



Publicado
2005-07-01
El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)
Con el objetivo de determinar la posibilidad de obtener tejidos sanos a partir de plantas infectadas con ASBVd como una estrategia para recuperar clones de alto valor agronómico, cultivos embriogénicos de aguacate fueron inducidos a partir del nucelo de tres cultivares de aguacate infectados con el viroide de la mancha del sol (ASBVd) en un medio de cultivo con las sales mayores B5, suplementado con las sales menores de Murashige y Skoog (MS) y 0.41 uM de picloram, y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100), sucrosa (30,000) y TC agar (8,000). Los cultivos embriogénicos inducidos fueron mantenidos en medio semisólido MS suplementado con 0.41 uM de picloram y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100), sucrosa (45,000) y TC agar (8,000) y en medio líquido MS modificado con 15 mM NH4NO3 y 30 mM KNO3, sales menores de MS, picloram 0.41 uM y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100) y sucrosa (45,000). La indexación usando RT-PCR un año después de inducidos los tejidos detectó bandas amplificadas en los electroferogramas y la clonación comprobó la presencia de 4 clones con mas de un 97% de similitud con la variante J02020 de ASBVd; con la secuencia se corroboró la existencia de las variantes CF3, CF8, CF13. Estos resultados demuestran que aunque ASBVd no es eliminado mediante el uso de embriogénesis somática, la aplicación de esta tecnología permite inducir, proliferar y mantener por largo tiempo tejidos infectados para el estudio de este patógeno.
|
|
||
|
Rev. Colomb. Biotecnol. Vol. VII No. 2 Diciembre 2005 10-18
|
||
|
|
||
|
El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)
Avocado sunblotch viroid (ASBVd) is persistent in nucellar cultures (Persea americana Mill.)
Isidro E. Suárez*, RichardE. Litz**, Raymond J. Schnell***, David N. Kuhn****
RESUMEN
Con el objetivo de determinar la posibilidad de obtener tejidos sanos a partir de plantas infectadas con ASBVd como una estrategia para recuperar clones de alto valor agronómico, cultivos embriogénicos de aguacate fueron inducidos a partir del nucelo de tres cultivares de aguacate infectados con el viroide de la mancha del sol (ASBVd) en un medio de cultivo con las sales mayores B5, suplementado con las sales menores de Murashige y Skoog (MS) y 0.41 uM de picloram, y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100), sucrosa (30,000) y TC agar (8,000). Los cultivos embriogénicos inducidos fueron mantenidos en medio semisólido MS suplementado con 0.41 uM de picloram y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100), sucrosa (45,000) y TC agar (8,000) y en medio líquido MS modificado con 15 mM NH4NO3 y 30 mM KNO3, sales menores de MS, picloram 0.41 uM y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100) y sucrosa (45,000). La indexación usando RT-PCR un año después de inducidos los tejidos detectó bandas amplificadas en los electroferogramas y la clonación comprobó la presencia de 4 clones con mas de un 97% de similitud con la variante J02020 de ASBVd; con la secuencia se corroboró la existencia de las variantes CF3, CF8, CF13. Estos resultados demuestran que aunque ASBVd no es eliminado mediante el uso de embriogénesis somática, la aplicación de esta tecnología permite inducir, proliferar y mantener por largo tiempo tejidos infectados para el estudio de este patógeno.
Palabras clave: ASBVd, embriogénesis somática, RT-PCR, electroforesis capilar, secuenciamiento
ABSTRACT
Avocado embryogenic cultures were induced from the nucelli of three ASBVd (avocado sunblotch viroid)-infected cultivars to see whether healthy tissues could be obtained from infected plants for providing clones having added agronomic value. Embryogenic cultures were induced from the nucelli of three ASBVd-infected cultivars in B5 medium supplemented with Murashige and Skoog (MS) minor salts, 0.41 uM pycloram and (in mg/L-1) thiamine HCl (0.4), myo-inositol (100), sucrose (30,000) and TC agar (8,000). Embryogenic cultures were kept in semi-solid MS medium with 0.41 uM pycloram and (in mg/L-1) thiamine HCl (0.4), myo-inositol (100), sucrose (45,000) and TC agar (8,000) and in liquid MS3:1N medium supplemented with MS major salts with 15 mM NH4NO3 and 30 mM KNO3, MS minor salts, 0.41 uM pycloram and (in mg/L-1) thiamine HCl (0.4), myo-inositol (100) and sucrose (45,000). RT-PCR indexing showed that the embryogenic cultures remained ASBVd-infected one year after induction and ASBVd variants were isolated from the cultures. The results proved that ASBVd was not eliminated via nucellus culture, although it showed that somatic embryogenesis can be used as a practical approach to inducing, multiplying and maintaining ASBVd-infected material for extended periods of times for research purposes.
Key words: ASBVd, somatic embryogenesis, RT-PCR, capillary electrophoresis, sequencing.
|
||
|
|
||
|
* Ph. D. Departamento de Agronomía, Universidad de Córdoba. Carrera 6 No. 76-103. Montería, Colombia. Correo electrónico: isuarez@sinu.unicordoba.edu.co
Ph. D. University of Florida, Trec 18905 SW 280 St. Homestead, FL 33031. Ph. D. USDA-ARS, 13601 Old Cutler Road, Miami, FL 33158. Ph. D. Department of Biological Sciences, Florida International University, Miami, FL 33199.
Recibido: Junio 30 de 2004 Aceptado: Mayo 26 de 2005
|
||
|
|
||
|
10
|
||
|
|
||
|
|
|||
|
EL VIROIDE DE LA MANCHA DEL SOL (ASBVd) ES PERSISTENTE EN CULTIVOS NUCELARES
|
|||
|
|
|||
|
INTRODUCCIÓN
La mancha del sol es una enfermedad de importancia económica en el cultivo del aguacate (Persea americana Mill.) que se reportó por primera vez en California (Estados Unidos) y descrita inicialmente como un problema fisiológico (Coit, 1928). El agente causal es el viroide de la mancha del sol del aguacate (ASBVd) (Palukaitis et ál., 1971), el cual consiste de una molécula circular de ARN de 246-249 nucleó-tidos de tamaño (Symons, 1981) perteneciente a la familia Avsunviroidae (Flores et ál., 1998). ASBVd se replica en los cloroplastos usando riboenzimas que se encuentran inmersas en su propia secuencia (Navarro et ál., 1999; Bonfiglioli et ál., 1994).
La mancha del sol afecta por igual a todos los cultivares de aguacate, se transmite a través de semilla, partes vegetativas, polen y herramientas contaminadas, y la erradicación de las plantas afectadas es el único medio de control disponible para prevenir su diseminación (Wallace y Drake, 1962; Horne y Parker, 1931; Desjardins et ál., 1979; Desjardins et ál., 1980; Stevens y Piper, 1941). Evaluaciones de productividad han demostrado que las plantas de aguacate infectadas con ASBVd pueden presentar una reducción de 27.3% en el rendimiento total de fruta y producir un incremento del 52.7% en frutos con baja calidad por la presencia de manchas en el fruto (Da Gracaetál., 1983). De manera adicional, la amenaza de ASBVd se extiende a los bancos de germoplasma. Un programa generalizado de inde-xación realizado en el Repositorio Nacional de Germoplasma en Miami, basado en RT-PCR (Schnell et ál., 1997), demostró que un 20% del total de la colección de aguacate estaba infectada con ASBVd, lo cual propició la suspensión en la distribución de material de propagación a los productores y viveristas.
Aunque utilizando una tecnología diferente para la indexación de las plantas, el cultivo de tejido nucelar vía embriogénesis somática ha sido reportado como una forma de recuperar individuos sanos a partir de plantas adultas infectadas con virus y viroi-des(WeathersyCalavan, 1959; Ranganetál., 1968; Bitters et ál., 1972), y más recientemente, usando RT-PCR como método de indexación, el uso de termoterapia junto con el cultivo de meristemos ha permitido obtener plantas sanas a partir de árboles infectados (Postman y Hadidi, 1995; Howell et ál., 1998). Con base en lo anterior y aplicando las técnicas de embriogénesis somática en aguacate se ha
|
diseñado el presente trabajo que tiene como objetivo utilizar embriogénesis somática para inducir y proli-ferar tejidos embriogénicos a partir del nucelo de plantas infectadas con ASBVd en un intento por recuperar clones sanos a partir de cultivares contaminados con ASBVd.
MATERIALES Y MÉTODOS
Material vegetal
Frutos inmaduros (<0.5 cm diam.) de los cultivares 'Vero Beach' SE2, 'Vero Beach' 1 y 'Yama' 381 plantados en el Repositorio Nacional de Germoplasma del Departamento de Agricultura de EE.UU. en Miami, Florida (USDA-ARS), previamente indexados y encontrados infectados con ASBVd basados en la técnica de RT-PCR (Schnell et ál., 1997), se utilizaron como fuente de explantes nucelares para la inducción de cultivos embriogénicos.
Inducción y mantenimiento de cultivos embriogénicos en medio semisólido
Los pétalos y los sépalos se removieron y los frutos se enjuagaron con agua corriente a temperatura ambiente durante 1 hora. Después del enjuague, los frutos se desinfectaron superficialmente por 20 min en una solución al 1.25% (v/v) de hipoclorito de sodio con 5 gotas de Tween 20® y se enjuagaron tres veces con agua destilada y esterilizada. Seguidamente, los frutos se bisecaron longitudinalmente dentro de una cámara de flujo laminar y, con la ayuda de un microscopio de disección, el embrión zigótico y el tejido endospérmico de cada fruto se removieron y descartaron, mientras que el nucelo adherido a los integumentos se aisló e inoculó en medio semisólido para inducción de cultivos embriogénicos de aguacate (MIA), previamente descrito por Witjaksono y Litz (1999). El MIA contenía las sales mayores B5 (Gamborgetál., 1968) y se suplementó con las sales menores de Murashige y Skoog (MS) (1962), 0.41 |iM de picloram y (en mg/L-1) tiamina HCl (0.4), mio-inositol (100), sucrosa (30,000) y TC agar (8,000) (Carolina Biological Supply). El medio de cultivo se esterilizó y distribuyó en porciones de 10 mL en platos de Petri plásticos (60 x 15 mm). Cuatro explantes, una mitad de nucelo cada uno, se inocularon en cada plato; los platos se sellaron con Para-film® y se mantuvieron en la oscuridad a una temperatura de 25 °C durante 4 a 5 semanas. De cada material se tomó el mayor número posible de
|
||
|
|
|||
|
11
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VII No. 2 Diciembre 2005 10-18
|
|||
|
|
|||
|
estructuras florales para la obtención de explantes, lo que permitió establecer un total de 464 explantes de 'Vero Beach' SE2, 80 de 'Vero Beach' 1 y 296 de 'Yama' 381. Se registró el número de cultivos em-briogénicos inducidos y se calcularon los porcentajes de inducción.
Los cultivos embriogénicos que se desarrollaron de los nucelos inoculados en MIA, se transfirieron individualmente a medio de mantenimiento semisólido (Witjaksono y Litz, 1999) donde prolifera-ron. El medio semisólido consistió de las sales mayores y menores de MS suplementado con 0.41 |iM de picloram y (en mg/L-1) tiamina HCl (0.4), mio-ino-sitol (100), sucrosa (45,000) y TC agar (8,000). El medio se esterilizó y distribuyó similarmente a lo indicado para MIA. Los cultivos se transfirieron a medio de cultivo fresco de la misma composición cada cuatro semanas.
Mantenimiento de cultivos en medio líquido
Los cultivos en suspensión se establecieron con aproximadamente 200 mg de tejido embriogénico de dos semanas de edad que se inocularon en erlen-meyers de 125 mL que contenían 40 mL de medio líquido MS3:1N (Witjaksono y Litz, 1999). MS3:1N consistió de MS modificado con 15 mM de NH4NO3 y 30 mM de KNO3, sales menores de MS, 0.41 |iM de 2,4-D y (en mg/L-1), tiamina HCl (0.4), mio-inositol (100) y sucrosa (45,000). Los recipientes se cubrieron con papel aluminio, se sellaron con Parafilm® y se mantuvieron en semioscuridad a 25 °C en un agitador orbital a 120 rpm. Los cultivos se transfirieron cada dos semanas a medio fresco de la misma formulación.
Con el fin de caracterizar el comportamiento de los tejidos inducidos a partir de plantas infectadas con ASBVd, después de tres subcultivos en medio fresco, el crecimiento de los cultivos en suspensión se analizó y comparó con los de la línea embriogéni-ca 'Hass'11.4.3 libre de ASBVd, inducida de embriones zigóticos obtenidos de la colección de germo-plasma de la Universidad de California, Riverside, CA, y mantenida en medio líquido MS3:1N por 18 meses al momento de la evaluación. Para la evaluación, los cultivos en suspensión de dos semanas de edad se homogenizaron a través de un filtro estéril de 1.8 mm de diámetro y la fracción más pequeña se colectó. El inóculo consistente de 1 mL de tejido embriogénico se obtuvo decantando los cultivos en
|
cilindros graduados de 50 mL e inoculándolo en er-lenmeyers de 125 mL de capacidad a los cuales se les adicionó medio MS3:1N hasta alcanzar un volumen final de 40 mL. Los tejidos se cultivaron como se indicó anteriormente y el incremento de volumen se registró a los 3, 6, 9, 12, 15, 18, 21, 24, 27 y 30 días después de la inoculación. Las muestras se distribuyeron en un diseño completamente al azar con cuatro genotipos y cinco réplicas por cada genotipo, para un total de 20 unidades experimentales. Los datos obtenidos se estudiaron con análisis de va-rianza usando el modelo estadístico Yi = ¡i + i + ei; donde Y fue el volumen, ifue el cultivar y e el error experimental. Los promedios se compararon usando la prueba de rangos múltiples de Duncan con un nivel de significancia de a = 0.05.
Esterilización de medios de cultivo
El pH de todos los medios se ajustó a 5,7 - 5,8 con KOH o HCl antes de la adición del agente solidificante; posteriormente el medio se esterilizó en un autoclave a 121 °C a 1.1 kg cm-2 por 15 min y finalmente se dispensó como se indicó para cada uno de los casos.
RT-PCR
Extracción de ARN. Se extrajo ARN usando el método de Ainsworth (1995) modificado por Schnell et ál. (1997) a partirde 100 mg de tejidos, se disolvió en 20 |iL de agua tratada con DEPC (dietil pirocarba-mato) y se almacenó a -20 °C.
Síntesis de la primera cadena complementaria de ADN. La primera cadena complementaria de ADN (cADN) se sintetizó usando 10 |iL de ARN mezclado con 3 |iL de 5X buffer (Gibco BRL) (50 mM Tris HCl (pH 8,3), 75 mM KCl y 3 mM MgCl2), 0.1 M DTT y 0.5 |ig del primer (5'-AAGTCGAAACTCAGAGTCGG-3) complementario a los nucleótidos 68-87 de la región conservada de la molécula de ASBVd (Bar-Joseph et ál., 1985). La mezcla se incubó a 100 °C por 5 minutos, luego se enfrió en hielo por 2 minutos y después se colocó a temperatura ambiente por 1 hora para permitir la adhesión del primer. Posteriormente, los siguientes componentes se adicionaron: 2 ¡A. de 5X buffer, 25 mM de cada dinucleotide (ATP, CTP, GTP, TTP), 500 |iM de p-mercaptoetanol, 20 unidades de RNAsin (inhibidor de RNAsa) (Promega) y 200
|
||
|
|
|||
|
12
|
|||
|
|
|||
|
|
|||
|
EL VIROIDE DE LA MANCHA DEL SOL (ASBVd) ES PERSISTENTE EN CULTIVOS NUCELARES
|
|||
|
|
|||
|
unidades de transcriptasa reversa M-MLV (Gibco BRL). El volumen se ajustó a un total de 25 ¡A. con agua esterilizada y la mezcla se incubó por 150 minutos a 42 °C.
Amplificación de la primera cadena de ADN. La primera cadena de ADN se amplificó por PCR usando el primer hacia delante 5'-(AAGTCGAAACTCAGAGTCGG-3, marcado con 6-FAM) y el primer de reversa 5'-(GTGAGAGAAGGAGGAGT-3, marcado con HEX) (Integrated DNA Technology-IDT). La reacción de amplificación consistió en 2.5 ¡A. de la reacción cADN y 22.5 |iL de una mezcla compuesta de 2.5 |iL de 10X bufferde RCP, 25 mM de cada dinucleótido, 1.25 unidades de Amplitaq DNA polimerase (Perkin Elmer), 10 |ig de cada primer y agua esterilizada para ajustar el volumen. Las amplificaciones se efectuaron en un termociclador PTC 100 (MJ Research) con 30 ciclos a las siguientes condiciones: 30 segundos a 94 °C, 30 segundos a 55 °C y 30 segundos a 72 °C, con una extensión final de 7 minutos a 72 °C.
Electroforesis capilar. Los productos amplificados se analizaron usando electroforesis capilar en un analizador genético ABI Prism® 3100 (Applied Biosystems) usando un polímero optimizado de 4% (POP 4), bufferde electroforesis con EDTAy un marcador estándar interno de 500 bases (500 ROX). Los datos obtenidos se analizaron usando el software GeneScan (Applied Biosystems).
Clonación y secuencia de fragmentos amplificados en uno de los cultivares
Con el fin de determinar si las bandas amplificadas correspondían a moléculas de ASBVd, se tomaron al azar los fragmentos amplificados de una muestra del cultivar 'Vero Beach' 1, los cuales se clonaron usando el kit TOPO® cloning (Invitrogen), de acuerdo con las especificaciones del fabricante, en bacterias E. colicompetentes y estas se inocularon en medio LB suplementado con 100 mL-1 de ampicilina. Las colonias putativamente resistentes a ampicilina se inocularon en 100 |iL de medio de cultivo líquido S.O.C. [2% bactotriptona, 0.5% extracto de levadura, 10 mM NaCl, 2.5 mM KCl, 20 mM (MgCl2 6H2O + MgSO4 7 H2O), 20 mM sucrosa] suplementado con ampicilina y se cultivaron en un agitador orbital por 20 horas a 37 °C. Los cultivos se centrifugaron a 10,000 rpm durante 20 min, el sobrenadante se
|
descartó y las bacterias peletizadas se resuspendie-ron en 50 ¡xL de 10 mM Tris-HCL (pH 8,0); esta suspensión se utilizó como templete para la amplificación de los fragmentos clonados. Los productos insertados se amplificaron usando los pri-mers M13 (Promega). Los fragmentos amplificados se secuenciaron con la versión 3 del ciclo de terminación en un analizador genético ABI Prism 3100 usando el software de secuencia de ABI Prism. Los datos de las secuencias se alinearon manualmente utilizando el software Sequencer 4.1® (Gene Codes) y se compararon con variantes de ASBVd reportadas previamente.
RESULTADOS Y DISCUSIÓN
Inducción y mantenimiento de cultivos embriogénicos en medio semisólido
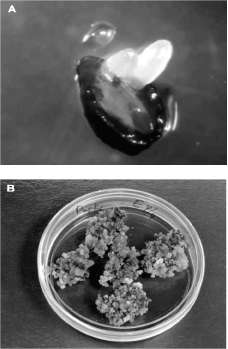
Los cultivos embriogénicos se indujeron entre la segunda y la sexta semanas después de la inoculación en MIA. Los tejidos proliferaron directamente de células presentes en los explantes sin la formación de un crecimiento intermedio de callo (figura 1A). Las frecuencias de inducción fueron de 1.5% para 'Vero Beach' SE2, 6.25% para 'Yama' 381 y 6.25% para 'Vero Beach' 1. Como resultado de los bajos índices de inducción no fue posible inferir ningún efecto estadístico. Las estructuras inducidas consistieron de proembriones, embriones somáticos en estado globular y masas proembriogénicas (MPEs) (datos no mostrados). Previamente, cultivos embriogénicos nu-celares han sido inducidos en especies de cítricos (Weathers y Calavan, 1959; Rangan et ál., 1968); Mangifera indica (Litz, 1984; Litz et ál., 1982), Carica pentagona (Vega de Rojas y Kitto, 1991) y Persea americana Mill. (Witjaksono et ál., 1999). Aunque las frecuencias de inducción observadas fueron relativamente bajas (1.5 - 6.5%) si se comparan con cultivos nucelares de cítricos (>10%) (Weathers y Calavan, 1959; Rangan et ál., 1968) o Carica pentagona (>40%) (Vega de Rojas y Kitto, 1991), los niveles de inducción obtenidos en este estudio están en correspondencia con los porcentajes de inducción previamente reportados para explantes nucelares de aguacate donde los porcentajes de inducción han estado entre 1 - 5% (Witjaksono et ál., 1999).
Una vez transferidos a medio de mantenimiento semisólido, los cultivos embriogénicos que proliferaron en MPEs mantuvieron esta característica después de varias transferencias a medio
|
||
|
|
|||
|
13
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VII No. 2 Diciembre 2005 10-18
|
|||
|
|
|||
|
fresco, mientras que los que fueron inducidos ini-cialmente como proembriones se tornaron más pequeños a medida que proliferaron y finalmente quedaron compuestos por MPEs después de varios subcultivos (figura 1B); los tejidos que iniciaron como embriones somáticos globulares se tornaron hiperhídricos y se deterioraron. A pesar que los datos de inducción y las características de los cultivos en el medio de mantenimiento no permiten realizar ninguna correlación con su origen a partir de plantas infectadas con ASBVd, al comparar los resultados obtenidos con los reportados para cultivos inducidos a partir de plantas sanas (Witjakso-noy Linz, 1999; Pliego-Alfaroy Murashige, 1988), se observa claramente que la infección de ASBVd en la planta que provee los explantes no es una limitante para la inducción y multiplicación de cultivos embriogénicos a partir de esta.
|
Mantenimiento de cultivos nucelares en medio líquido
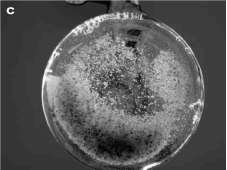
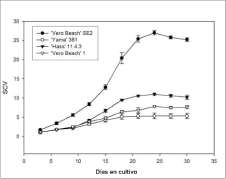
Las masas proembriogénicas establecidas en el medio de cultivo líquido mantuvieron esta característica después de varios subcultivos (figura 1C). La figura 2 muestra el comportamiento de los cultivos en medio líquido y la prueba de Duncan (tabla 1) muestra que el cultivar 'Vero Beach' SE2 obtuvo el mayor volumen de precipitado de células desde el tercer día después de la inoculación y mantuvo esta tendencia. Al final del período de cultivo, 'Vero Beach' SE2 alcanzó el mayor volumen, seguido por el cultivar "Hass" 11.4.3 y los cultivares 'Vero Beach' 1 y 'Ya-ma' 381, en su orden. Todos los cultivares alcanzaron el mayor volumen a los 24 días después de inoculados y la mayor tasa de crecimiento ocurrió entre los 15 y los 18 días para 'Vero Beach' SE2, "Hass" 11.4.3 y 'Vero Beach' 1, mientras que para 'Yama' 381 esta se dio entre los días 9 y 12 después de la inoculación.
El análisis del incremento de volumen a través del tiempo demostró que todos los cultivos presentaron un crecimiento típico de una suspensión celular observado en estudios realizados con material sano (Witjaksono y Litz, 1999), con una fase inicial de bajo crecimiento durante la primera semana, seguida por una de crecimiento más activo en la segunda semana, decreciendo un poco hacia el final de la tercera semana y terminando con una fase de cese del crecimiento al finalizar la cuarta semana de cultivo (figura 2). Este comportamiento de los tejidos indica que, independientemente de la condición patológica de la
|
||
 |
|||
 |
|||
|
|
|||
|
Figura 1. Cultivos embriogénicos de aguacate; A: inducción a partir del nucelo, B: mantenimiento en medio semisólido y C: mantenimiento en medio líquido.
|
|||
|
|
|||
|
14
|
|||
|
|
|||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
EL VIROIDE DE LA MANCHA DEL SOL (ASBVD) ES PERSISTENTE EN CULTIVOS NUCELARES
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
sido inducidos mostró que estos permanecieron infectados con ASBVd.
Electroforesis capilar
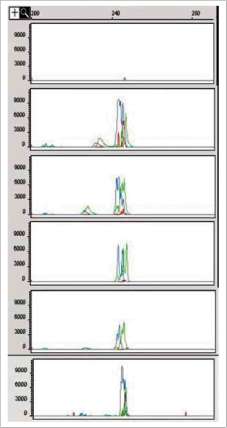
El electroferograma (figura 3) muestra que los tejidos embriogénicos consistentes de MPEs mantenidos un año después de inducidas de los cultivares 'Vero Beach' SE2, 'Vero Beach' 1 y 'Yama' 381 amplificaron fragmentos similares en tamaño (aprox. 250 bp) a los del control positivo (tejido floral de 'Vero Beach' SE2), mientras que ningún tipo de amplificación se observó en las MPEs de la línea em-briogénica 'Hass' 11.4.3 utilizada como control negativo libre de ASBVd. Resultados similares se observaron a los 6 y 9 meses después de la inducción (datos no mostrados). Bitters et ál. (1972), con base en pruebas realizadas con plantas indicadoras, informaron la recuperación de plantas cítricas sanas regeneradas a partir de tejido nucelar de plantas infectadas con el viroide de exocortis de los cítricos (CEVd). Recientemente, Postman y Hadidi (1995), aplicando termoterapia y cultivo de meristemos, y aplicando RT-PCR, obtuvieron un 86% de plantas de pera sanas a partir de tejidos vegetales aislados de árboles infectados con el viroide de la mancha de la piel del peral (ASSVd); igualmente, Howell et ál. (1998) reportaron la eliminación de ASSVd de
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Figura 2. Incremento de volumen de cultivos embriogénicos en medio líquido.
planta de origen, los tejidos pueden proliferar en medio de cultivo líquido en un comportamiento similar al de un cultivo sano, demostrando que cultivos embriogénicos de plantas infectadas pueden desarrollarse en medio líquido para efectos de estudio de ASBVd en condiciones in vitro.
RT-PCR
La indexación con RT-PCR efectuada en los cultivos nucelares a los 6, 9 y 12 meses después de haber
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Tabla 1. Volumen de precipitado de células en medio de cultivo líquido y significancia con base en la prueba de Duncan (a = 0.05)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Valores con diferentes letras son significativamente diferentes al 0.05%.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
15
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VII No. 2 Diciembre 2005 10-18
|
|||
|
|
|||
 |
tejidos nucelares, se cree que esto es resultado de la ausencia de vasos vasculares en este tejido (Ran-gaswami, 1981), lo cual es soportado por el hecho de que CEVd y otros miembros de la familia Pospivi-roidae se localizan y replican en el floema, conducto que igualmente utilizan para desplazarse dentro de la planta (Momma y Takahashi, 1983; Bonfiglioli et ál., 1996). ASBVd y los otros miembros de la familia Avsunviroidae tienen marcadas diferencias estructurales y moleculares con respecto a los viroides de la familia Pospiviroidae. Los primeros no tienen una región central conservada de nucleótidos (RCC), se localizan y replican en los cloroplastos y tienen la capacidad de autofragmentarse durante el proceso de replicación usando solamente riboenzimas presentes en su estructura. Contrariamente, los segundos se caracterizan por tener una RCC, replicarse en el núcleo de la célula y utilizar enzimas del hospedero para separarse en moléculas singulares durante la replicación (Flores et ál., 1998). Aunque el mecanismo que los viroides de la familia Avsunviroidae utilizan para desplazarse dentro de la planta no se ha descubierto, los resultados de la presente investigación indican que este seguramente es diferente al movimiento vascular a través del floema, utilizado por los viroides de la familia Pospiviroidae, ya que solo de esta manera se explicaría el hecho de que ASBVd puede infectar un tejido sin conexiones vasculares como es el nucelo. Recientemente, indicadores como la conformación de la estructura secundaria y la secuencia de nucleótidos de la molécula (Fadda et ál., 2003a, b) se consideran como base para diferencias taxonómicas.
Clonación, secuencia y análisis de variantes
Los fragmentos amplificados de la muestra del cultivar 'Vero Beach' 1 fueron seleccionados, clonados y secuenciados. Cuando las secuencias obtenidas se compararon en la base de datos del banco de genes del National Center for Biotechnology Information (NCBI), la búsqueda reveló que cinco clones contenían secuencias con más de un 97% de similari-dad con variantes de ASBVd previamente reportadas. La alineación de las secuencias de los clones seleccionados permitió establecer que la población clonada estaba compuesta por tres variantes de ASBVd, como se detalla a continuación: un clon contenía la variante CF3, uno contenía la variante CF8, dos clones contenían la variante CF13 y uno más tenía insertada la variante CF24, todas previamente clonadas de plantas establecidas en el National
|
||
|
Figura 3. Electroferogramas de (en orden de arriba abajo): MPEs de 'Hass' 11.4.3. (control negativo), 'Vero Beach' SE2 (12 meses después de inducidas - DDI), 'Vero Beach' 1 (12 meses DDI), 'Yama' 381 (12 meses DDI) y tejido floral de 'Vero Beach' SE2 (control positivo).
|
|||
|
plantas de manzana después de usar termoterapia y cultivo de meristemos y realizar indexación con RT-PCR. Los resultados obtenidos en el presente estudio demuestran que ASBVd no se elimina mediante el cultivo de tejidos embriogénicos inducidos a partir de células nucelares y, más importante aún, que permanece por tiempo indefinido replicándose en la medida en que los cultivos embriogénicos proli-feran. Aunque no se conoce el mecanismo exacto por el cual CEVd es eliminado mediante el cultivo de
|
|||
|
|
|||
|
16
|
|||
|
|
|||
|
|
|||
|
EL VIROIDE DE LA MANCHA DEL SOL (ASBVD) ES PERSISTENTE EN CULTIVOS NUCELARES
|
|||
|
|
|||
|
Germoplasm Repository (NGR, Miami, FL). Estos resultados demuestran que los fragmentos amplificados en el proceso de RT-PCR fueron en efecto moléculas de ASBVd. La presencia de variabilidad en la secuencia de las moléculas de ASBVd afectando a un solo individuo ha sido previamente reportada (Semancik y Szychowsky, 1994; Schnell et ál., 20001a, b), fenómeno que ha sido relacionado con una variación del concepto de cuasiespecies (Duar-te et ál., 1994; Ambros et ál., 1999; Schnell et ál., 2001a). Los resultados de las secuencias obtenidas demuestran la persistencia de ASBVd en plantas recuperadas a partir de tejidos similares a los aquí inducidos, lo cual indica que el cultivo nucelar no es un mecanismo efectivo para obtener plantas de aguacate sanas a partir del cultivo de nucelos obtenidos de cultivares infectados con ASBVd.
CONCLUSIONES
En el presente estudio, a pesar de no lograr la eliminación de ASBVd mediante el procedimiento utilizado, el uso de la embriogénesis somática ha permitido la inducción, multiplicación y mantenimiento de material de aguacate infectado con ASBVd en condiciones in vitro, y aunque se ha demostrado que ASBVd persiste en los cultivos inducidos a partir de plantas infectadas, los resultados indican que existe la posibilidad de realizar estudios de análisis de variabilidad genética en tejidos embriogénicos de aguacate para profundizar en el conocimiento de este particular patógeno.
AGRADECIMIENTOS
Los autores desean agradecer el apoyo de la Universidad de Córdoba, Colciencias, la Comisión Ful-bright, University of Florida - Tropical Research and Education Center, California Avocado Commission, Pamela Moon y Cecile T. Olano.
BIBLIOGRAFÍA
Ainsworth, C. 1995. Isolation of RNA from floral tissue of Ru-mexacetosa (Sorrel). PlantMol. Biol. Rep. 12:198-203.
Ambros, S.; Hernández, C.; Flores, R. 1999. Rapid generation of genetic heterogeneity in progenies from individual cDNA clones of peach latent mosaic viroid in its natural host. Gen. Virol. 80: 2239-2252.
Bar-Joseph, M.; Segev, D.; Twizer, S.; Rosner, A. 1985. De-tection of avocado sunblotch viroid by hybridization with synthetic oligonucleotide probes. J. Virol. Meth. 10: 69-73.
|
Bitters, W. P.; Murashige, T.; Rangan, T. S.; Nauer, E. 1972. Investigation on establishing virus free citrus plants through citrus culture. In: Proceedings of International Organization of Citrus Virologists. Gainesville: University of Florida Press, pp. 267-271.
Bonfiglioli, R. G.; Webb, D. R.; Symons, R. H. 1996. Tissue and intra-cellular distribution of coconut cadang cadang viroid and citrus exocortis viroid determined by in situ hybridization and confocal laser scanning and transmis-sion electron microscopy. Plant J. 9: 457-465.
Bonfiglioli, R. G.; McFadden, G. I.; Symons, R. H. 1994.In situ hybridization localizes avocado sunblotch viroid on chloroplast thylakoid membranes and coconut cadang cadang viroid in the nucleus. Plant J. 6: 99-103.
Coit, E. 1928. Sunblotch of the avocado. A serious physiologi-cal disease. Cal. Avo. Soc. Yrb. 14: 27-32.
Da Graca, J. V.; Mason, T. E.; Antel, H. J. 1983. Effectof avocado sunblotch disease on fruit yield. S. Afr. Avo. Grow. Assoc. Yrb. 6: 86-87.
Desjardins, P. R. 1958. Callus tissue growth on avocado stem segments cultured on artificial media. Cal. Avo. Soc. Yrb. 42:99-101.
Desjardins, P. R.; Drake, R. J.; Atkins, E. L.; Bergh, B. O.1979. Pollen transmission of avocado sunblotch virus experi-mentally demonstrated. Cal. Agric. 33: 14-15.
Desjardins, P. R.; Drake, R. J.; Swiecki, S. A. 1980. Infectivity studies of avocado sunblotch disease causal agent, possibly a viroid rather than a virus. Plant. Dis. 64: 313-315.
Duarte, E. A.; Novella, I. S.; Weaver, S. C.; Domingo, E.; Wain-Hobson, S; Clark, D. K.; Moya, A.; Elena, S. F.; de la Torre, J. C.; Holland, J. J. 1994. RNA virus quasispe-cies: significance for viral disease and epidemiology. Infec. Agents Dis. 3: 201-214.
Fadda, Z.; Daros, J. A.; Flores, R.; Duran-Vila, N. 2003a. Eggplant latent viroid, the candidate type species for a new genus within the Avsunviroidae family (hammmer-head viroids). J. Virol. 67: 6528-6532.
Fadda, Z.; Daros, J. A.; Flores, R.; Duran-Vila, N. 2003b. Iden-tification in eggplant of a variantof citrus exocortis viroid (CEVd) with a 95 nucleotide duplication on the right terminal region of the rod-like secondary structure. Virus Res. 97: 145-149.
Flores, R.; Randles, J. W.; Bar-Josep, M.; Diener, T. O. 1998. A proposed scheme for viroid classification and nomen-clature. Arch. Virol. 143: 623-639.
Gamborg, O. L.; Miller, R. A.; Ojima, K. 1968. Plant cell cultures. I. Nutrient requirements of suspension cultures of soybean rootcells. Exp. Cell Res. 50: 151-158.
Horne, W. T.; Parker, E. R. 1931. The avocado disease called sunblotch. Phytopathology. 21: 235-238.
Howell, W. E.; Burgess, J.; Mink, G. I.; Skrzcczkowsky, L. J.; Zhang, Y. P. 1998. Elimination of apple fruit and bark deforming agents by heat therapy. Acta Hortic. 472: 641-646.
|
||
|
|
|||
|
17
|
|||
|
|
|||
|
|
|||
|
Rev. Colomb. Biotecnol. Vol. VII No. 2 Diciembre 2005 10-18
|
|||
|
|
|||
|
Litz, R. E. 1984. In vitro somatic embryogenesis from nucellar callus of monoembryonic Mangifera indica L. HortScien-ce. 19:715-717.
Litz, R. E.; Knight, R. J. Jr.; Gazit, S. 1982. Somatic embryos from cultured ovules of Mangifera indica L. Plant Cell Rep. 1:264-266.
Momma, T.; Takahashi, T. 1983. Cytopathology of shoot apical meristem of hop plants infected with hop stuntviroid. Phytopathology. 106: 272-289.
Murashige, T.; Skoog, F. 1962. A revised medium for rapid growth of and bioassays with tobacco tissue cultures. Physiol. Plant. 15: 473-497.
Navarro, J. A.; Daros, J. A.; Flores, R. 1999. Complexes con-taining both strands of avocado sunblotch: identification in chloroplasts and characterization. Virology. 253: 77-85.
Palukaitis, P.; Hatta, T.; Alexander, D.; Symons, R. 1971. Characterization of a viroid associated with avocado sunblotch disease. Virology. 99: 145-151.
Pliego-Alfaro, F.; Murashige, T. 1988. Somatic embryogenesis in avocado (Persea americana Mill.). Plant Cell Tiss. Org. Cult. 12:61-66.
Postman, J. D.; Hadidi, A. 1995. Elimination of apple scar skin viroid from pears by in vitro termo-therapy and apical meristem culture. Acta Hortic. 386: 536-543.
Rangan, T. S.; Murashige, T.; Bitters, W. P. 1968. In vitro initia-tion of nucellarembryos in monoembryonic citrus. Hort-Science. 3: 226-227.
Rangaswami, N. S. 1981. Nucellus as an experimental system in basic and applied tissue culture research. In: Rao, A. N. (ed.). Tissue Culture of Economically Important Plants. Singapore: COSTED and ANBS, pp. 269-286.
Schnell, R. J.; Kuhn, D. N.; Ronning, C. M.; Harkin, D. 1997. Application of RT-PCR for indexing avocado sunblotch viroid. Plant Dis. 81: 1023-1026.
|
Schnell, R. J.; Kuhn, D. N.; Olano, C. T.; Quintanilla, W. E. 2001a. Sequence diversity among avocado sunblotch viroids isolated from single avocado trees. Phytoparasi-tica. 29:451-460.
Schnell, R. J.; Olano, C. T.; Kuhn, D. N. 2001b. Detection of avocado sunblotch viroid variants using fluorescent sin-gle-strand conformation polymorphism analysis. Elec-trophoresis. 22: 427-432.
Semancik, J. S.; Szychowsky, J. A. 1994. Avocado sunblotch: a persistent viroid infection in which variants are associated with differential symptoms. J. Gen. Virol. 75: 1543-1549.
Stevens, H. E.; Piper, R. B. 1941. Sunblotch in avocado disea-ses in Florida. USDA. Washington DC, pp. 40-46.
Symons, R. H. 1981. Avocado sunblotch viroid: primary sequence and proposed secondary structure. Nuc. Acids Res. 9: 6527-6537.
Vega de Rojas, R.; Kitto, S. L. 1991. Regeneration of babaco (Carica pentagona) from ovular callus. J. Amer. Soc. Hort.Sci. 116:747-752.
Wallace, J. M.; Drake, R. J. 1962. Ahigh rateofseed transmis-sion of avocado sunblotch from symptomless trees and the origin of such trees. Phytopathology. 52: 237-241.
Weathers, L. G.; Calavan, E. C. 1959. Nucellar embryony as a means of freeing citrus clones of viruses In: Wallace, J. M (ed.). Citrus Virus Diseases, Berkeley: University of California Division of Agricultural Sciences, pp. 197-200.
Witjaksono; Litz, R. E. 1999. Induction and growth characteris-tics of embryogenic avocado cultures. Plant. Cell Tiss. Org. Cult. 59: 19-29.
Witjaksono; Litz, R. E.; Pliego-Alfaro, F. 1999. Somatic embryogenesis of avocado. In: Jain, S. M.; Gupta P. K.; Newton, R. J. (eds.). Somatic Embryogenesis of Woody Plants, vol. 5. Dordrecht: Kluwer, pp. 197-214.
|
||
|
|
|||
|
18
|
|||
|
|
|||
Cómo citar
APA
Suárez, I. E. (2005). El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.). Revista Colombiana de Biotecnología, 7(2), 10–18. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490
ACM
[1]
Suárez, I.E. 2005. El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.). Revista Colombiana de Biotecnología. 7, 2 (jul. 2005), 10–18.
ACS
(1)
Suárez, I. E. El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.). Rev. colomb. biotecnol. 2005, 7, 10-18.
ABNT
SUÁREZ, I. E. El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.). Revista Colombiana de Biotecnología, [S. l.], v. 7, n. 2, p. 10–18, 2005. Disponível em: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490. Acesso em: 20 mar. 2026.
Chicago
Suárez, Isidro E. 2005. «El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)». Revista Colombiana De Biotecnología 7 (2):10-18. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490.
Harvard
Suárez, I. E. (2005) «El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)», Revista Colombiana de Biotecnología, 7(2), pp. 10–18. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490 (Accedido: 20 marzo 2026).
IEEE
[1]
I. E. Suárez, «El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)», Rev. colomb. biotecnol., vol. 7, n.º 2, pp. 10–18, jul. 2005.
MLA
Suárez, I. E. «El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)». Revista Colombiana de Biotecnología, vol. 7, n.º 2, julio de 2005, pp. 10-18, https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490.
Turabian
Suárez, Isidro E. «El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.)». Revista Colombiana de Biotecnología 7, no. 2 (julio 1, 2005): 10–18. Accedido marzo 20, 2026. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490.
Vancouver
1.
Suárez IE. El viroide de la mancha del sol (ASBVd) es persistente en cultivos nucelares de aguacate (Persea americana Mill.). Rev. colomb. biotecnol. [Internet]. 1 de julio de 2005 [citado 20 de marzo de 2026];7(2):10-8. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/490
Descargar cita
Visitas a la página del resumen del artículo
858
Descargas
Los datos de descargas todavía no están disponibles.
Licencia
Derechos de autor 2005 Revista Colombiana de Biotecnología

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Esta es una revista de acceso abierto distribuida bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (CC BY). Se permite el uso, distribución o reproducción en otros medios, siempre que se citen el autor(es) original y la revista, de conformidad con la práctica académica aceptada. El uso, distribución o reproducción está permitido desde que cumpla con estos términos.
Todo artículo sometido a la Revista debe estar acompañado de la carta de originalidad. DESCARGAR AQUI (español) (inglés).















