



Published
Mechanisms of resistance to Neoleucinodes elegantalis (Guenée) in wild germplasm of the genus Solanum
Keywords:
tomatoes, Solanum habrochaites, defense mechanisms, trichomes. (es)Downloads
Several phytosanitary problems affect tomato (Solanum lycopersicum) crops, one of the most important being the tomato fruit borer, which has caused losses of up to 70% in areas of the Valle del Cauca department (Colombia). To find resistance mechanisms to this pest, plants of three wild introductions of Solanum (PI134417, PI134418, LA1264) and the commercial cultivar Unapal-Maravilla were planted in a screenhouse. A completely randomized design was used with eight replicates. Five releases of N. elegantalis were carried out and the response was evaluated by the antixenosis or no preference test. Five racemes of each introduction were also suspended from the top of a field cage (1.8 x 1.5 x 1.5 m) to evaluate their response to six releases of the pest. For the two experiments, both an analysis of variance and analysis of means were performed. The average oviposition per plant was significantly higher in Unapal-Maravilla as compared with the wild introductions, and the average number of eggs per fruit was significantly lower in LA1264. The insects laid more eggs on the fruit surface (>70%). Glandular trichomes types I, IV and VI found in the wild introductions and their associated chemicals had an antibiotic and antixenotic effect on N. elegantalis.
PLANT BREEDING, GENETIC RESOURCES & MOLECULAR BIOLOGY
Mechanisms of resistance to Neoleucinodes elegantalis (Guenée) in wild germplasm of the genus Solanum
Mecanismos de la resistencia a Neoleucinodes elegantalis (Guenée) en germoplasma silvestre dénero Solanum
Nelson Enrique Casas-Leal1 Franco Alirio Vallejo-Cabrera1 and Edgar Iván Estrada-Salazar1
1Faculty of Agricultural Sciences, Universidad Nacional de Colombia. Palmira (Colombia). favallejoc@unal.edu.coReceived for publication: 8 February, 2013. Accepted for publication: 5 June, 2013.
ABSTRACT
Several phytosanitary problems affect tomato (Solanum lycopersicum) crops, one of the most important being the tomato fruit borer, which has caused losses of up to 70% in areas of the Valle del Cauca department (Colombia). To find resistance mechanisms to this pest, plants of three wild introductions of Solanum (PI134417, PI134418, LA1264) and the commercial cultivar Unapal-Maravilla were planted in a screenhouse. A completely randomized design was used with eight replicates. Five releases of N. elegantalis were carried out and the response was evaluated by the antixenosis or no preference test. Five racemes of each introduction were also suspended from the top of a field cage (1.8 x 1.5 x 1.5 m) to evaluate their response to six releases of the pest. For the two experiments, both an analysis of variance and analysis of means were performed. The average oviposition per plant was significantly higher in Unapal-Maravilla as compared with the wild introductions, and the average number of eggs per fruit was significantly lower in LA1264. The insects laid more eggs on the fruit surface (>70%). Glandular trichomes types I, IV and VI found in the wild introductions and their associated chemicals had an antibiotic and antixenotic effect on N. elegantalis.
Key words: tomatoes, Solanum habrochaites, defense mechanisms, trichomes.
RESUMEN
Diversos problemas fitosanitarios afectan el cultivo de tomate (Solanum lycopersicum), siendo el pasador del fruto uno de los más importantes, causando pérdidas de hasta el 70% en zonas del departamento del Valle del Cauca (Colombia). Para encontrar mecanismos de resistencia a esta plaga, se sembraron plantas de tres introducciones silvestres de Solanum (PI134417, PI134418, LA1264) y un cultivar comercial Unapal-Maravilla en una casa de malla circular, utilizando un diseño completamente al azar con ocho repeticiones. Se realizaron cinco liberaciones de N. elegantalis y la respuesta se evaluó mediante la prueba de antixenosis o de no preferencia. Adicionalmente, cinco racimos de cada introducción fueron suspendidos de la parte superior de una jaula (1,8 x 1,5 x 1,5 m) para evaluar su respuesta a seis liberaciones de la plaga. Ambos ensayos se sometieron a análisis de varianza y análisis de medias. Unapal- Maravilla presentó un promedio de ovoposición por planta significativamente mayor que las introducciones silvestres, y LA1264 presentó un número promedio de huevos por fruto significativamente menor. Los insectos depositaron más huevos en la superficie del fruto (>70%). Los tricomas glandulares tipos I, IV y VI presentes en introducciones silvestres y sus sustancias químicas asociadas ejercieron un efecto antixenótico y antibiótico en N. elegantalis.
Palabras clave: tomate, Solanum habrochaites, mecanismos de defensa, tricomas.
Introduction
The tomato (Solanum lycopersicum L.) is one of the most important vegetables in Colombia and the world, in terms of fresh consumption and industrial purposes. One of the main problems affecting this crop, however, is its high susceptibility to pests. In Colombia, four species of insect pests severely limit tomato production: fruit borer Neoleucinodes elegantalis, citrus gall midge Prodiplosis longifila, whitefly Bemisia tabaci and tomato leaf miner Tuta absoluta. Crop losses of up to 70%, attributable to N. elegantalis, have been reported in the department of Valle del Cauca and, as this pest remains inside the fruit throughout its larval stage, it is very difficult to control. Furthermore, all tomato cultivars used in Colombia are susceptible to this species. The problem is so serious that for production to be economically viable, producers must resort to extreme control practices such as the indiscriminate use of highly toxic, residual pesticides, which greatly increases production costs and poses risks not only for the environment but also for the health of those using these products and consumers.
The development of cultivars genetically resistant to this pest is a realistic, efficient and long-term management alternative. To date, however, few studies have been conducted in Colombia that can serve as a basis to develop tomato cultivars resistant to N. elegantalis and, as a result, the information available is insufficient to begin the selection of resistant tomato genotypes. This study therefore aimed to study the response of insect resistance mechanisms present in three wild introductions of the genus Solanum, for subsequent breeding into cultivated tomatoes.
Materials and methods
Location
The study was carried out at the Experimental Center of the Universidad Nacional de Colombia-Palmira campus (CEUN P), located in the municipality of Candelaria, department of Valle del Cauca (03°25'25.3'' N, 76°25'47.8'' W) at an altitude of 972 m a.s.l. and with an average temperature of 26°C and average relative humidity of 76%.
Evaluation under screenhouse conditions
The wild introductions Solanum habrochaites var. glabratum PI134417 and PI134418 and introduction S. habrochaites var. typicum LA1264, all identified as highly resistant, were used, as well as the Solanum lycopersicum commercial cultivar Unapal-Maravilla, identified as very susceptible based on assessments made by Restrepo et al. (2003).
A completely randomized design was used with the four introductions, eight plants per introduction. Five racemes were evaluated per plant. The plants were grown in 25-L plastic buckets, inside a circular screenhouse, with a 9 m diameter and 2.5 m in height, covered with muslin cloth. Plants of each introduction were randomly distributed in a circle, equidistant from the center of the screenhouse. Every week, plants were removed from the screenhouse for 8 h to facilitate pollination of the wild introductions by insects. The racemes formed were covered with muslin-cloth bags. One two-stemmed plant was used per bucket, and no insecticides were applied after the onset of fructification to avoid altering insect behavior.
Five releases of adult N. elegantalis were carried out according to the procedure described by Casas and Estrada (2005) each consisting of three females and one male, in the center of the screenhouse, to allow free choice of host plants. These releases were carried out when the diameter was greater than 1 cm for half of the raceme fruits of all plants of the four introductions.
Five days after each release, the racemes were carefully cut to avoid the release of the eggs laid by the insects, stored in a Bellotti-type containment tray, and placed in the laboratory to carry out measurements aided by a microscope. The variables evaluated were: number of oviposited fruits per plant, number of eggs per fruit, number of eggs per plant, number of eggs per raceme, location of laid eggs and presence of trichomes on the fruit.
The data corresponding to the variable oviposited fruit per plant were transformed using arcsine and those of variables number of eggs per fruit and number of eggs per plant, using. Descriptive analysis was performed as well as analysis of variance using the GLM procedure of SAS ®. Means comparison was performed using the LSD test, with a significance level of P≤0.05.
Evaluation in field cage conditions
To confirm the results obtained in the screenhouse, further testing was conducted in a field cage measuring 1.5 x 1.5 x 1.8 m, covered with muslin cloth. Fruit racemes of uniform development and number of fruits of each of the four introductions were hung inside the cage (Fig.1).

A completely randomized design was used with the same four introductions, six racemes per introduction. Six releases of adult N. elegantalis were carried out, each with 12 females and three males. Racemes were evaluated 5 d after each release following the same procedure as in the screenhouse. The racemes were classified by position on the plant as follows: level A (low racemes: 1, 2 and 3) and level B (middle or upper racemes: 4 and 5).
The following variables were measured: oviposition per raceme, average number of eggs per raceme, total number of eggs per raceme and location of laid eggs. The data corresponding to the variable oviposition per raceme were transformed using arcsine and that of the variables average number of eggs per raceme and total eggs per raceme, using. Analysis of variance was performed using the GLM procedure of SAS ®, and mean comparison was performed using the LSD test, with a significance level of P≤0.05.
Results and discussion
Evaluation under screenhouse conditions
Number of oviposited fruit per plant
Significant differences were detected between introductions regarding number of oviposited fruits per plant. Unapal- Maravilla presented the highest oviposition per plant as compared with PI134417, PI134418 and LA1264. Likewise, LA1264 recorded an average significantly lower than those of Unapal-Maravilla, PI134417 and PI134418. There were no significant differences between the introductions of S. habrochaites var. glabratum PI134417 and PI134418 (Tab.1).
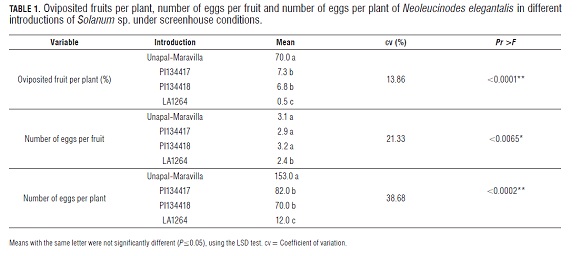
Based on the number of oviposited fruits per plant, the high susceptibility of cultivated Unapal-Maravilla tomatoes and the high resistance of wild introductions to N. elegantalis were confirmed. In the case of Unapal-Maravilla, 70.0% of the fruits were oviposited, compared with only 7.3% of the fruits of S. habrochaites var. glabratum PI134417 and 6.8% of those of PI134418. Introduction S. habrochaites var. typicum LA1264 presented the lowest oviposition value, with only 0.5% of its fruits presenting eggs. The extremely low oviposition of N. elegantalis in wild introductions, as compared with cultivated tomatoes, suggests that they have an antixenotic effect on the tomato fruit borer.
Number of eggs per fruit
Significant differences were found between the introductions regarding the number of eggs per fruit. LA1264 presented a significantly lower value than Unapal-Maravilla, PI134417 and PI134418. There were no significant differences between Unapal-Maravilla, PI134417 and PI134418 (Tab.1). It should be taken into account that the number of eggs per fruit is conditioned by the natural reproductive activity of the insect, which is impossible to control.
Number of eggs per plant
The number of eggs per plant varied significantly between introductions. Unapal-Maravilla recorded a value significantly higher than the averages of PI134417, PI134418 and LA1264. Similarly, LA1264 presented a value significantly lower than those of Unapal-Maravilla, PI134417 and PI134418. There were no significant differences between PI134417 and PI134418 (Tab.1).
Kennedy (2003) reported that S. habrochaites var. glabratum and S. habrochaites var. typicum have high levels of resistance to a large number of arthropod species. The results of this study verify the resistance of wild Solanum sp. introductions to the tomato fruit borer N. elegantalis.
Location of laid eggs
The location of N. elegantalis eggs laid on the four Solanum introductions is shown in both Fig.2 and Tab.2, with 69.3% of the eggs being laid on the fruit and the remaining 30.7% on the calyx and peduncle.
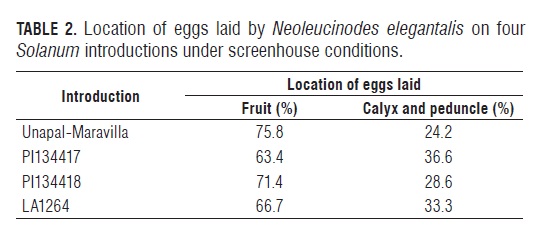

These results show that N. elegantalis prefers to lay eggs on the fruit surface rather than other structures such as the calyx and peduncle, regardless of the type of tomato (cultivated or wild). In a few cases, eggs were found in small leaflets connected to the raceme, in raceme rachis and even in leaf folioles near the racemes.
Number of eggs per raceme. Fig.3 shows the percentage of oviposition in the first five racemes, with susceptible introduction Unapal-Maravilla presenting the highest number of eggs. The apparent increase in eggs laid on racemes 4 and 5 of wild introductions can be attributed to the low availability of racemes and fruits of the susceptible introduction.

A significant increase in oviposition per raceme was observed in wild introductions, especially in PI134417 and PI134418, increasing from 3.5% in the lower racemes to 12% in middle racemes. In the case of LA1264, the difference in raceme position was not significant, oviposition being below 1% in both.
When only a low number of fruits was available on the susceptible introduction Unapal-Maravilla, the insects were forced to ovipositon the fruits of wild introductions, although these were not the preferred host plants. This situation can be attributed to the instinct of survival and selfpreservation of the species in conditions of confinement. Restrepo et al. (2007) reported the presence of several eggs on fruits of wild introductions under field conditions, thus converting them into host plants of this insect; the fruits, however, showed no damage.
While there is evidence of the presence of different mechanisms that allow wild introductions to defend themselves against different pests, ranging from physical (different types of trichomes) to chemical barriers (toxic substances in the trichomes and release of volatile repellents), these did not prevent adult female N. elegantalis from ovipositing on wild introductions under controlled conditions, especially on S. habrochaites var. glabratum PI134417 and PI134418, although to a much lesser degree than on cultivated tomato Unapal-Maravilla.
Van Schie et al. (2007) stated that S. lycopersicum plants produce and liberate a variety of volatile organic compounds, mainly terpenes present in their trichomes. Despite this, the present study demonstrated that, under controlled conditions, the cultivated tomato Unapal-Maravilla was highly susceptible to N. elegantalis attack.
Presence of trichomes in fruit and oviposition
Most of the eggs laid on the wild introductions attached to type 1 trichomes of the fruit (Fig.4). This behavior probably resulted from the attempt to avoid the fruit surface, where type IV and VI trichomes are found, largely responsible for the high levels of resistance in these introductions as mentioned by Simmons and Gurr (2005), Kennedy (2003), De Souza et al. (2002) and Aragão et al. (2000).

These results validate the studies conducted by Kennedy (2003) and Simmons et al. (2004), who put forth that both glandular and non-glandular trichomes are essential plant defense components of the genus Solanum.
The presence of eggs attached to type 1 trichomes of the fruit in wild Solanum introductions suggests an antixenotic effect on adult N. elegantalis because it alters the insect's reproductive behavior when compared with its behavior on the susceptible introduction Unapal-Maravilla, where oviposition occurs on the fruit surface. The larvae hatching from eggs laid on this type of trichomes, after descending and reaching the fruit surface where type IV and VI trichomes are found, died quickly and very near the chorions (Fig.5A). Some larvae were also observed trying to escape from the fruit surface, again ascending the type I trichomes, where they died shortly afterwards (Fig.5B). These reactions of neonate larvae of N. elegantalis suggest a possible antibiotic effect.
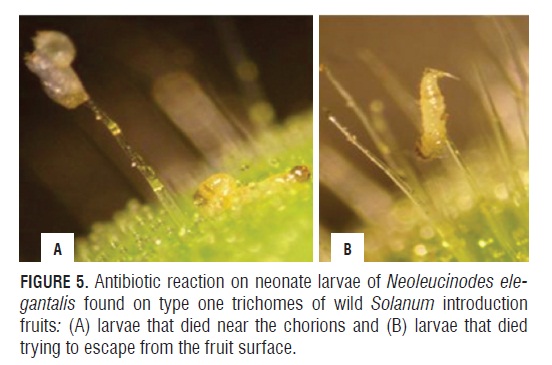
Regarding the antibiotic and antixenotic effect of type I, IV and VI glandular trichomes as well as non-glandular trichomes present in wild introductions, Aragão et al. (2000) asserted that non-glandular trichomes can act directly on the insects, affecting oviposition, feeding, locomotion and behavior by density and size. Simmons and Gurr (2005) stated that type VI glandular trichomes of S. habrochaites var. typicum contain several sesquiterpenes, including zingiberene, which is predominant in this type of trichome. Kennedy (2003) indicated that the leaves of S. habrochaites var. glabratum PI134417 were lethal to multiple pest species due to the presence of toxic substances such as methylketones (2-tridecanone and 2-undecanone) in type VI glandular trichomes. The ketones make up 90% of the content of the gland head of type VI glandular trichomes in PI134417.
Eggs laid directly on the fruit surface were observed in the wild introductions PI134417, PI134418 and LA1264, but in smaller amounts. Larvae hatched from these eggs died a few millimeters from the chorions (Fig.6), indicating an alteration in insect biology related to a possible antibiotic effect on the first development stage of the borer. Type IV and VI trichomes of the three wild introductions PI134417, PI134418 and LA1264 trapped the neonate larvae of N. elegantalis, limiting their movements and causing their death by action of the secondary metabolites they contain (Fig.6). Simmons and Gurr (2005) indicated that when the contents of glandular trichomes are released and come into contact with the pest, they can cause irritation, trap the larvae and even kill them.

Peter and Shanower (1998) found that tomato trichomes produce large amounts of 2-tridecanone, 2-undecanoneanda viscous mixtures of acyl sugars. They also release volatile chemicals such as (E)-betafarnesene and nepetalactone. Aragão et al. (2000) stated that glandular trichomes secrete terpenes, alkaloids, phenolics and other substances that act as olfactory or taste repellents. Similarly, Simmons and Gurr (2005) indicated that the exudates of these trichomes can deter or repel pests, suggesting an important antixenotic effect in response to chemicals present in trichome exudates, such as the acyl sugars in type IV trichomes that have a deterrent effect on insects.
Several of the larvae that were able to initially over come the barriers of type I, IV and VI glandular trichomes as well as the effect of their chemical components tried to penetrate the fruit, but died in the attempt (Fig.7A). Living larvae were observed penetrating the fruits (Fig.7B), but soon after penetration were found dead inside the fruit (Fig.7C), slightly damaging the affected fruits, mainly the mesocarp tissue. These reactions on insect biology suggest a possible antibiotic effecton N. elegantalis.

Simmons and Gurr (2005) stated that antibiosisis is probably conferred by the chemical components of trichome exudates. Two types of methylketones (2-tridecanone and 2-undecanone) are present in type VI trichomes of S. habrochaites var. glabratum, and are associated with many adverse effects on different Lepidopterans and Hemipterans. They also indicated that the trichomes associated with adverse effects on pests (mortality, survival and entrapment) are type IV and VI glandular trichomes present in the wild introductions used in this study.
The adverse reactions observed in insect reproductive behavior and biology demonstrate a possible antibiotic and antixenotic effect on N. elegantalis, mainly attributed to the presence of type IV and VI glandular trichomes in fruits of wild introductions PI134417, PI134418 and LA1264. In the case of Unapal-Maravilla, the female insects laid most of the eggs on the fruit surface and, to a lesser extent, on the calyx. Eggss howed no alteration in their development and larvae easily penetrated the fruit and developed normally.
The fruits of the wild introductions PI134417, PI134418 and LA1264 showed type I, IV and VI glandular trichomes (Fig.8), which are the main defense against different pests, including the tomato fruit borer N. elegantalis. Cultivated tomatoes present type I, III, V, VI and VII glandular trichomes on the fruit, young stems, shoot sand flower buds (Luckwill, 1943). They also contain steroidal glycoalkaloids, such as a-tomatine, that act as toxic defense compounds. These are found throughout the plant, including fruits and roots. However, despite having a great diversity of trichomes, Unapal-Maravilla is highly susceptible to N. elegantalis attack. Some researchers, such as Williams et al. (1980), have indicated that the glandular trichomes of cultivated tomatoes have concentrations of 2-tridecanone and 2-undecanone 72 times lower than those found in wild introductions of S. habrochaites.

The different reactions observed in neonate larvae of N. elegantalis suggest that the type I, IV and VI glandular trichomes present in the wild introductions exert a strong antibiotic effect on neonate larvae of this pest, causing 100% mortality because no larvae of advanced instars or living larvae were found after two days of observation.
Evaluation in field cage conditions
The analysis of variance for the variables percentage of oviposition, average number of eggs per raceme and number of eggs per raceme showed significant differences among the introductions. Values for Unapal-Maravilla were significantly higher than those for PI134417, PI134418 and LA1264. No significant differences were observed between the values of wild introductions PI134417, PI134418 and LA1264 (Tab.3).

Tab.3 shows the susceptibility of the cultivated tomato Unapal-Maravilla and the resistance of the wild introductions to the tomato fruit borer N. elegantalis. In Unapal-Maravilla, 36.3% of the fruits were oviposited as compared with only 6.9% in S. habrochaites var. glabratum PI134417 and 3.0% in PI134418. In the case of S. habrochaites var. typicum LA1264, 4.1% of its fruit demonstrated oviposition. The notable difference in percentage of oviposition in wild introductions as compared with cultivated tomatoes suggests an antixenotic effect of these introductions, following a trend similar to that observed under the screenhouse conditions.
As observed in the screenhouse trial (Tab.1), the cultivated tomato Unapal-Maravilla differed significantly from the wild introductions regarding percentage of oviposited fruit as well as total number of eggs laid on 10 racemes, with 76% of the oviposition occurring on the fruit and the remaining 24% on the calyx and peduncle. These results are similar to those observed under the screenhouse conditions where oviposition on the fruit was 69.3% and 30.7% on the other plant structures (Tab.4).
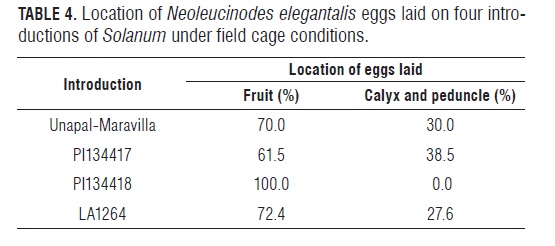
Just as in the screenhouse, thefemale insects preferred to oviposit onthe fruit surfacerather than on otherstructuressuch asthe calyxand peduncle.
In introduction PI134418, all oviposited eggs were on the fruit, whereas oviposition on the fruit was 70.0% in Unapal- Maravilla, 61.5% in PI134417and 72.4% in LA124. No eggs of the tomato fruit borer N. elegantalis were laid on the shoots accompanying the racemes, probably because of the rapid deterioration of plant material promoted by tissue dehydration. Despite this fact, the fruits of the evaluated racemes had a good appearance when evaluations were done 5 dafter cutting, with racemes looking similar to those of whole plants in the field.
The reactions on the reproductive behavior and biology of the neonate larvae of this pest observed in the field cage conditions were similar to those observed in the screenhouse trials, confirming the apparent antibiotic and antixenotic effects on N. elegantalis, mainly due to the presence of type IV and VI glandular trichomes in the fruits of the wild Solanum introductions PI134417, PI134418 and LA1264.
Literature cited
Aragão, C.A., D.B. França, F. Rodrigo, and B.R. Gandolfi. 2000. Tricomas foliares em tomateiro com teores contrastantes do aleloquímico 2-tridecanona. Sci. Agric. 57(4), 813-816.
Casas, N.E. and E.I. Estrada. 2005. Estudios preliminares sobre la utilización de ultrasonido en el control del pasador del fruto Neoleucinodes elegantalis Guenée en el cultivo del tomate Lycopersicum esculentum Mill. M.Sc. thesis. Faculty of Agricultural Sciences, Universidad Nacional de Colombia, Palmira, Colombia.
De Souza, C., A. Viana-Bailez, and J. Blackmer. 2002. Toxicity of tomato allelochemicals to eggs and neonato larvae of Neoleucinodes elegantalis (Guenée) Service (Lep: Crambidae). p. 295. In: Abstracts, Entomology International Congress Proceedings. Agricultural Research Service, Washington DC.
Kennedy, G.G. 2003. Tomato, pests, parasitoids, and predators: tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol. 48, 51-72.
Luckwill, L.C. 1943. The genus Lycopersicon: An historical, biological,g and taxonomic survey of the wild and cultivated tomatoes. Aberd. Univ. Stud. 120, 1-44.
Peter, A.J. and T.G. Shanower. 1998. Plant glandular trichomes chemical factories with many potential uses. Resonance 3, 41-45.
Restrepo, E.F., F.A. Vallejo, and M. Lobo. 2003. Estudios básicos para iniciar la producción de cultivares de tomate Solanum lycopersicum L. con resistencia al pasador del fruto Neoleucinodes elegantalis (guenée). Ph.D. thesis. Faculty of Agricultural Sciences, Universidad Nacional de Colombia, Palmira, Colombia.
Restrepo, E., F.A. Vallejo, and M. Lobo. 2007. Producción de poblaciones segregantes resistentes al pasador del fruto a partir de cruzamientos entre tomate y accesiones silvestres de Lycopersicum spp. Acta Agron. 56(1), 1-6.
Simmons, A.T., G.M. Gurr, D. McGrath, P.M. Martin, and H.I. Nicol. 2004. Entrapment of Helicoverpa armigera (Hübner) (Lep: Noctuidae) on glandular trichomes of Lycopersicon species. Aust. J. Entomol. 43, 196-200.
Simmons, A.T. and G.M. Gurr. 2005. Trichomes of Lycopersicon species and their hybrids: Effects on pests and natural enemies. Agric. For. Entomol. 7(4), 265-276.
Van Schie, C.C., M.A. Haring, and R.C. Schuurink. 2007. Tomato linalool synthase is induced in trichomes by jasmonic acid. Plant Mol. Biol. 64, 251-263.
Williams, W.G., G.G. Kennedy, E.T. Yamamoto, J.D. Thacker, and J. Bordner. 1980. 2-tridecanone: a naturally occurring insecticide from the wild tomato Lycopersicon hirsutum f. glabratum. Science 207(4433), 888-889.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
Article abstract page views
Downloads
License
Copyright (c) 2013 Agronomía Colombiana

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
© Centro Editorial de la Facultad de Ciencias Agrarias, Universidad Nacional de Colombia
Reproduction and quotation of material appearing in the journal is authorized provided the following are explicitly indicated: journal name, author(s) name, year, volume, issue and pages of the source. The ideas and observations recorded by the authors are their own and do not necessarily represent the views and policies of the Universidad Nacional de Colombia. Mention of products or commercial firms in the journal does not constitute a recommendation or endorsement on the part of the Universidad Nacional de Colombia; furthermore, the use of such products should comply with the product label recommendations.
The Creative Commons license used by Agronomia Colombiana journal is: Attribution - NonCommercial - ShareAlike (by-nc-sa)

Agronomia Colombiana by Centro Editorial of Facultad de Ciencias Agrarias, Universidad Nacional de Colombia is licensed under a Creative Commons Reconocimiento-NoComercial-CompartirIgual 4.0 Internacional License.
Creado a partir de la obra en http://revistas.unal.edu.co/index.php/agrocol/.
















