



Publicado
Estructura, diversidad y dinámica de la vegetación en el ecotono bosque-páramo: revisión de la evidencia en la Cordillera de Mérida
Vegetation Structure, Diversity and Dynamics in the Forest-Paramo Ecotone: A Review of the Evidence in the Cordillera de Mérida
DOI:
https://doi.org/10.15446/abc.v20n3.46721Palabras clave:
Andes tropicales, cambio climático, disturbio agrícola, formas de vida, gradiente altitudinal, sucesión secundaria (es)agricultural disturbance, climate change, elevation gradient, life forms, secondary succession, tropical Andes (en)
En los Andes tropicales, el límite altitudinal de los bosques corresponde a una compleja zona de transición entre el bosque paramero y el páramo andino. En el límite inferior de esta zona ocurre a su vez una transición gradual entre el bosque y las selvas nubladas. La ordenación del territorio en estos ecosistemas se ha convertido en un tema prioritario en virtud de la creciente amenaza que representan para su conservación las actividades agropecuarias y la minería. En este trabajo se revisaron las investigaciones realizadas en la Cordillera de Mérida (Venezuela) sobre los cambios en la estructura, diversidad y respuesta a disturbios de la vegetación a lo largo del ecotono bosque-páramo. Los resultados de estas investigaciones documentan la alta riqueza de especies y formas de vida que caracterizan la vegetación del ecotono, así como una alta tasa de recambio de especies a lo largo del gradiente de elevación. Así mismo, estudios sobre la dinámica sucesional de la vegetación en áreas intervenidas por la agricultura en diferentes posiciones a lo largo de la transición, indican que las leñosas del bosque muestran una capacidad mucho menor de colonización de las áreas perturbadas que las especies dominantes del páramo. Con base en la evidencia disponible, se discuten elementos estructurales que pueden servir de base para la delimitación de los ecosistemas presentes en la zona de transición y se enfatiza la necesidad de diseñar estrategias de conservación y restauración asistida que partan de interpretar el ecotono bosque-páramo como una unidad dinámica integrada.
Doi: https://doi.org/10.15446/abc.v20n3.46721.
Artículo de revisión/Review article
ESTRUCTURA, DIVERSIDAD Y DINÁMICA DE LA VEGETACIÓN EN EL ECOTONO BOSQUE-PÁRAMO: REVISIÓN DE LA EVIDENCIA EN LA CORDILLERA DE MÉRIDA
Vegetation Structure, Diversity and Dynamics in the Forest-Paramo Ecotone: A Review of the Evidence in the Cordillera de Mérida
Luis Daniel LLAMBÍ1
1 Profesor Asociado, Instituto de Ciencias Ambientales y Ecológicas, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101,0426/5770977, Venezuela.
For correspondence. llambi@ula.ve,ldllambi@gmail.com.
Received: 22nd October 2014, Returned for revision: 20th December 2014, Accepted: 13th February 2015.
Associate Editor: Susana Raquel Feldman.
Citation / Citar este artículo como: Llambí LD. Estructura, diversidad y dinámica de la vegetación en el ecotono bosque-páramo: revisión de la evidencia en la Cordillera de Mérida. Acta biol. Colomb. 2015;20(3):5-19. doi: https://doi.org/10.15446/abc.v20n3.46721.
RESUMEN
En los Andes tropicales, el límite altitudinal de los bosques corresponde a una compleja zona de transición entre el bosque paramero y el páramo andino. En el límite inferior de esta zona ocurre a su vez una transición gradual entre el bosque y las selvas nubladas. La ordenación del territorio en estos ecosistemas se ha convertido en un tema prioritario en virtud de la creciente amenaza que representan para su conservación las actividades agropecuarias y la minería. En este trabajo se revisaron las investigaciones realizadas en la Cordillera de Mérida (Venezuela) sobre los cambios en la estructura, diversidad y respuesta a disturbios de la vegetación a lo largo del ecotono bosque-páramo. Los resultados de estas investigaciones documentan la alta riqueza de especies y formas de vida que caracterizan la vegetación del ecotono, así como una alta tasa de recambio de especies a lo largo del gradiente de elevación. Así mismo, estudios sobre la dinámica sucesional de la vegetación en áreas intervenidas por la agricultura en diferentes posiciones a lo largo de la transición, indican que las leñosas del bosque muestran una capacidad mucho menor de colonización de las áreas perturbadas que las especies dominantes del páramo. Con base en la evidencia disponible, se discuten elementos estructurales que pueden servir de base para la delimitación de los ecosistemas presentes en la zona de transición y se enfatiza la necesidad de diseñar estrategias de conservación y restauración asistida que partan de interpretar el ecotono bosque-páramo como una unidad dinámica integrada.
Palabras clave: Andes tropicales, cambio climático, disturbio agrícola, formas de vida, gradiente altitudinal, sucesión secundaria.
ABSTRACT
In the tropical Andes, the mountain treeline corresponds to a complex transition zone between páramo forests and páramos. In the lower limit of this ecotone there is a more or less gradual transition between mountain cloud forests and páramo forests. Land planning in these ecosystems has become a highly topical subject given the growing threat for conservation generated by agricultural and mining activities. In this paper we review the research carried out in the Cordillera de Mérida (Venezuela) on the changes in vegetation structure, diversity and response to disturbance along the forest-páramo transition zone. The results of these studies document the high species and life form richness which characterize the ecotone vegetation, as well as a high rate of species turnover (beta diversity) along the elevation gradient. Studies on elevation changes in vegetation successional dynamics in plots disturbed by agriculture at different altitudes along the transition zone indicate that forest woody species have a lower capacity to colonize disturbed areas than dominant páramo species, a process that results in forests patches with a paramo physiognomy. On the basis of the available evidence, the structural elements that could be used for delimiting the different ecosystems present along transition zone are discussed, emphasizing the need to design conservation and restoration strategies which interpret the forest-páramo ecotone as a dynamic and integrated system.
Keywords: agricultural disturbance, climate change, elevation gradient, life forms, secondary succession, tropical Andes.
INTRODUCCIÓN
El interés por el estudio del límite altitudinal montano de los bosques tiene una larga tradición entre los naturalistas, botánicos y ecólogos (Humboldt y Bonpland, 1807; Brockmann-Jerosch, 1919; Cuatrecasas, 1934; Troll, 1973; Acosta-Solís, 1984; Vareschi, 1992). Entender los factores que regulan la posición altitudinal y dinámica de este conspicuo ecotono entre la vegetación de los bosques montanos y la vegetación abierta de las zonas alpinas, podría de hecho considerarse una de las "pruebas de fuego" que enfrenta la ecología. En el caso de los Andes Tropicales, la delimitación de la altitud y distribución espacial de la zona de transición bosque-páramo se ha convertido además en un tema muy polémico, ya que en casos como el de Colombia y Ecuador, se ha desarrollado legislación que protege explícitamente a los páramos de diferentes actividades antrópicas y especialmente, de la creciente amenaza de la minería (ej. el Código de Minas de Colombia).
El objetivo del presente trabajo es presentar una revisión de las investigaciones realizadas en la Cordillera de Mérida, en los Andes de Venezuela, sobre la estructura y dinámica de la vegetación en la zona de transición bosque-páramo. En primer lugar se presenta una discusión de carácter más general sobre la definición de la zona de transición y los ecosistemas que la conforman en los Andes Tropicales del Norte (Venezuela, Colombia y Ecuador), analizando los principales procesos que influencian su posición altitudinal y dinámica. En segundo lugar, se revisan las investigaciones realizadas sobre los patrones en la estructura y diversidad de la vegetación a lo largo de la zona de transición en Venezuela. Luego, se discute la evidencia sobre los cambios en la dinámica de la vegetación a lo largo del ecotono bosque-páramo en Venezuela, contrastando la respuesta en zonas no intervenidas y áreas en sucesión secundaria post-agrícola. Finalmente, se discuten las implicaciones de estas investigaciones sobre la estructura y dinámica de la transición bosque-páramo para el diseño de estrategias de conservación, gestión y restauración ecológica en los altos Andes tropicales.
Definiciones: tipología de la vegetación de la transición bosque-páramo
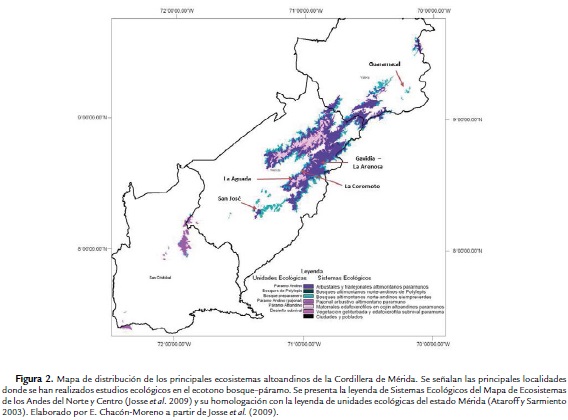
Existen muchas definiciones y conceptos en la literatura asociados al límite altitudinal del bosque y de los árboles en zonas montanas a nivel global (conocidos en la literatura en inglés como "forest line" y "treeline", respectivamente, Körner y Paulsen, 2004). En el caso de los Andes tropicales, se consideró apropiado utilizar la interpretación propuesta por Bader et al. (2007a), entendiendo el ecotono como una zona compleja de transición entre los bosques de la alta montaña tropical y los páramos, con lenguas e islas de bosques que se interdigitan con la vegetación de porte más bajo de los páramos (Ramírez et al., 2009; Fig. 1).
Estructura y dinámica de la vegetación del ecotono bosque-páramo

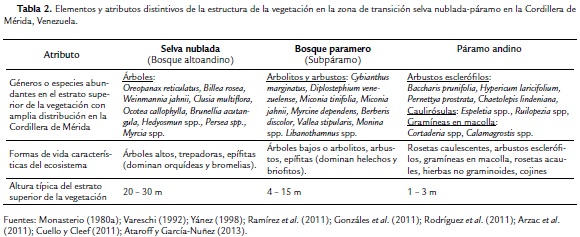
Los bosques de la zona de transición se conocen en Venezuela como bosques parameros (Monasterio 1980a) y en Colombia y Ecuador como subpáramos (Cuatrecasas, 1934; Cuatrecasas, 1958). El estrato superior de estos bosques está dominado por arbustos altos y árboles pequeños o arbolitos, generalmente con un porte menor a los 10 m. De hecho, a lo largo de la banda altitudinal de los bosques parameros o subpáramos se observa en muchos casos una disminución progresiva de la altura máxima de las leñosas (lo que en la literatura alpina se conoce como "elfin forests" o "krummhol", (Körner, 2012)). Partiendo de esta observación, Cleef (1981) dividió al subpáramo en dos cinturones: el subpáramo superior dominado por arbustos enanos y el subpáramo inferior dominado por arbustos altos y arbolitos.
En el límite altitudinal inferior del ecotono ocurre a su vez una transición más o menos gradual entre el bosque paramero continuo y las selvas altoandinas (Fig. 1). Estas selvas se conocen generalmente en Venezuela como selvas nubladas o de neblina (Vareschi, 1992) y en Colombia y Ecuador como bosques altoandinos (Cuatrecasas, 1958; Acosta-Solis, 1984). El estrato superior de la selva nublada está dominado por árboles de porte aslto (entre 20 y 40 m). Las selvas nubladas están generalmente ubicadas en las vertientes húmedas de los Andes y se caracterizan por una alta frecuencia de neblinas y nubes bajas. Sin embargo, en el caso de las vertientes más secas asociadas a bolsones secos interandinos, el límite inferior de la zona de transición es entre el bosque paramero y diferentes formaciones de bosques estacionalmente secos (Ataroff, 2003; Suárez y Chacón-Moreno, 2011).
Por otro lado, el límite superior del ecotono corresponde a la zona donde pequeñas islas de bosque paramero se encuentran insertas en diferentes comunidades de páramo andino (Monasterio, 1980a, Fig. 1), al que Cuatrecasas (1958) denomina páramo propiamente dicho. Entre los tipos fisionómicos de páramo andino más comunes en la zona de transición se encuentran arbustales parameros, rosetales-arbustales y diferentes tipos de pajonales. Los páramos bajos de la transición se caracterizan por la presencia de gramíneas en macolla, rosetas caulescentes, arbustos esclerófilos enanos y hierbas no graminoides; sin embargo, pueden observarse en muchos casos, como elementos dispersos, arbolitos y arbustos altos de varias especies del bosque, incluyendo géneros como Diplostephium, Gynoxis y Buddleja (Luteyn, 1999).
Finalmente, por encima de la zona de la transición entre el bosque continuo y el páramo, alcanzando altitudes que pueden variar entre los 3500 y más de 4200 m s.n.m., se encuentran en algunos casos pequeños bosques aislados de Polylepis. Estos bosquetes están generalmente inmersos en la matriz del páramo andino, siendo posible encontrarlos también en zonas a mayor elevación en el páramo altoandino (Monasterio, 1980a) o superpáramo (Cuatrecasas, 1958,Fig. 1); los bosquetes de Polylepis están frecuentemente asociados a "refugios post-glaciales" con condiciones ambientales muy particulares: sobre laderas y escarpes con bloques de rocas, bordes de lagunas parameras, etc. (Monasterio, 1980a). Estos bosques son muy relevantes en el contexto del estudio de las estrategias adaptativas que permiten a los árboles sobrevivir a grandes elevaciones (ej. Rada et al., 1996; Rada et al., 2011). Sin embargo, deben ser distinguidos claramente de los bosques parameros de la zona de transición, ya que tienden a aparecer como parches aislados más o menos distantes del límite forestal continuo y presentan un composición de leñosas diferente y menos diversa, aun cuando pueden compartir algunos elementos arbustivos (Vareschi, 1992).
Factores que influencian la elevación y dinámica de la transición bosque-páramo
A nivel global, la posición altitudinal del límite del bosque ha sido vinculada con limitaciones térmicas para el crecimiento de los árboles (Körner y Paulsen, 2004). Sin embargo, se reconoce que a nivel regional y local, muchos otros procesos incluyendo cambios en los regímenes de disturbio como el fuego y el pastoreo, las interacciones bióticas de competencia o facilitación y limitaciones en los procesos de dispersión, pueden también modular su dinámica (Körner,2012).
La altitud y conformación del ecotono bosque-páramo en los Andes Tropicales puede ser muy variable y responder de forma compleja a la heterogeneidad espacio-temporal Andina, aún en áreas sin intervención antrópica, en función de factores como el grado de aislamiento y altitud máxima del macizo montañoso, la estacionalidad de las precipitaciones, la exposición, topografía y las características de los suelos (Monasterio y Reyes, 1980; van der Hammen y Cleef, 1986; Bader et al., 2007b; Cuello y Cleef, 2011; Ataroff y García-Nuñez, 2013). La tendencia general es que el ecotono se ubique a menores elevaciones en: a) montañas más bajas o aisladas (el llamado efecto "top", "telescopio" o "Massenerhebung"; van Steenis, 1961; Grubb, 1971); b) en vertientes más secas y con mayor estacionalidad hídrica (Ataroff, 2003); y c) en áreas sujetas a intervención antrópica (fuegos, pastoreo, agricultura), un proceso conocido como "paramización" del bosque (Cortés et al., 1999).
En cuanto a la dimensión temporal, la posición altitudinal del límite bosque-páramo ha mostrado una respuesta muy dinámica durante los ciclos glaciares-interglaciares del Pleistoceno y los cambios más recientes durante el Holoceno (van der Hammen, 1974; Salgado-Laboriau, 1991; Wille etal., 2002; Bakker etal., 2008). Sabemos por ejemplo que durante el último máximo glacial, la temperatura llegó a ser 7 a 8 °C más baja, el clima se hizo generalmente más seco y la posición altitudinal del límite llegó a descender entre 800 y 1000 m s.n.m., por debajo de su posición actual. Esta respuesta tan dinámica a los cambios ambientales en el pasado, sugiere que la estructura y distribución del ecotono bosque-páramo pudiera cambiar también como respuesta al cambio climático global actual, con un aumento de temperatura esperado de 2-4 °C en el norte de Sur América para el año 2100 (Rull et al., 2007). El aumento de temperatura pudiera resultar, especialmente en ausencia de disturbios como el fuego o el pastoreo, en un desplazamiento hacia arriba del bosque (o elementos leñosos dominantes del bosque) sobre el páramo (Bader et al., 2007b; Suárez y Chacón-Moreno, 2011; Llambí et al., 2013a).
Sin embargo, dada la marcada heterogeneidad de la zona de transición y nuestra limitada comprensión de los procesos abióticos y bióticos que controlan la dinámica de la vegetación, se hacen necesarios estudios detallados sobre estos procesos, para poder predecir la velocidad de respuesta de la vegetación del límite bosque-páramo frente al cambio climático. Esto es particularmente cierto si consideramos la complejidad resultante de la interacción entre los cambios en el uso de la tierra y el cambio climático global (Sarmiento y Frolich, 2002; Young y León, 2007; González et al., 2011).
Aun cuando a lo largo de los Andes del Norte existen zonas bien conservadas, el ecotono bosque-páramo ha sido objeto de importantes procesos de intervención antrópica (Hofstede et al., 1998; Sarmiento y Frolich, 2002; Velasco-Linares y Vargas, 2008; Sarmiento y Llambí, 2011). Estas transformaciones incluyen: a) actividades agrícolas intensivas y semi-tradicionales (especialmente para la producción de papa, hortalizas y trigo); b) ganadería generalmente extensiva y las quemas asociadas al manejo de los pastos en algunas zonas; c) forestación con especies exóticas (i.e. pinos y eucaliptos); y d) actividades mineras. Los cambios generados por estas actividades humanas, actuando en muchos casos de forma conjunta (ej. fuego-pastoreo, agricultura-pastoreo, minería-forestación) han ocasionado el remplazo y degradación de extensas zonas de las selvas nubladas, bosques parameros y páramos andinos. Adicionalmente, en muchos casos han significado una marcada alteración de la posición altitudinal y dinámica del ecotono bosque-páramo (ej. descenso altitudinal, límites abruptos, sucesiones detenidas), generándose una sustitución de la vegetación boscosa por una vegetación abierta secundaria dominada por especies del páramo con buena capacidad de colonización (Kok et al., 1995; Cortés et al., 1999; Wille et al., 2002; Bader et al., 2007a; Velasco-Linares y Vargas, 2008; González et al., 2011).
Así, debemos ser capaces de analizar la interacción entre procesos como el aumento en la temperatura, que pudieran inducir un ascenso altitudinal del bosque, y procesos que inducen el desplazamiento de su límite por debajo de su óptimo climático (ej. paramización inducida por disturbios como la agricultura, el fuego y el pastoreo).
Estructura y diversidad de la vegetación de la transicion bosque-paramo en la Cordillera de Mérida
Los estudios detallados sobre la estructura y diversidad de la vegetación en el ecotono bosque-páramo en Venezuela se han concentrado en unas cinco localidades en la Cordillera de Mérida (Fig. 2). Este constituye el principal macizo montañoso del país, con elevaciones máximas de 4980 m (Pico Bolívar), 100 km de ancho y 450 km de extensión en dirección suroeste a noreste. En esta región, una proporción muy importante de la vegetación en el ecotono bosque-páramo se encuentra dentro de áreas protegidas (c. 53 %), incluyendo los Parques Nacionales Páramos del Batallón y La Negra, Sierra Nevada, Sierra de La Culata y Guaramacal (Monasterio y Molinillo, 2003).
Entre los estudios pioneros en la Cordillera de Mérida que presentan una visión general sobre las características del bosque paramero y sus diferencias con las formaciones vegetales del páramo andino, están los clásicos de Vareschi de 1980 ("'Vegetationsõkologie der Tropen", reeditado en español en 1992) y Monasterio (1980a, "Las formaciones vegetales en los páramos de Venezuela").
Monasterio (1980a) describe al menos tres asociaciones vegetales distintas de bosques parameros en diferentes zonas y condiciones ambientales en la cordillera: los bosques de Espeletia neriifolia Sch.Bip., Aragoa cupressina Kunth y Alnus spp.
Por su parte, Vareschi (1992) describe los bosques parameros como una formación vegetal distinta a las selvas nubladas y los llama "bosques bajos de los altos Andes" o "chirivitales" (nombre vernáculo en los Andes Venezolanos); así mismo, enfatiza que aunque tienen un porte más bajo (generalmente con un estrato emergente de entre 4 y 10 m), pueden incluso llegar a ser más ricos en especies y estructuralmente más complejos que la franja superior de la selva nublada, al combinar elementos florísticos de estas, con elementos propios del subpáramo y del páramo andino. En estos bosques, distingue al menos tres estratos: a) uno emergente con árboles pequeños y arbustos altos; b) uno rico en formas arbustivas y subfrútices; y c) uno herbáceo más bajo y variable. En el rodal analizado por Vareschi utilizando un enfoque fitosociológico en las cercanías de la Laguna Coromoto (3.400 m s.n.m., Sierra Nevada de Mérida), identificó 50 especies de plantas vasculares y una alta diversidad de formas de vida y de morfologías y tamaños de las láminas foliares.
Posteriormente, una serie de trabajos en el sector La Aguada del sistema teleférico de Mérida (Parque Nacional Sierra Nevada) han permitido contar con una visión más detallada de los cambios en la estructura y diversidad de la vegetación a lo largo del gradiente altitudinal entre la selva nublada y el páramo andino en una zona con buen estado de conservación (Yánez, 1998; Berg y Suchi, 2000; Ramírez et al., 2009; Llambí et al., 2013b).
Yánez (1998) realizó un análisis de los cambios en la composición y riqueza de especies en una transecta continua (1240 m de longitud y 2 m de ancho) entre 2850 m s.n.m. (selva nublada montana alta) y 3600 m s.n.m. (páramo de pajonal-rosetal), registrando 287 especies de plantas vasculares, siendo la zona de máxima riqueza la correspondiente al límite superior del bosque (31003300 m s.n.m.); mientras que en la parte alta de la selva nublada, por debajo de los 3000 m s.n.m., la riqueza de especies en segmentos de la transecta de 2 x 20 m osciló entre 25 y 42 especies de plantas vasculares, en la zona de transición bosque paramero - páramo la riqueza varió entre un mínimo de 32 y un máximo de 54 especies. Al igual que lo señalado por Vareschi (1992), Yánez (1998) relaciona este aumento local en la riqueza de especies con la coexistencia en el ecotono de elementos florísticos de la selva nublada montana alta (ej. Podocarpus oleifolius D.Don, Weinmannia fagaroides Kunth) y del páramo (ej. Calamagrostispittieri Hack., Ruilopezia atropurpurea (A.C.Sm.) Cuatrec.) con elementos propios del bosque paramero (ej. Diplostephium venezuelense Cuatrec., Berberis discolorTurcz.).
Mientras Yánez (1998) interpretó el ecotono con un continuo en que las especies se van alternando a lo largo del gradiente altitudinal, Ramírez et al. (2009) y Llambí et al. (2013b) analizaron en más detalle los contrastes y discontinuidades en la composición y abundancia de especies en los diferentes tipos fisionómicos de vegetación que configuran el mosaico del paisaje en la misma zona de estudio (3250-3550 m s.n.m.): a) los bosques parameros continuos; b) las lenguas e islas de bosque y sus bordes de contacto con el páramo; y c) los páramos de pajonal-rosetal a diferentes altitudes.
Estos autores encontraron diferencias marcadas en la composición y abundancia de especies del estrato leñoso entre los bosques continuos y las lenguas e islas de bosque más arriba en el gradiente (Fig. 3), sugiriendo que las islas boscosas en la zona superior del ecotono no constituyen relictos o remanentes de los bosques continuos producto de la intervención (ver también el estudio de Wille et al., 2002 en Ecuador); la riqueza de plantas leñosas disminuyó progresivamente desde los bosques continuos y lenguas de bosque (15 especies) hasta los páramos a 3550 m s.n.m. (una especie). Las islas de bosque estaban dominadas por leñosas abundantes también en los bordes de las lenguas de bosque como D. venezuelense, Hesperomeles ferrugínea Lindl. y Cybianthus marginatus (Benth.) Pipoly, que a su vez resultaron dominantes en el estrato leñoso disperso de los páramos más bajos en la zona (Fig. 3). Estos resultados sugieren que estas especies típicas de las islas y bordes del bosque, y de los páramos bajos, pudieran potencialmente actuar como pioneras en escenarios de avance del bosque sobre el páramo producto del cambio climático (ver Bader et al., 2007b; Llambí et al., 2013a). Sin embargo, Llambí et al., (2013a) no encontraron evidencia de que individuos aislados de D. venezuelense establecidos en los páramos abiertos en la zona del ecotono facilitaran a su vez el establecimiento de otras especies leñosas bajo sus copas (es decir, de que actuaran como pioneras en un proceso de avance del bosque promovido por procesos de nodricismo).
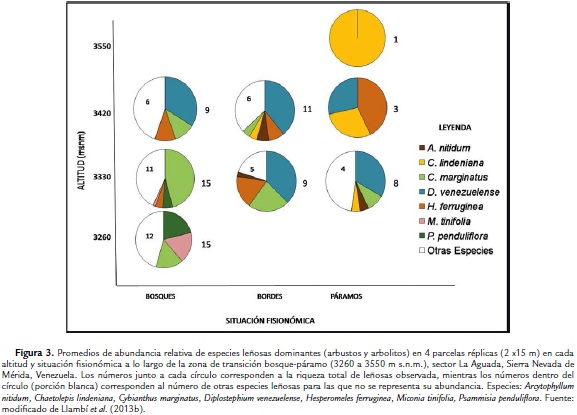
Otro estudio realizado en las zonas basal y apical de las lenguas de bosque en la transición bosque-páramo fue realizado por Rodríguez et al., (2011) en el sector de San José de la Sierra Nevada de Mérida (3150 m s.n.m.). Estos autores también reportaron un cambio claro en la abundancia de árboles y arbustos altos entre el interior, los bordes y el exterior de estas lenguas de bosque: Clusia multiflora Kunth. y C. marginatus fueron las especies dominantes en el interior mientras que D. venezuelense resultó la leñosa dominante en los bordes de las lenguas y la única encontrada en los páramos abiertos adyacentes.
Por su parte, en un estudio realizado en el páramo de Gavidia en la Sierra Nevada de Mérida por Llambí et al., (2013c), se comparó la riqueza y diversidad de plantas vasculares de los bosques parameros con las diferentes comunidades de páramo en la zona, estudiando ocho parcelas de 20 x 20 m en cada caso (Tabla 1). Tanto la riqueza observada y estimada con el índice de Jackknife (Palmer, 1990), como los índices de alfa diversidad de Shannon y Weaver (1949) y Kempton y Taylor (1978), fueron significativamente mayores en los bosques parameros que en los páramos ubicados a mayor altitud; por su parte, la proporción de especies propias o exclusivas de cada comunidad vegetal varió de 58 % en los bosques a 13-24 % en los páramos (Tabla 1). Así mismo, Llambí et al. (2013c) señalan que de las 105 especies de plantas presentes en los bosques, 20 % son endémicas del país.

Por otro lado, las formaciones y asociaciones vegetales presentes a lo largo del gradiente altitudinal entre las selvas nubladas y los páramos en el extremo noreste de la Cordillera de Mérida, en el Parque Nacional Guaramacal, han sido objeto de una serie de estudios detallados utilizando un enfoque fitosociológico (Cuello, 2002; Cuello y Cleef, 2009a; Cuello y Cleef; 2009b, Cuello y Cleef, 2011). En el límite superior de la vegetación boscosa, estos autores describen tres asociaciones de bosques enanos: los bosques continuos de Libanothamnus griffinii Cuatrec. y Geissanthus andinus MezMiconia jahnii Pittier y los parches o islas de bosque de Gaultheria anastomosans (Mutis ex L.f.) Kunth-Hesperomeles obtusifolia (D.C.) Lindl. (todos con altas abundancias en el estrato superior de C. marginatus). Entre los aspectos resaltantes de la vegetación del ecotono bosque-páramo en esta zona están: a) su ubicación a elevaciones relativamente bajas, entre los 2750 m s.n.m. y la altura máxima del macizo a 3130 m s.n.m., lo que los autores relacionan con el efecto "top" descrito por Grubb (1971); y b) la tendencia a una disminución en la riqueza de especies entre la franja superior de la selva nublada y los bosques parameros, con solo 27 y 31 especies registradas respectivamente en las dos asociaciones boscosas descritas a mayor altitud.
Finalmente, vale la pena resaltar que otra de las estrategias utilizadas para analizar los cambios estructurales a lo largo del ecotono bosque-páramo ha sido el uso de esquemas de clasificación de las especies en formas de vida (Vareschi, 1992; Arzac et al., 2011). Esta estrategia tiene la ventaja de permitir una aproximación que facilita el análisis comparativo entre localidades con composición florística diferente, así como la interpretación de los patrones de abundancia en términos de las estrategias adaptativas y respuestas funcionales de las plantas a los cambios en los factores limitantes ambientales (ej. temperatura, precipitación, radiación incidente). Un estudio realizado por Arzac et al. (2011) a lo largo de varias transectas en la zona de transición bosque-páramo en La Aguada reportó una disminución relativamente abrupta de la abundancia de los árboles, trepadoras, epífitas y helechos mesófilos a lo largo de la zona superior del ecotono, y un aumento de la importancia relativa de formas de vida características de los páramos abiertos como las rosetas caulescentes pubescentes, los arbustos esclerófilos de hojas más pequeñas (leptófilos) y las rosetas acaules (Fig. 4). A su vez, los resultados de este estudio enfatizan la alta tasa de recambio de formas de vida (diversidad beta) que caracteriza la vegetación de la transición y el efecto que pequeñas diferencias en la orientación de las laderas puede tener sobre la distribución de formas de vida como los arbustos con diferentes tamaños de láminas foliares y las rosetas caulescentes pubescentes vs. glabras.

A manera de síntesis, en la Tabla 2 se presenta una revisión comparativa de algunos aspectos distintivos de la estructura de la vegetación a lo largo de la zona de transición entre la franja superior de las selvas nubladas, los bosques parameros y los páramos andinos en la Cordillera de Mérida. Estos atributos estructurales pudieran servir como elementos claves en procesos de definición y demarcación de los diferentes ecosistemas de la transición.
Regeneración y dinámica de la transición bosque-páramo en respuesta a disturbios
Como se mencionó arriba, en la Cordillera de Mérida, una alta proporción del ecotono-bosque páramo está incluido dentro de áreas protegidas (Josse et al., 2009). Sin embargo, en los valles altos Andinos se desarrollaron a partir de la Colonia actividades agropecuarias a lo largo de la transición, con un sistema trigo-ganadería extensiva entre los siglos XVI y XIX (especialmente en los valles secos interandinos) y un sistema papa-ganadería extensiva que tiene su auge a partir de 1950 (Monasterio, 1980b; Monasterio y Molinillo, 2003). Una modalidad importante de producción han sido los agroecosistemas con descansos largos para la producción de papa, manteniéndose su actividad en algunos casos dentro de los límites de las áreas protegidas (ej. bajo figuras de zonificación como las zonas de uso poblacional autóctono).
Estos sistemas han venido sufriendo en los últimos años un proceso marcado de intensificación, que ha implicado un aumento en el uso de insumos (agrotóxicos, gallinazo, etc.) y una reducción o eliminación de los descansos (Sarmiento y Llambí, 2011). Sin embargo, los sistemas con descansos largos nos han ofrecido una oportunidad única para el estudio de los procesos de sucesión secundaria y de respuesta de la vegetación al disturbio agrícola.
En el caso del páramo de Gavidia (Parque Nacional Sierra Nevada) a partir de 1990 se desarrolló un programa de investigación de largo plazo sobre la dinámica de la vegetación y el funcionamiento de estos agroecosistemas (véase revisión en Sarmiento y Llambí, 2011). Sarmiento et al. (2003), estudiaron una serie de parcelas entre 1 y 12 años de descanso, mostrando que durante la sucesión se produce un cambio marcado en la estructura de la vegetación con una recuperación relativamente rápida de la fisionomía, abundancia de formas de vida y estructura de la vegetación del páramo de rosetal-arbustal, aunque la riqueza de especies en las áreas en sucesión (20 especies promedio por parcela) no alcanza los niveles registrados en los páramos no cultivados de la parte alta del valle (con un promedio de 35 especies). Por otro lado, en la parte baja del valle, la vegetación característica de los fragmentos remanentes de bosque paramero no parece mostrar la misma capacidad de regeneración natural. De hecho, en estas zonas bajas, las parcelas sucesionales tardías son colonizadas preferencialmente por especies dominantes de los páramos más altos de rosetal-arbustal, como la roseta caulescente Espeletia schultzii Wedd. y los arbustos esclerófi los Baccharis prunifolia Kunth. e Hypericum laricifolium Juss. (Sarmiento et al., 2003; Posada, 2013, ver Tabla 3).

A su vez, Jiménez (2013) y Bueno y Llambí (2015) reportaron que aun cuando las densidades de plántulas y juveniles de leñosas dominantes del bosque paramero (B. discolor, Vallea stipularis L.f. , Chaetolepis lindeniana (Naudin) Triana) son extremadamente bajas en las parcelas en sucesión, los pocos individuos que logran colonizar las áreas en sucesión parecen hacerlo preferencialmente bajo la protección de plantas ya establecidas, especialmente de H. laricifolium y B. prunifolia. Este mismo efecto "nodriza" generado por las copas compactas de H. laricifolium fue reportado por Llambí et al. (2013a) al analizar los patrones de establecimiento de otra leñosa dominante del límite superior del bosque paramero (D. venezuelense) en páramos bien conservados en el límite superior del bosque en La Aguada. Esto sugiere que los procesos de nodricismo-facilitación pueden ser importantes en el establecimiento de las leñosas del bosque tanto en áreas intervenidas en sucesión, como en áreas bien conservadas justo por encima de la transición bosque-páramo.
Por otro lado, en un sector cercano a Gavidia ubicado en la vertiente llanera de la Sierra Nevada (La Arenosa), el abandono de las zonas de cultivo a todo lo largo del ecotono bosque-páramo en los años 1960s permitió analizar el proceso de sucesión secundaria en las pequeñas parcelas abandonadas a mucho más largo plazo (González, 2010; González et al., 2011). En la Figura 5 se presenta un análisis multivariado de ordenación basado en datos de cobertura de plantas vasculares en las parcelas en sucesión post-agrícola vs. áreas de control no cultivadas a lo largo de un gradiente altitudinal en la zona. En este análisis se observa claramente como la estructura de la vegetación de las parcelas en el páramo es mucho más similar entre las áreas control y en sucesión que en el caso de las parcelas del bosque paramero, que muestran grandes diferencias estructurales florísticas entra parcelas control e intervenidas; de hecho, las parcelas sucesionales del bosque se desplazan en el diagrama hacia el extremo ocupado por los páramos, indicando una mayor similaridad florística con estos.

De modo que, al igual que en el caso de Gavidia, estos estudios sugieren una mayor capacidad de regeneración o resiliencia de la vegetación del páramo respecto a la del bosque paramero, observándose el proceso de paramización del bosque y una capacidad muy limitada de colonización en las áreas intervenidas de las leñosas características del bosque denso (ej. Myrcianthes myrsinoides (Kunth) Grifo), aún después de más de cinco décadas de regeneración espontánea. De hecho, las parcelas en sucesión en las zonas más abajas del gradiente estaban dominadas por especies parameras como los arbustos Acaena elongata L. e H. laricifolium y la roseta caulescente E. schultzii, observándose también abundante colonización de juveniles y adultos de arbolitos que son importantes en los arbustales más arriba en el gradiente (ej. Myrsine dependens (Ruiz & Pav.) Spreng. y H. ferruginea) (Tabla 3).
Implicaciones para la conservación y la restauración ecológica
El conocimiento acumulado en las últimas décadas sobre el complejo escenario del ecotono bosque-páramo encierra, en nuestra opinión, algunas lecciones para la gestión y la conservación de la biodiversidad en los Andes tropicales. En primer lugar, este ecotono es una de las manifestaciones más evidentes de la gran diversidad de especies, formas de vida, ecosistemas y paisajes que caracterizan a este "hotspot" global de biodiversidad; un patrimonio único que plantea interesantes desafíos para su estudio y conservación.
Los trabajos reseñados sobre la estructura de la vegetación no solo muestran una alta riqueza de especies en la zona de transición, sino una alta tasa de cambio en la composición y abundancia de especies y formas de vida a lo largo del continuo entre la franja alta de la selva nublada, el bosque paramero y el páramo (Tabla 1, Figs. 3 y 4). Así mismo, la evidencia disponible sugiere que existen marcadas diferencias en la composición florística entre las localidades estudiadas en la Cordillera de Mérida, es decir, una alta diversidad a escala regional (diversidad gamma).
Aun así, es posible establecer elementos diagnósticos de validez general que permiten la delimitación de los diferentes ecosistemas que conforman la zona ecotonal. Estos incluyen aspectos estructurales como la altura máxima del dosel y la estratificación vertical de la vegetación, géneros y especies de amplia distribución geográfica pero que muestran diferencias claras en su abundancia a diferentes elevaciones, así como algunas formas de vida características de cada ecosistema (teniendo particular importancia la dominancia de árboles altos, arbolitos y arbustos esclerófi los como elementos diagnósticos de las selvas, bosques de la transición y páramos).
Por otro lado, la investigación sobre la dinámica de respuesta del ecotono bosque-páramo también encierra lecciones importantes. En primer lugar, la evidencia palinológica muestra que no solo las selvas y bosques, sino también los páramos, tuvieron una amplia extensión mucho antes de la colonización humana de los espacios altoandinos (van der Hammen, 1974; Salgado-Labouriau, 1991). Asimismo, la evidencia en zonas poco intervenidas sugiere que características estructurales del ecotono que son atribuidas a veces a la mano del hombre, como la presencia de islas de bosque dentro de la matriz del páramo, no necesariamente son el resultado de la intervención antrópica (Wille et al., 2002; Ramírez et al., 2009).
Es claro que en zonas extensas las selvas y bosques de la alta montaña tropical (particularmente en algunas zonas de Ecuador o el sur de Colombia) han sufrido procesos históricos de larga data de transformación hacia pajonales secundarios, producto principalmente del pastoreo y las quemas (White, 2013). Sin embargo, esto no justifica la conclusión a la que a veces se llega, de que la estructura y posición altitudinal del ecotono bosque-páramo a lo largo de todos los Andes del Norte tengan como determinante fundamental transformaciones antrópicas (Ellemberg, 1979; Sarmiento y Frolich, 2002).
En cuanto a la respuesta de la vegetación a la intervención antrópica, la investigación en sistemas con descansos largos de la Sierra Nevada de Mérida nos sugiere que el bosque paramero tiene una capacidad de regeneración y resiliencia más baja que la vegetación de los páramos. Esto parece deberse, en buena medida, a las marcadas limitaciones que enfrentan muchas de las leñosas dominantes de los bosques para colonizar y establecerse en áreas intervenidas en que se removió la cobertura del dosel, mientras que especies dominantes del páramo muestran una alta capacidad de colonización en áreas abiertas (Tabla 3).
Las investigaciones de Bader et al. (2007b) en Ecuador y de Puentes (2010) en la Sierra Nevada de Mérida sugieren que uno de los factores clave que pudieran actuar como barreras para la colonización de leñosas fuera de los bosques, es su limitada resistencia a los altos niveles de radiación característicos del páramo, vinculada al efecto de las bajas temperaturas y altas radiaciones sobre la eficiencia fotosintética de árboles dominantes del bosque. Este mecanismo pudiera a su vez generar un proceso de retroalimentación positiva entre la presencia de un dosel continuo y la regeneración de las leñosas (Bader et al., 2007b), que pudiera modular o retardar la velocidad de respuesta del bosque en escenarios de cambio ambiental (i.e. en respuesta al calentamiento global y/o a disturbios que eliminan la cobertura forestal continua). Aun así, vimos como algunas de las especies dominantes del páramo y que tienen también un alta capacidad de colonización en áreas intervenidas (ej. arbustos esclerófi los como H. laricifolium o B. prunifolia), pueden facilitar el establecimiento de juveniles de árboles y arbolitos, tanto en áreas perturbadas en sucesión, como en los páramos ubicados inmediatamente por encima del ecotono (Bueno y Llambí, 2015; Jiménez, 2013; Llambí et al., 2013a). Todo esto enfatiza las complejas y estrechas relaciones dinámicas de interdependencia que existen entre la vegetación de los bosques y los páramos.
En este sentido, la investigación sobre el ecotono bosque-páramo apunta hacia la necesidad, para el diseño de estrategias de conservación y restauración, de contar con una visión integral de las continuidades y discontinuidades características de los fenómenos ecológicos; es decir, una visión de conjunto de los elementos compartidos que confieren integridad estructural y funcional al ecotono (ej. formas de vida dominantes) y los elementos disímiles que le confieren su diversidad (ej. rápido recambio de especies). En consecuencia, es fundamental plantear estrategias y acciones de ordenación y manejo del territorio que garanticen la conservación del mosaico de ecosistemas y redes de interacciones que conforman la zona de transición (ej. flujos hídricos, procesos dinámicos de colonización e intercambio de especies entre el bosque y el páramo, etc.). En la Tabla 4 se sintetizan algunas de estas estrategias y acciones de conservación y manejo que se consideran prioritarias.

En particular, los estudios disponibles en la Cordillera de Mérida enfatizan la importancia de la conservación de los bosques parameros o subpáramos como ecosistemas de transición: aun cuando estos bosques han recibido menos atención en la literatura que las selvas nubladas y los páramos, son ecosistemas de una elevada diversidad de especies y formas de vida, una alta proporción de especies propias, un alto grado de endemismo y que han sido particularmente afectados por los procesos de transformación antrópica de los paisajes altoandinos, mostrando además una baja capacidad de regeneración frente a los disturbios.
A su vez, los acelerados procesos de transformación y fragmentación de la vegetación y la limitada capacidad de regeneración natural de los bosques del ecotono, plantean la necesidad urgente de implementar estrategias integradas de restauración asistida. Estas deben necesariamente partir de analizar las diferencias en estructura y resiliencia de los diferentes tipos de vegetación de la zona de transición bajo diferentes condiciones ambientales, regímenes de disturbio e historias de transformación (evitando partir de preconcepciones y extrapolaciones de una región a otra). Así, son necesarias menos "recetas de cocina" y más procesos de restauración adaptados al contexto, que integren a las acciones de manejo, actividades explícitas de investigación, monitoreo de largo plazo y participación de múltiples actores (Vargas, 2008).
Un tema particularmente preocupante en este sentido, es la práctica extendida a lo largo de los Andes del Norte de promover procesos de "reforestación" de los páramos y otros ecosistemas altoandinos utilizando únicamente árboles. De hecho, se siguen utilizando en muchos casos exóticas como pinos y eucaliptos y especies nativas de los Andes centrales como Polylepis racemosa, pero que son introducidas en los páramos (Segovia y Quijia, 2013). Estos procesos de forestación prestan además muy poca atención a la vegetación de referencia y la historia de uso del área a recuperar, asumiendo erróneamente que los árboles son elementos dominantes de todos los ecosistemas altoandinos, incluyendo los páramos arbustivos y de pajonal o las zonas de humedales.
Finalmente, es fundamental contar con una comprensión más mecanística de los procesos y barreras que limitan la colonización y establecimiento de la vegetación a lo largo del ecotono bosque-páramo. Esto implica profundizar el análisis del efecto que tienen sobre la dinámica de establecimiento cambios en las condiciones micro-ambientales y de nutrientes, inducidos por disturbios antrópicos o por la transición natural entre un dosel cerrado y la vegetación abierta del páramo. Asimismo, se requiere profundizar el análisis del papel que las interacciones inter-específicas de facilitación, competencia y herbivoría/pastoreo, así como la frugivoría, la dispersión de semillas y simbiosis cómo las relaciones micorrízicas cumplen en modular la capacidad respuesta de la vegetación. Este conocimiento no solo será clave para el diseño de programas de restauración más efectivos, sino para enfrentar el reto de predecir la respuesta del ecotono bosque-páramo frente al cambio climático global.
AGRADECIMIENTOS
A Patricia Velazco-Linares, el equipo de investigación del Jardín Botánico José Celestino Mutis de Bogotá y el equipo del Proyecto de Delimitación de Páramos del Instituto de Investigación de Recursos Biológicos Alexander von Humboldt de Colombia, quienes me inspiraron y animaron a escribir este trabajo. Así mismo, al entusiasta equipo de investigadores, técnicos y estudiantes del ICAE y el Postgrado en Ecología Tropical de la ULA, compañeros del proyecto LEAF (CRN II005, IAI), por sus valiosos aportes y reflexiones en torno a la dinámica del ecotono-bosque páramo en la Sierra Nevada de Mérida. A Eulogio Chacón por la elaboración del mapa de ecosistemas altoandinos en la Cordillera de Mérida incluido en esta publicación. Finalmente, a Robert Hofstede, Luis E. Gámez y dos revisores anónimos por su revisión crítica del manuscrito.
REFERENCIAS
Acosta-Solis, M. Los páramos Andinos del Ecuador. Quito: Publicaciones cientificas MAS, Quito; 1984. 220 p.
Arzac A, Chacón-Moreno E, Llambí, LD, Dulhoste R. Distribución de formas de vida de plantas en el límite superior del ecotono bosque-páramo en los Andes Tropicales. Ecotrópicos. 2011;24(1):26-46.
Ataroff M. Selvas y bosques de montaña. In: Aguilera, Azócar M, González A, Jiménez E, editors. Biodiversidad en Venezuela, Tomo II. Caracas, Venezuela: Fundación Polar, Ministerio de Ciencia y Tecnología, FONACIT; 2003. p. 762-810.
Ataroff M, García-Nuñez C. Selvas y bosques nublados de Venezuela. In: Medina E., Hubber O, Nassar J, editors. Recorriendo el Paisaje Vegetal de Venezuela. Homenaje a Volmar Vareschi. Caracas: Ediciones IVIC: 2013. 319 p.
Bader M, Rietkerk M, Bregt A. Vegetation structure and temperature regimes of tropical alpine treelines. Arct Antarct Alp Res. 2007a:39:353-364.
Bader M, van Geloof I, Rietkerk. M. High solar radiation hinders tree regeneration above the alpine treeline in northern Ecuador. Plant Ecol. 2007b;191:33-45. Doi:10.1007/s11258-006-9212-6.
Bakker J, Moscol-Olivera M, Hooghiemstra H. Holocene environmental change at the upper forest line in northern Ecuador. The Holocene. 2008;18(6):877-893. Doi: 10.1177/0959683608093525.
Berg A, Suchi S. Vegetación de los Páramos La Aguada, La Fría y Espejo en los Andes Venezolanos. Plántula.2000;3(1):47-64.
Brockmann-Jerosch H. Baumgrenze und Klimacharakter. Planzengeographische Kommission der Schweizerischen Naturforschenden Gesellschaft, Beitráge zur geobotanischen Landesaufnahme. 1919;6:1-255.
Bueno A, Llambí LD. Facilitation and edge effects influence vegetation regeneration in old-fields at the tropical Andean forest-line. Appl Veg Sci. 2015. Doi: 10.1111/avsc.12186.
Cleef A. The Vegetation of the Páramos of the Colombian Cordillera Oriental. Volume 61. Vaduz: J. Cramer; 1981.
Cortés-S SP, van der Hammen T, Rangel-Ch JO. Comunidades vegetales y patrones de degradación y sucesión en la vegetación de los cerros occidentales de Chía Cundinamarca-Colombia. Rev Acad Colomb Cienc.1999;23(89):529-554.
Cuatrecasas J. Observaciones geobotánicas en Colombia. Trabajos Museo Nacional de Ciencias Naturales, Serie Botánica. 1934;27:1-144.
Cuatrecasas J. Aspectos de la Vegetación Natural de Colombia. Rev Acad Colomb Cienc Ex Fis y Nat.1958;10(4):221-264.
Cuello N. Altitudinal Changes in Forest Diversity and Composition in the Ramal de Guaramacal in the Venezuelan Andes. Ecotrópicos. 2002;15(2):160-176.
Cuello N, CleefA M. The forests of Ramal de Guaramacal in the Venezuelan Andes. Phytocoenologia. 2009a;39(1):109-156. Doi: 10.1 127/0340-269X/2009/0039-0109.
Cuello N, Cleef AM. The paramo vegetation of Ramal de Guaramacal, Trujillo State, Venezuela. I. Zonal communities. Phytocoenologia. 2009b;39(3):295-329. Doi: 10.1 127/0340-269X/2009/0039-0295.
Cuello N, Cleef AM. Bosques de los Andes de Venezuela: caso El Ramal de Guaramacal. Biollania. 2011;10:74-105.
Ellemberg H. Man's influence on tropical mountain ecosystems in South-America. J Ecol. 1979; 67:401-416.
Fariñas M. Análisis de la Vegetación y sus Relaciones Ambientales Mediante Métodos Multivariantes de Ordenamiento. Mérida: Postgrado en Ecología Tropical, Universidad de Los Andes de Venezuela; 1996. 183 pp.
González WY. Dinámica sucesional de la vegetación en la zona de transición bosque-páramo en la Cordillera de Mérida (tesis de maestría). Mérida: Postgrado en Ecología Tropical, Facultad de Ciencias, Universidad de Los Andes de Venezuela; 2010. 96 p.
González WY, Llambí LD, Smith JK, Gámez LE. Dinámica sucesional del componente arbóreo en la zona de transición bosque-Páramo en Los Andes Tropicales. Ecotrópicos. 2011;24(1):60-79.
Grubb PJ. Interpretation of the 'Massenerhebung' effect on tropical mountains. Nature. 1971; 229:44-45. Doi: 10.1038/229044a0.
Hofstede R, Lips J, Jongsma W. Geografía, Ecología y Forestación de la Sierra Alta del Ecuador. Revisión de literatura. Quito: Editorial Abya Yala;1998. 242 p.
Jiménez D. Estructura poblacional y nicho de establecimiento de especies leñosas del bosque paramero en áreas en sucesión secundaria (tesis de Maestría) Mérida: Postgrado en Ecología Tropical, Universidad de los Andes de Venezuela; 2013. 170 p.
Josse C, Cuesta F, Navarro G, Barena V, Cabrera E, Chacón-Moreno E, et al. Ecosystemas de los Andes del Norte y Centro. Lima: SGCAN-Proyecto Páramo Andino-Ecobona; 2009. 95 p.
Kempton RA, Taylor LR. The Q-statistics and the diversity of floras. Nature.1978;275:252-253.
Kok K, Verweij P, Beukema H. Effects of cutting and grazing on Andean treeline vegetation. In: Churchill SP, Balslev H, Forero E, Luteyn JL, editors. Biodiversity and Conservation of Neotropical Montane Forest. New York: The New York Botanical Garden;1995. p. 527-539.
Körner C. Alpine treelines: functional ecology of the global high elevation tree limits. 1 ed. Springer, 2012. 220 p.
Körner C, Paulsen J. A world-wide study of high altitude treeline temperatures. J Biogeogr. 2004;31(5):713-732. Doi: 10.1111/j.1365-2699.2003.01043.x.
Llambí LD, Puentes J, Garcia-Núñez C. Spatial relations and population structure of a dominant tree along a treeline ecotone in the Tropical Andes: interactions at gradient and plant-neighborhood scales. Plant Ecol Divers. 2013a;6(3-4):343-353. Doi: 10.1080/17550874.2013.810312.
Llambí LD, Ramírez L, Schwarzkopf T (b). Patrones de distribución de plantas leñosas en el ecotono bosque-páramo de la Sierra Nevada de Mérida: ¿Qué nos sugieren sobre la dinámica del límite del bosque?. In: Cuesta F, Sevink J, Llambí LD, De Bièvre B, PosnerJ, editors. Avances en investigación para la conservación de los páramos andinos. CONDESAN. Quito;2013b. p. 53-78.
Llambí LD, Fariñas M, Smith JK, Castañeda SM, Briceño B. Diversidad de la vegetación en dos paramos de Venezuela: un enfoque multiescalar con fines de conservación. In: Cuesta F, Sevink J, Llambí LD, De Bièvre B, Posner J, editors. Avances en investigación para la conservación de los páramos andinos. Quito: CONDENSAN; 2013c. p. 41-68.
Luteyn J. Paramos: A checklist ofplant diversity, geographical distribution and botanical literature. Mem NY Bot Gard.1999;84:1-278.
Monasterio M. Las formaciones vegetales de los páramos de Venezuela. In: Monasterio M, editor. Estudios Ecológicos en los Paramos Andinos. Mérida: Editorial de la Universidad de los Andes; 1980a. p. 93-158.
Monasterio M. Poblamiento humano y uso de la tierra en los altos Andes de Venezuela. In: Monasterio M, editor. Estudios Ecológicos en los Paramos Andinos. Editorial de la Universidad de los Andes. Mérida;1980b. p. 170-198.
Monasterio M, Reyes S. Diversidad ambiental y variación de la vegetación en los páramos de los Andes Venezolanos. In: Monasterio M, editor. Estudios Ecológicos en los Paramos Andinos. Mérida: Editorial de la Universidad de los Andes; 1980. p. 47-91.
Monasterio M, Molinillo M. Venezuela: El paisaje y su diversidad. In: Hofstede R, Segarra P, Mena P, editores. Los Páramos del Mundo. Quito: Global Peatland Initiative; 2003. p. 205-236.
Palmer MW. The estimation of species richness by extrapolation. Ecology. 1990;71:1 195-1198. Doi: 10.2307/1937387.
Posada J. El Papel de la Dispersión de Semillas en la Regeneración de la Vegetación del Páramo en un Paisaje Fragmentado (tesis de Maestría). Mérida: Postgrado en Ecología Tropical, Universidad de Los Andes de Venezuela; 2013. 149 p.
Puentes J. Patrones y Mecanismos de Establecimiento de dos Especies Leñosas en la Zona de Transición entre el Bosque Paramero y el Páramo en los Andes Tropicales (tesis de maestría). Mérida: Postgrado en Ecología Tropical, Universidad de los Andes en Venezuela; 2010. 130 p.
Rada F, Azocar A, González J, Briceño B, García-Núñez C. Carbon and Water Balance in Polylepis serícea, a tropical tree-lines species. Trees. 1996;10(4):218-222. Doi: 10.1007/BF02185672.
Rada F, Garcia-Núñez C, Rangel S. Microclimate and regeneration patterns of Polylepis sericea in a treeline forest of the Venezuelan Andes. Ecotrópicos. 2011;24(1):113-122.
Ramírez L, Llambí LD, Schwarzkopf T, Gámez LE, Márquez NJ. Vegetation structure along the forest-páramo transition belt in the Sierra Nevada de Mérida: implications for understanding treeline dynamics. Ecotrópicos. 2009;22(2):83-98.
Rodríguez E, Schwarzkopf T, Gámez LE, Dugarte W, Dulhoste R. Canopy structure and Woody species composition at the upper treeline in the Venezuelan Andes. Ecotrópicos. 2011;24(1):47-59.
Rull V, Vegas T, Nogué S, Montoya E, Cañellas N, Lara A. Quaternary paleoclimatology, neotropical biodiversity and potential effects of global warming. Contrib to Sci.2007;3(3):405-413.
Salgado-Labouriau ML. Palynology ofthe Venezuelan Andes. Grana. 1991;30(2):342-349.
Sarmiento FO, Frolich LM. Andean cloud forest tree lines: Naturalness, agriculture and the human dimension. Mt Res Dev. 2002;22(3):278-287.
Sarmiento L, Llambí LD, Escalona A, Marquez N. Vegetation patterns, regeneration rates and divergence in an old-field succession of the high tropical Andes. Plant Ecol. 2003; 166:63-74.
Sarmiento L, Llambí LD. Regeneración del páramo luego de un disturbio agrícola: una síntesis de 20 años de investigaciones en sistemas con descansos largos en la Cordillera de Mérida. En Herrera F, Herrera I, editors. La Restauración Ecológica en Venezuela: fundamentos y experiencias. Caracas: Ediciones Instituto Venezolano de Investigaciones Científicas; 2011. p. 123-148.
Segovia-Salgado MA, Quijia-Lamiña P. Citogeografía de Cuatro Especies de Polylepis (Rosaceae) en el Ecuador: información relevante para el manejo y conservación de los bosques Andinos. En Cuesta F, Sevink J, Llambí LD, De Bièvre B, Posner J, editors. Avances en investigación para la conservación de los páramos andinos. Quito: CONDESAN; 2013. p. 467-486.
Shannon CE, Weaver W. The Mathematical Theory of Communication. Urbana, Illinois: University of Illinois Press; 1949. 120 p.
Suarez P, Chacón-Moreno E. Modelo espacial de distribución del ecotono bosque paramo en los Andes Venezolanos. Ubicación potencial y escenarios de cambio climático. Ecotrópicos. 2011;24(1): 3-25.
Troll C. The upper timberlines in different climatic zones. Arct Antarct Alp Res. 1973;5(3):A3-A18.
van der Hammen T. The Pleistocene changes in vegetation and climate in tropical South America. J Biogeogr. 1974;1(1):3-26. Doi: 10.2307/3038066.
van der Hammen T. Plan ambiental de la cuenca alta del Rio Bogotá análisis y orientaciones para el ordenamiento territorial. 1 ed. Bogotá: Corporación Autónoma Regional; 1998. 142 p.
van der Hammen T, Cleef AM. Development of the high Andean paramo flora and vegetation. In: Vuilleumier F, Monasterio M, editors. High altitude tropical biogeography. New York: Oxford University Press; 1986.p. 153-201.
Vareschi V. Ecología de la Vegetación Tropical. 1 ed. Caracas: Edición especial de la Sociedad Venezolana de Ciencias Naturales; 1992. 306 p.
van Steenis CGGJ. An attempt towards an explanation of the effect of mountain mass elevation. Proc K Ned Akad Wet C. 1961;64:435-442.
Vargas O. Estrategia para la restauración ecológica del bosque altoandino (El caso de la reserva forestal municipal de Cogua, Cundinamarca). 2 ed. Bogotá: Universidad Nacional de Colombia; 2008. 372 p.
Velasco-Linares P, Vargas O. Problemática de los Bosques Altoandinos. En: Vargas O, editor. Estrategias para la restauración ecológica del bosque altoandino (El caso de la Reserva Forestal Municipal de Cogua, Cundinamarca). Bogotá: Universidad Nacional de Colombia; 2008. p. 41-56.
von Humboldt A, Bondpland A. Essay on the Geography of Plants. Chicago: University of Chicago Press; 1807. 296 p.
White S. Grass páramo as hunter-gatherer landscape. The Holocene. 2013;23(6):898-915. Doi: 10.1177/0959683612471987.
Wille M, Hooghiemstra H, Hofstede R, Fehse J, Sevink J. Upper forest line reconstruction in a deforested area in northern Ecuador based on pollen and vegetation analysis. J Trop Ecol. 2002;18(3):409-440. Doi: 10.1017/S0266467402002286.
Yánez P. Caracterización florística en un sector de cambio páramo- selva nublada en el parque nacional Sierra Nevada, Venezuela. Revista Forestal Venezolana. 1998;42(1):51-62.
Young K, Leon B. Tree-line changes along the Andes: implications of spatial patterns and dynamics. Philos Trans R Soc Lond B Biol Sci. 2007;362(1478):263-272. Doi: 10.1098/rstb.2006.
Referencias
Acosta-Solis, M. Los páramos Andinos del Ecuador. Quito: Publicaciones cientificas MAS, Quito; 1984. 220 p.
Arzac A, Chacón-Moreno E, Llambí, LD, Dulhoste R. Distribución de formas de vida de plantas en el límite superior del ecotono bosque-páramo en los Andes Tropicales. Ecotrópicos. 2011;24(1):26-46.
Ataroff M. Selvas y bosques de montaña. In: Aguilera, Azócar M, González A, Jiménez E, editors. Biodiversidad en Venezuela, Tomo II. Caracas, Venezuela: Fundación Polar, Ministerio de Ciencia y Tecnología, FONACIT; 2003. p. 762-810.
Ataroff M, García-Nuñez C. Selvas y bosques nublados de Venezuela. In: Medina E., Hubber O, Nassar J, editors. Recorriendo el Paisaje Vegetal de Venezuela. Homenaje a Volmar Vareschi. Caracas: Ediciones IVIC: 2013. 319 p.
Bader M, Rietkerk M, Bregt A. Vegetation structure and temperature regimes of tropical alpine treelines. Arct Antarct Alp Res. 2007a:39:353-364.
Bader M, van Geloof I, Rietkerk. M. High solar radiation hinders tree regeneration above the alpine treeline in northern Ecuador. Plant Ecol. 2007b;191:33-45. Doi:10.1007/s11258-006-9212-6.
Bakker J, Moscol-Olivera M, Hooghiemstra H. Holocene environmental change at the upper forest line in northern Ecuador. The Holocene. 2008;18(6):877-893. Doi: 10.1177/0959683608093525.
Berg A, Suchi S. Vegetación de los Páramos La Aguada, La Fría y Espejo en los Andes Venezolanos. Plántula.2000;3(1):47-64.
Brockmann-Jerosch H. Baumgrenze und Klimacharakter. Planzengeographische Kommission der Schweizerischen Naturforschenden Gesellschaft, Beitráge zur geobotanischen Landesaufnahme. 1919;6:1-255.
Bueno A, Llambí LD. Facilitation and edge effects influence vegetation regeneration in old-fields at the tropical Andean forest-line. Appl Veg Sci. 2015. Doi: 10.1111/avsc.12186.
Cleef A. The Vegetation of the Páramos of the Colombian Cordillera Oriental. Volume 61. Vaduz: J. Cramer; 1981.
Cortés-S SP, van der Hammen T, Rangel-Ch JO. Comunidades vegetales y patrones de degradación y sucesión en la vegetación de los cerros occidentales de Chía Cundinamarca-Colombia. Rev Acad Colomb Cienc.1999;23(89):529-554.
Cuatrecasas J. Observaciones geobotánicas en Colombia. Trabajos Museo Nacional de Ciencias Naturales, Serie Botánica. 1934;27:1-144.
Cuatrecasas J. Aspectos de la Vegetación Natural de Colombia. Rev Acad Colomb Cienc Ex Fis y Nat.1958;10(4):221-264.
Cuello N. Altitudinal Changes in Forest Diversity and Composition in the Ramal de Guaramacal in the Venezuelan Andes. Ecotrópicos. 2002;15(2):160-176.
Cuello N, CleefA M. The forests of Ramal de Guaramacal in the Venezuelan Andes. Phytocoenologia. 2009a;39(1):109-156. Doi: 10.1 127/0340-269X/2009/0039-0109.
Cuello N, Cleef AM. The paramo vegetation of Ramal de Guaramacal, Trujillo State, Venezuela. I. Zonal communities. Phytocoenologia. 2009b;39(3):295-329. Doi: 10.1 127/0340-269X/2009/0039-0295.
Cuello N, Cleef AM. Bosques de los Andes de Venezuela: caso El Ramal de Guaramacal. Biollania. 2011;10:74-105.
Ellemberg H. Man's influence on tropical mountain ecosystems in South-America. J Ecol. 1979; 67:401-416.
Fariñas M. Análisis de la Vegetación y sus Relaciones Ambientales Mediante Métodos Multivariantes de Ordenamiento. Mérida: Postgrado en Ecología Tropical, Universidad de Los Andes de Venezuela; 1996. 183 pp.
González WY. Dinámica sucesional de la vegetación en la zona de transición bosque-páramo en la Cordillera de Mérida (tesis de maestría). Mérida: Postgrado en Ecología Tropical, Facultad de Ciencias, Universidad de Los Andes de Venezuela; 2010. 96 p.
González WY, Llambí LD, Smith JK, Gámez LE. Dinámica sucesional del componente arbóreo en la zona de transición bosque-Páramo en Los Andes Tropicales. Ecotrópicos. 2011;24(1):60-79.
Grubb PJ. Interpretation of the 'Massenerhebung' effect on tropical mountains. Nature. 1971; 229:44-45. Doi: 10.1038/229044a0.
Hofstede R, Lips J, Jongsma W. Geografía, Ecología y Forestación de la Sierra Alta del Ecuador. Revisión de literatura. Quito: Editorial Abya Yala;1998. 242 p.
Jiménez D. Estructura poblacional y nicho de establecimiento de especies leñosas del bosque paramero en áreas en sucesión secundaria (tesis de Maestría) Mérida: Postgrado en Ecología Tropical, Universidad de los Andes de Venezuela; 2013. 170 p.
Josse C, Cuesta F, Navarro G, Barena V, Cabrera E, Chacón-Moreno E, et al. Ecosystemas de los Andes del Norte y Centro. Lima: SGCAN-Proyecto Páramo Andino-Ecobona; 2009. 95 p.
Kempton RA, Taylor LR. The Q-statistics and the diversity of floras. Nature.1978;275:252-253.
Kok K, Verweij P, Beukema H. Effects of cutting and grazing on Andean treeline vegetation. In: Churchill SP, Balslev H, Forero E, Luteyn JL, editors. Biodiversity and Conservation of Neotropical Montane Forest. New York: The New York Botanical Garden;1995. p. 527-539.
Körner C. Alpine treelines: functional ecology of the global high elevation tree limits. 1 ed. Springer, 2012. 220 p.
Körner C, Paulsen J. A world-wide study of high altitude treeline temperatures. J Biogeogr. 2004;31(5):713-732. Doi: 10.1111/j.1365-2699.2003.01043.x.
Llambí LD, Puentes J, Garcia-Núñez C. Spatial relations and population structure of a dominant tree along a treeline ecotone in the Tropical Andes: interactions at gradient and plant-neighborhood scales. Plant Ecol Divers. 2013a;6(3-4):343-353. Doi: 10.1080/17550874.2013.810312.
Llambí LD, Ramírez L, Schwarzkopf T (b). Patrones de distribución de plantas leñosas en el ecotono bosque-páramo de la Sierra Nevada de Mérida: ¿Qué nos sugieren sobre la dinámica del límite del bosque?. In: Cuesta F, Sevink J, Llambí LD, De Bièvre B, PosnerJ, editors. Avances en investigación para la conservación de los páramos andinos. CONDESAN. Quito;2013b. p. 53-78.
Llambí LD, Fariñas M, Smith JK, Castañeda SM, Briceño B. Diversidad de la vegetación en dos paramos de Venezuela: un enfoque multiescalar con fines de conservación. In: Cuesta F, Sevink J, Llambí LD, De Bièvre B, Posner J, editors. Avances en investigación para la conservación de los páramos andinos. Quito: CONDENSAN; 2013c. p. 41-68.
Luteyn J. Paramos: A checklist ofplant diversity, geographical distribution and botanical literature. Mem NY Bot Gard.1999;84:1-278.
Monasterio M. Las formaciones vegetales de los páramos de Venezuela. In: Monasterio M, editor. Estudios Ecológicos en los Paramos Andinos. Mérida: Editorial de la Universidad de los Andes; 1980a. p. 93-158.
Monasterio M. Poblamiento humano y uso de la tierra en los altos Andes de Venezuela. In: Monasterio M, editor. Estudios Ecológicos en los Paramos Andinos. Editorial de la Universidad de los Andes. Mérida;1980b. p. 170-198.
Monasterio M, Reyes S. Diversidad ambiental y variación de la vegetación en los páramos de los Andes Venezolanos. In: Monasterio M, editor. Estudios Ecológicos en los Paramos Andinos. Mérida: Editorial de la Universidad de los Andes; 1980. p. 47-91.
Monasterio M, Molinillo M. Venezuela: El paisaje y su diversidad. In: Hofstede R, Segarra P, Mena P, editores. Los Páramos del Mundo. Quito: Global Peatland Initiative; 2003. p. 205-236.
Palmer MW. The estimation of species richness by extrapolation. Ecology. 1990;71:1 195-1198. Doi: 10.2307/1937387.
Posada J. El Papel de la Dispersión de Semillas en la Regeneración de la Vegetación del Páramo en un Paisaje Fragmentado (tesis de Maestría). Mérida: Postgrado en Ecología Tropical, Universidad de Los Andes de Venezuela; 2013. 149 p.
Puentes J. Patrones y Mecanismos de Establecimiento de dos Especies Leñosas en la Zona de Transición entre el Bosque Paramero y el Páramo en los Andes Tropicales (tesis de maestría). Mérida: Postgrado en Ecología Tropical, Universidad de los Andes en Venezuela; 2010. 130 p.
Rada F, Azocar A, González J, Briceño B, García-Núñez C. Carbon and Water Balance in Polylepis serícea, a tropical tree-lines species. Trees. 1996;10(4):218-222. Doi: 10.1007/BF02185672.
Rada F, Garcia-Núñez C, Rangel S. Microclimate and regeneration patterns of Polylepis sericea in a treeline forest of the Venezuelan Andes. Ecotrópicos. 2011;24(1):113-122.
Ramírez L, Llambí LD, Schwarzkopf T, Gámez LE, Márquez NJ. Vegetation structure along the forest-páramo transition belt in the Sierra Nevada de Mérida: implications for understanding treeline dynamics. Ecotrópicos. 2009;22(2):83-98.
Rodríguez E, Schwarzkopf T, Gámez LE, Dugarte W, Dulhoste R. Canopy structure and Woody species composition at the upper treeline in the Venezuelan Andes. Ecotrópicos. 2011;24(1):47-59.
Rull V, Vegas T, Nogué S, Montoya E, Cañellas N, Lara A. Quaternary paleoclimatology, neotropical biodiversity and potential effects of global warming. Contrib to Sci.2007;3(3):405-413.
Salgado-Labouriau ML. Palynology ofthe Venezuelan Andes. Grana. 1991;30(2):342-349.
Sarmiento FO, Frolich LM. Andean cloud forest tree lines: Naturalness, agriculture and the human dimension. Mt Res Dev. 2002;22(3):278-287.
Sarmiento L, Llambí LD, Escalona A, Marquez N. Vegetation patterns, regeneration rates and divergence in an old-field succession of the high tropical Andes. Plant Ecol. 2003; 166:63-74.
Sarmiento L, Llambí LD. Regeneración del páramo luego de un disturbio agrícola: una síntesis de 20 años de investigaciones en sistemas con descansos largos en la Cordillera de Mérida. En Herrera F, Herrera I, editors. La Restauración Ecológica en Venezuela: fundamentos y experiencias. Caracas: Ediciones Instituto Venezolano de Investigaciones Científicas; 2011. p. 123-148.
Segovia-Salgado MA, Quijia-Lamiña P. Citogeografía de Cuatro Especies de Polylepis (Rosaceae) en el Ecuador: información relevante para el manejo y conservación de los bosques Andinos. En Cuesta F, Sevink J, Llambí LD, De Bièvre B, Posner J, editors. Avances en investigación para la conservación de los páramos andinos. Quito: CONDESAN; 2013. p. 467-486.
Shannon CE, Weaver W. The Mathematical Theory of Communication. Urbana, Illinois: University of Illinois Press; 1949. 120 p.
Suarez P, Chacón-Moreno E. Modelo espacial de distribución del ecotono bosque paramo en los Andes Venezolanos. Ubicación potencial y escenarios de cambio climático. Ecotrópicos. 2011;24(1): 3-25.
Troll C. The upper timberlines in different climatic zones. Arct Antarct Alp Res. 1973;5(3):A3-A18.
van der Hammen T. The Pleistocene changes in vegetation and climate in tropical South America. J Biogeogr. 1974;1(1):3-26. Doi: 10.2307/3038066.
van der Hammen T. Plan ambiental de la cuenca alta del Rio Bogotá análisis y orientaciones para el ordenamiento territorial. 1 ed. Bogotá: Corporación Autónoma Regional; 1998. 142 p.
van der Hammen T, Cleef AM. Development of the high Andean paramo flora and vegetation. In: Vuilleumier F, Monasterio M, editors. High altitude tropical biogeography. New York: Oxford University Press; 1986.p. 153-201.
Vareschi V. Ecología de la Vegetación Tropical. 1 ed. Caracas: Edición especial de la Sociedad Venezolana de Ciencias Naturales; 1992. 306 p.
van Steenis CGGJ. An attempt towards an explanation of the effect of mountain mass elevation. Proc K Ned Akad Wet C. 1961;64:435-442.
Vargas O. Estrategia para la restauración ecológica del bosque altoandino (El caso de la reserva forestal municipal de Cogua, Cundinamarca). 2 ed. Bogotá: Universidad Nacional de Colombia; 2008. 372 p.
Velasco-Linares P, Vargas O. Problemática de los Bosques Altoandinos. En: Vargas O, editor. Estrategias para la restauración ecológica del bosque altoandino (El caso de la Reserva Forestal Municipal de Cogua, Cundinamarca). Bogotá: Universidad Nacional de Colombia; 2008. p. 41-56.
von Humboldt A, Bondpland A. Essay on the Geography of Plants. Chicago: University of Chicago Press; 1807. 296 p.
White S. Grass páramo as hunter-gatherer landscape. The Holocene. 2013;23(6):898-915. Doi: 10.1177/0959683612471987.
Wille M, Hooghiemstra H, Hofstede R, Fehse J, Sevink J. Upper forest line reconstruction in a deforested area in northern Ecuador based on pollen and vegetation analysis. J Trop Ecol. 2002;18(3):409-440. Doi: 10.1017/S0266467402002286.
Yánez P. Caracterización florística en un sector de cambio páramo- selva nublada en el parque nacional Sierra Nevada, Venezuela. Revista Forestal Venezolana. 1998;42(1):51-62.
Young K, Leon B. Tree-line changes along the Andes: implications of spatial patterns and dynamics. Philos Trans R Soc Lond B Biol Sci. 2007;362(1478):263-272. Doi: 10.1098/rstb.2006.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Miguel Ángel Rendón Espinosa, Marius Bottin, Adriana Sanchez, Carlos Vargas, Lauren Raz, Adriana Corrales. (2024). Diversity of Mycorrhizal Types Along Altitudinal Gradients in the Tropical Andes. Global Ecology and Biogeography, 33(12) https://doi.org/10.1111/geb.13923.
2. Alberto Arzac, Luis D. Llambí, Raphael Dulhoste, José M. Olano, Eulogio Chacón-Moreno. (2019). Modelling the effect of temperature changes on plant life-form distribution across a treeline ecotone in the tropical Andes. Plant Ecology & Diversity, 12(6), p.619. https://doi.org/10.1080/17550874.2019.1655108.
3. Carlos Quiroz Dahik, Franklin Marín, Ruth Arias, Patricio Crespo, Michael Weber, Ximena Palomeque. (2019). Comparison of Natural Regeneration in Natural Grassland and Pine Plantations across an Elevational Gradient in the Páramo Ecosystem of Southern Ecuador. Forests, 10(9), p.745. https://doi.org/10.3390/f10090745.
4. Carolina Ramos‐Montaño, N. Milena Cárdenas‐Avella, Ledis V. Montenegro‐Rubiano, Angélica del Pilar Dallos, Rafael Andrés Rodríguez‐Sabogal. (2026). Water and Gas Exchange Responses of Five Related Blueberry Species (Ericaceae spp.) to the Dry Season in the High‐Andean Forest of the Colombian Eastern Cordillera. Plant-Environment Interactions, 7(4) https://doi.org/10.1002/pei3.70177.
5. Liz Alejandra Ávila Rodríguez, Paola Marcela Triviño. (2019). Descripción del hábitat de cuatro especies del género Lymanopoda (Lepidoptera: Nymphalidae: Satyrinae) en áreas de páramo de Colombia. Acta Biológica Colombiana, 24(1), p.125. https://doi.org/10.15446/abc.v24n1.70285.
6. Marcial Quiroga-Carmona. (2023). Exploring the effects of the quaternary glacial–interglacial cycles on the geographic distributions of tropical Andean rodents: species in the genus Aepeomys Thomas, 1898 (Thomasomyini: Sigmodontinae: Cricetidae) as a case study . Studies on Neotropical Fauna and Environment, 58(2), p.283. https://doi.org/10.1080/01650521.2021.1948654.
7. César I. Carvajal Hernández, Jorge Antonio Gómez Díaz, Alma P. Bautista-Bello, Thorsten Krömer. (2020). Encyclopedia of the World's Biomes. , p.79. https://doi.org/10.1016/B978-0-12-409548-9.11752-X.
8. Fermin Rada, Aura Azócar, Carlos García-Núñez. (2019). Plant functional diversity in tropical Andean páramos. Plant Ecology & Diversity, 12(6), p.539. https://doi.org/10.1080/17550874.2019.1674396.
9. Jesús Mavárez, Sylvain Bézy, Thomas Goeury, Angel Fernández, Serge Aubert. (2019). Current and future distributions of Espeletiinae (Asteraceae) in the Venezuelan Andes based on statistical downscaling of climatic variables and niche modelling. Plant Ecology & Diversity, 12(6), p.633. https://doi.org/10.1080/17550874.2018.1549599.
10. Lohengrin A. Cavieres, Luis Daniel Llambí, Fabien Anthelme, Robert Hofstede, Mary T.K. Arroyo. (2025). High-Andean Vegetation Under Environmental Change: A Continental Synthesis. Annual Review of Environment and Resources , 50(1), p.219. https://doi.org/10.1146/annurev-environ-111523-101920.
11. Luis Guillermo Acosta-Vargas, Dagoberto Arias-Aguilar, Juan Carlos Valverde. (2024). CARACTERIZACIÓN FLORÍSTICA DEL BOSQUE MONTANO Y SUBPÁRAMO DEL VOLCÁN IRAZÚ, COSTA RICA. Acta Biológica Colombiana, 29(1), p.32. https://doi.org/10.15446/abc.v29n1.97088.
12. Vanessa Correa Agudelo, María José Sanín, Dino Tuberquia. (2022). Transición bosque altoandino – páramo: composición, estructura y fisonomía de la vegetación en la Cordillera Central de Colombia. Caldasia, 45(1), p.174. https://doi.org/10.15446/caldasia.v45n1.102305.
13. Alexandra Buitrago‐Guacaneme, Carlos Molineri, Luciana Cristóbal, Daniel Andrés Dos Santos. (2022). The inter‐forest line could be the master key to track biocoenotic effects of climate change in a subtropical forest. Biotropica, 54(1), p.57. https://doi.org/10.1111/btp.13026.
14. Lirey A. Ramírez, Luis D. Llambí, Carmen J. Azocar, Manuel Fernandez, J. Eloy Torres, Maaike Y. Bader. (2022). Patterns in climate and seedling establishment at a dry tropical treeline. Plant Ecology, 223(9), p.1047. https://doi.org/10.1007/s11258-022-01257-2.
15. Carlos Quiroz Dahik, Patricio Crespo, Bernd Stimm, Reinhard Mosandl, Jorge Cueva, Patrick Hildebrandt, Michael Weber. (2021). Impacts of pine plantations on carbon stocks of páramo sites in southern Ecuador. Carbon Balance and Management, 16(1) https://doi.org/10.1186/s13021-021-00168-5.
16. Luis Daniel Llambí, Fermín Rada. (2019). Ecological research in the tropical alpine ecosystems of the Venezuelan páramo: past, present and future. Plant Ecology & Diversity, 12(6), p.519. https://doi.org/10.1080/17550874.2019.1680762.
17. Eulogio Chacón-Moreno, Mayanín Rodríguez-Morales, Daniela Paredes, Patricia Suárez del Moral, Anderson Albarrán. (2021). Impacts of Global Change on the Spatial Dynamics of Treeline in Venezuelan Andes. Frontiers in Ecology and Evolution, 9 https://doi.org/10.3389/fevo.2021.615223.
18. Maria Pia Calcagno-Pissarelli, Jorge Luis Avila-Nuñez, Miguel E. Alonso-Amelot. (2023). Neotropical Gradients and Their Analysis. , p.37. https://doi.org/10.1007/978-3-031-22848-3_2.
19. Gwendolyn Peyre, Henrik Balslev, Xavier Font. (2018). Phytoregionalisation of the Andean páramo. PeerJ, 6, p.e4786. https://doi.org/10.7717/peerj.4786.
20. Yisela Fernanda Solarte Erazo, María Elena Solarte Cruz, Javier Gulías León, Jaume Flexas. (2025). Rasgos fisiológicos de plantas endémicas y no endémicas presentes en el ecosistema de páramo. Boletín Científico Centro de Museos Museo de Historia Natural, 29(1), p.13. https://doi.org/10.17151/bccm.2025.29.1.1.
21. Clara Morales-Rozo, Ian S. Acuña-Rodríguez, Juan C. Benavides, Víctor J. Rincón-Parra, Mauricio Diazgranados. (2024). Prioritization of important plant areas for conservation of frailejones (Espeletiinae, Asteraceae) in the Northern Andes. Biodiversity and Conservation, 33(14), p.4073. https://doi.org/10.1007/s10531-024-02939-1.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2015 Acta Biológica Colombiana

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
1. La aceptación de manuscritos por parte de la revista implicará, además de su edición electrónica de acceso abierto bajo licencia Attribution-NonCommercial-ShareAlike 4.0 (CC BY NC SA), la inclusión y difusión del texto completo a través del repositorio institucional de la Universidad Nacional de Colombia y en todas aquellas bases de datos especializadas que el editor considere adecuadas para su indización con miras a incrementar la visibilidad de la revista.
2. Acta Biológica Colombiana permite a los autores archivar, descargar y compartir, la versión final publicada, así como las versiones pre-print y post-print incluyendo un encabezado con la referencia bibliográfica del articulo publicado.
3. Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
4. Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos institucionales, en su página web o en redes sociales cientificas como Academia, Researchgate; Mendelay) lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).












