, Ecología y conservación de un ecosistema Altoandino. Bogotá: Editorial Presencia; 1993. p. 67-79
GENTRY AH. Patterns of diversity and floristic composition in Neotropical montane forest. Biodiversity and conservation of Neotropical montane forest. En: Churcill SP, Balslev H, Forero E, Luteyn JL, eds. Biodiversity and Conservation of Neotropical Montane Forests. Nueva York: The New York Botanical Garden; 1995. p. 85-123.
GENTRY AH, EMONNS LH. Geographical variation in fertility, phenology, and composition of the understory of Neotropical forests. Biotropica. 1987;19(3):216-227.
HARTSHORN GS. Neotropical forest dynamics. Biotropica. 1980;12(suppl.):23-30.
HENDERSON AS, CHURCHILL L, LUTEYN L. Neotropical plant diversity. Nature. 1991;351:21-22.
IDEAM. Reporte de valores totales mensuales de precipitación, estación pluviométrica 2121017. Promedio desde 1987 al 2003. Tolima, municipio de Ibagué, corriente de Cocora, Hacienda Palogrande. Ibagué: IDEAM; 2005
KAPPELLE M. Costa Rica. En: Kappelle M, Brown AD, eds. Bosques nublados del neotrópico. Santo Domingo de Heredia, Costa Rica: Editorial Inbio; 2001. p. 301-370.
MAGURRAN A.E. Ecological diversity and its measurement. Princeton, Nueva Jersy: Princeton University Press; 1988.
MUELLER DOMBOIS D.ELLENBERG H. Aims and methods of vegetation ecology. Nueva York: John Wiley & Son; 1974.
PALACIOS WA. Forest species communities in tropical rain forest of Ecuador. Lyonia. 2004;7(1):33-40.
RANGEL O, ed. Colombia Diversidad Biótica. I. Instituto de Ciencias Naturales. Convenio Inderena-Universidad Nacional de Colombia. Bogotá: Ed. Guadalupe Ltda; 1995.
RANGEL O, GARZON A, GARZÓN C. Aspectos de la estructura de la diversidad y de la dinámica de la vegetación del parque regional natural Ucumarí. En: O. Rangel, ed. Ucumarí: un caso típico de la diversidad biótica andina. Bogotá: Corporación Autónoma Regional de Risaralda. Universidad Nacional de Colombia. Instituto de Ciencias Naturales. 1994. p. 85-108.
ROMERO-SALTOS HG. Diversidad, análisis estructural y aspectos florísticos relevantes de las lianas en una parcela de bosque muy húmedo premontano, Amazonía Ecuatoriana (Trabajo de grado). Quito: Departamento de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador; 1999. p. 103.
SCHNEIDER JVJ, GAVIRIA G, ZIZKA G. Inventario florístico de un bosque altimontano húmedo en el Valle de San Javier, Edo. Mérida, Venezuela. Plantula. 2003;3(2):65-81.
VAN DER HAMMEN A, CLEEF A. Datos para la historia de la flora andina. Revista Chilena de Hist Nal. 1983;56:97-107.
VARGAS W G. Guía ilustrada de las plantas de las montañas del Quindío y los Andes Centrales. Manizales: Editorial Universidad de Caldas; 2002.
WAGNER U. Efectos del manejo maderero sobre la biodiversidad florística y la estructura de bosques húmedos de Costa Rica. En: Memoria del Simposio Internacional sobre posibilidades de Manejo Sostenible en América Tropical. Santa Cruz de la Sierra, Bolivia: BOLFOR, IUFRO, CIFOR; 1997. p. 50-58.
YOCKTENG R, CAVELIER J. Diversidad y mecanismos de dispersión de árboles de la Isla Gorgona y de los bosques húmedos tropicales del Pacífico colombo-ecuatoriano. Rev Biol Trop. 1998;46(1):45-53.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2010 Acta Biológica Colombiana

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
1. La aceptación de manuscritos por parte de la revista implicará, además de su edición electrónica de acceso abierto bajo licencia Attribution-NonCommercial-ShareAlike 4.0 (CC BY NC SA), la inclusión y difusión del texto completo a través del repositorio institucional de la Universidad Nacional de Colombia y en todas aquellas bases de datos especializadas que el editor considere adecuadas para su indización con miras a incrementar la visibilidad de la revista.
2. Acta Biológica Colombiana permite a los autores archivar, descargar y compartir, la versión final publicada, así como las versiones pre-print y post-print incluyendo un encabezado con la referencia bibliográfica del articulo publicado.
3. Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
4. Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos institucionales, en su página web o en redes sociales cientificas como Academia, Researchgate; Mendelay) lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).












