



Publicado
2004-07-01
Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis
Transcriptional analysis of genetic region RvD1 of Mycobacterium bovis
Palabras clave:
Mycobacterium bovis, BCG, RNA, Real Time, RT-PCR, RvD1, RvD1. (es)
Mycobacterium bovis comparte una identidad del 99,9% con los genomas de M. tuberculosis, M. africanum y M. microti. Dentro del 0,1% de esta diferencia se encuentran dos regiones genéticas propias de M. bovis: RvD1 y RvD2, las cuales se encuentran delecionadas del genoma de M. tuberculosis H37Rv y, según el análisis bioin-formático, contienen probables marcos abiertos de lectura (Open Reading Frames: ORF). Con el fin de determinar si la región RvD1, transcribe los ORF predichos por bioinformática: ORF1, ORF2 y Rv2024, se extrajeron muestras de ARN total de M. bovis BCG Pasteur, en diferentes puntos de una curva de crecimiento micobacteriano, las cuales fueron analizadas mediante la técnica de Transcripción Reversa y Reacción en Cadena de la Polimerasa (RTq-PCR) en tiempo real. Los hallazgos obtenidos en esta cinética de transcripción por RTq-PCR en tiempo real demostraron que los probables marcos de lectura abiertos ORF1, ORF2 y Rv2024 de la región RvD1 de M. bovis, sí se transcriben y lo hacen de manera constitutiva, hecho que no había sido reportado. Los resultados de esta investigación sirven como un primer paso para determinar la función que desempeña la región RvD1 de M. bovis, y su posible papel en la patogénesis y en la interacción huésped-patógeno de la tuberculosis bovina y humana.
Palabras clave: Mycobacterium bovis, BCG, RNA, RT-PCR, RvD1.
Mycobacterium bovis, shares 99.9% of genomic identity with M. tuberculosis, M. africanum and M. microti. Within this 0.1 % of difference, there are two genetic regions characteristics of M. bovis that are deleted in M. tuberculosis H37Rv: RvD1 and RvD2. According to bioinformatic analysis, these regions contain Open Reading Frames (ORFs). With the purpose of determining if the RvD1 region transcribes the ORFs predicted by bioinformatics (ORF1, ORF2 and Rv2024); total RNA was extracted from a culture of M. bovis BCG Pasteur, at different time points along the growth curve. The RNA samples were analyzed by Real Time Reverse Transcription - Poly-merase Chain Reaction (RTq-PCR). The findings show that ORF1, ORF2 and Rv2024, were transcribed consti-tutively, something that has not been reported previously. These results are a first step in order to determine the function of M. bovis RvD1 region, its possible role in pathogenesis and its interaction with both cattle and humans.
Key words: Mycobacterium bovis, BCG, RNA, Real Time, RT-PCR, RvD1
|
|
||
|
REVISTA COLOMBIANA DE BIOTECNOLOGÍA VOL.VI No. 2 Diciembre 2004 62-66___________________
Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis
Transcriptional analysis of genetic region RvD1 of Mycobacterium bovis
Víctor Manuel Tibatá R.*, Clara Eugenia González**, Juan Germán Rodríguez***, Patricia del Portillo***
RESUMEN
Mycobacterium bovis comparte una identidad del 99,9% con los genomas de M. tuberculosis, M. africanum y M. microti. Dentro del 0,1% de esta diferencia se encuentran dos regiones genéticas propias de M. bovis: RvD1 y RvD2, las cuales se encuentran delecionadas del genoma de M. tuberculosis H37Rv y, según el análisis bioin-formático, contienen probables marcos abiertos de lectura (Open Reading Frames: ORF). Con el fin de determinar si la región RvD1, transcribe los ORF predichos por bioinformática: ORF1, ORF2 y Rv2024, se extrajeron muestras de ARN total de M. bovis BCG Pasteur, en diferentes puntos de una curva de crecimiento micobacteriano, las cuales fueron analizadas mediante la técnica de Transcripción Reversa y Reacción en Cadena de la Polimerasa (RTq-PCR) en tiempo real. Los hallazgos obtenidos en esta cinética de transcripción por RTq-PCR en tiempo real demostraron que los probables marcos de lectura abiertos ORF1, ORF2 y Rv2024 de la región RvD1 de M. bovis, sí se transcriben y lo hacen de manera constitutiva, hecho que no había sido reportado. Los resultados de esta investigación sirven como un primer paso para determinar la función que desempeña la región RvD1 de M. bovis, y su posible papel en la patogénesis y en la interacción huésped-patógeno de la tuberculosis bovina y humana.
Palabras clave: Mycobacterium bovis, BCG, RNA, RT-PCR, RvD1.
ABSTRACT
Mycobacterium bovis, shares 99.9% of genomic identity with M. tuberculosis, M. africanum and M. microti. Within this 0.1 % of difference, there are two genetic regions characteristics of M. bovis that are deleted in M. tuberculosis H37Rv: RvD1 and RvD2. According to bioinformatic analysis, these regions contain Open Reading Frames (ORFs). With the purpose of determining if the RvD1 region transcribes the ORFs predicted by bioinformatics (ORF1, ORF2 and Rv2024); total RNA was extracted from a culture of M. bovis BCG Pasteur, at different time points along the growth curve. The RNA samples were analyzed by Real Time Reverse Transcription - Poly-merase Chain Reaction (RTq-PCR). The findings show that ORF1, ORF2 and Rv2024, were transcribed consti-tutively, something that has not been reported previously. These results are a first step in order to determine the function of M. bovis RvD1 region, its possible role in pathogenesis and its interaction with both cattle and humans.
Key words: Mycobacterium bovis, BCG, RNA, Real Time, RT-PCR, RvD1
|
||
|
|
||
|
MV. M. Se, Investigador Corporación Corpogen. ** Bacterióloga, investigadora Corporación Corpogen. *** Biólogo, investigador Corporación Corpogen. **** Directora ejecutiva, Corporación Corpogen, Cra. 5 No. 66-88, Bogotá. Colombia. Correo electrónico:
corpogen@etb.net.co
Recibido: septiembre 27 de 2004. Aceptado: octubre 29 de 2004.
|
||
|
|
||
|
62
|
||
|
|
||
|
|
|||
|
ANÁLISIS TRANSCRIPCIONAL DE LA REGIÓN RvD1 DE M. bovis
|
|||
|
|
|||
|
INTRODUCCIÓN
El complejo M. tuberculosis además de M. bovis, que tiene la capacidad de infectar al bovino, al hombre y a gran variedad de huéspedes mamíferos de importancia económica y silvestre, también agrupa a las especies M. microti patógeno del ratón yaM tuberculosis y M. africanum patógenos para el humano (O'Reilly, 1995). Estos cuatro microorganismos comparten una identidad del 99,9% entre sus genomas, por tanto, el estudio de la diferencia genética del 0,1 % se constituye en una clave fundamental para entender los diferentes mecanismos de adaptación a sus huéspedes, la interacción hospedero-patógeno, la virulencia para el hombre y las características fisiológicas de estas bacterias (Gordon et al., 1999).
Con el fin de encontrar una región específica de M. bovis, dentro del 0,1% de diferencia genética, que distinguiese esta especie de las demás micobacte-rias, en particular de M. tuberculosis, en 1995 el grupo de investigación, mediante la técnica de Amplificación Arbitraria de Fragmentos Polimórficos de DNA (RAPD), identificó un fragmento polimórfico de 2,9 kb, presente en el genoma de Mycobacterium bovis y ausente en 29 especies diferentes de micobacterias incluyendo M. tuberculosis H37Rv (Rodríguez et al., 1995). Este fragmento de 2,9 kb fue posteriormente secuenciado por el grupo de S. T. Cole, quien demostró que hace parte de la región que ellos denominaron RvD1 de M. bovis y que compromete 6000 pb que se encuentran delecionadas en el genoma de M. tuberculosis H37Rv (Gordon et al., 1999).
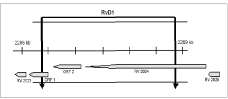
El análisis bioinformático de esta región permitió inferir la presencia de tres probables marcos abiertos de lectura (Open Reading Frames, ORF): ORF1, ORF2 y Rv2024, que in vivo podrían codificar una o más proteínas (figura 1). Las proteínas hipotéticas que codificarían dos de estos marcos de lectura
|
no presentan homología significativa con otras proteínas reportadas, mientras que la secuencia traducida del marco de lectura Rv2024 presenta similitud con proteínas hipotéticas de Helicobacterpylor, y con helicasas y metiltransferasas de diferentes microorganismos. El presente trabajo tuvo como objetivo principal demostrar la presencia de los transcritos de la región RvD1 de M. bovis.
|
||
|
MATERIALES Y MÉTODOS
Cepa de Mycobacterium bovis. La cepa que se empleó fue la BCG Pasteur 1173 P-2, la cual fue utilizada hasta hace algunos años por el Instituto Nacional de Salud (INS) para la preparación de la vacuna contra la tuberculosis de uso humano que se aplicaba en Colombia. Esta cepa de M. bovis fue cultivada a 37 °C con agitación a 90 rpm en un agitador orbital, en 100 mL de medio líquido Middlebrook 7H9 (pH 6,8) suplementado con glicerol al 0,2% (v/v), Tween 80 al 0,05% y 10% de complejo albúmina dextrosa (ADC). Se tomaron alícuotas de este cultivo en la mitad de la fase logarítmica de crecimiento y con ellas se realizó un subcultivo en un volumen de 400 mL. A los 4,10,18,22,25,29, 35,42 días se midió la densidad óptica a 600 nm; en un espectrofotómetro Beck-man DU 530, se realizaron recuentos de unidades formadoras de placa (UFC) y se tomó un volumen de 50 mL para llevara cabo la extracción del RNAtotal.
Extracción de RNA. La extracción de RNA de los cultivos fue llevada a cabo con el kit RNA Total de Corpogen. El protocolo recomendado por el fabricante fue modificado ligeramente con el objeto de poder romper con facilidad la pared de las micobacterias, sin que el RNA se degradara. Esencialmente, 50 mL de cultivo fueron sometidos a centrifugación y el precipitado bacteriano fue lavado con el tampón de lavado del kit. Se tomaron entre 50 y 200 mg de peso húmedo de bacterias, los cuales fueron resus-pendidos en 200 |jL de agua tratada con DEPC. Se agregaron 500 |jL de una solución detergente, 500 |LiL de fenol ácido, 100 |jL de cloroformo y 100 mg perlas de circonio sílica de 0,1 mm. Esta mezcla fue sometida a rompimiento mecánico mediante la agitación por un minuto en un mini Bead-Beater (Bios-pec products), posteriormente fueron mantenidas en hielo por 10 minutos y centrifugadas a 12000 g por 10 minutos a 4 °C. Se recuperó la fase acuosa, y ésta fue extraída nuevamente con un volumen de
|
|||
 |
|||
|
Figura 1. Región RvD1 de Mycobacterium bovis con los tres marcos de lectura: ORF1, ORF2 y Rv2024.
|
|||
|
|
|||
|
63
|
|||
|
|
|||
|
|
||||
|
REVISTA COLOMBIANA DE BIOTECNOLOGÍA VOL.VI No. 2 Diciembre 2004 62-66
|
||||
|
|
||||
|
mezcla cloroformo-alcohol isoamílico (24:1) y posteriormente fue sometida a precipitación con un volumen de isopropanol. Por último, el RNA precipitado fue lavado con etanol al 70% (v/v) y reconstituido en un volumen apropiado de tampón de reconstitución.
Cada una de las muestras de RNA total fue analizada por electroforesis en gel de agarosa al 1% (p/v) en TBE 0,5 X (Tris Borato 0,045 mM EDTA 0,001 mM, pH 8,3), teñidos con bromuro de etidio. El RNA extraído fue cuantificado a 260 nm en un es-pectrofotómetro Beckman DU 530 y tratado con DNAsa RQ1 libre de RNAsas (Promega), con el fin de asegurar la ausencia de contaminación con DNA en la preparación.
Iniciadores. Mediante el análisis bioinformático, y con la ayuda del programa OLIGO V, fueron diseñados juegos de iniciadores específicos para el ORF1, ORF2, ORF Rv2024 y para el transcrito del factor Sig-ma alternativo sigA, el cual se incluyó como gen nor-malizador para la curva de crecimiento (Hu y Coates, 1999). Todos los iniciadores fueron diseñados para que anillaran a la misma temperatura, de tal forma que todas las amplificaciones se pudieran realizar y analizar simultáneamente con cada muestra.
Las secuencias de estos iniciadores son las siguientes:
|
Como control de falsos positivos debido a la posible amplificación de DNA contaminante se incluyó una reacción que no contenía la enzima transcriptasa reversa. Este control no debía producir amplificación.
El sistema empleado para esta técnica fue el QuantiTect SYBR Green RT-PCR de Qiagen. Cada reacción se llevó a cabo en un volumen de 50 |jJ de mezcla, compuesta de: QuantiTec SYBR Green RT PCR Master Mix (1X final), iniciadores: 20 nMy 100 |ug de RNA tratado con DNAsa.
Las amplificaciones fueron realizadas en un termociclador DNA Engine Opticon de MJ Research, con los siguientes ciclos de temperatura: 50° por 30 minutos para la transcripción reversa, activación inicial: 94 °C por 15 minutos; 39 ciclos de Denatura-ción: 96 °C por 15 seg, anillaje 60 °C por 30 seg, extensión 72 °C por 30 seg; curvas de disociación: lecturas a 0,1 seg; entre 70 y 90 °C cada 0,2 °C. Con el fin de realizar la cuantificación de cada ORF, se comprobó la especificidad y la eficiencia de la amplificación de cada par de iniciadores, mediante una PCR en tiempo real con DNA genómico de Mycobacterium bovis BCG Pasteur.
Cuantificación. Para estimar el nivel de transcripción de cada uno de los marcos de lectura abiertos de la región RvD1, ORF1, ORF2 y Rv2024, durante las diferentes condiciones de crecimiento, se empleó la técnica de cuantificación comparativa, analizando el ACT generado por la diferencia entre el ciclo umbral (threshold cycle) CT del transcrito de interés y el CT del transcrito normalizador (factor sigma alternativo sigA).
El análisis de los datos de la curva de crecimiento se llevó a cabo mediante análisis de varianza a una vía de los CT para cada ORF y para sigA. Para comparar si existían diferencias de transcripción entre los tres ORF y sigA se empleó la prueba de comparación múltiple de Tukey-Kramer del programa GraphPad InStat. (www.graphpad.com).
RESULTADOS Y DISCUSIÓN
Extracción de RNA total a partir de la curva de crecimiento de M. Bovis. La medición de la densidad óptica y de los recuentos de UFC de la curva de crecimiento normal permitió observar que la fase logarítmica se inició aproximadamente desde el día 5
|
|||
|
SigA1
SigA2
ORF1-1
ORF1-2
ORF2-2
ORF2-2
Rv2024-1
Rv2024-2
|
5' AAG CTG GGC CGC ATTT CAA 3 5' TTC TCC GGG GTG ATG TCC AT 3' 5' CGC GCA ACA TTG TAT GAA TC 3' 5' GCG TGA AGA ATT CAA ACA 3' 5' TGC GGT CCT GTC GAA CCT3' 5' GCG GTC CAA GCT GCT TGT 3' 5' CCA CGC CAA GGA TTT TCA 3' 5' CCC GAA CGT CAG CTC GTC 3'
|
|||
|
RT PCR cuantitativa (RTq-PCR). Con el fin de determinar si estaban presentes los diferentes transcritos (mRNA) de cada uno de los marcos abiertos de lectura (ORF), se realizó la RT-PCR en tiempo real, a partir de cada una de las muestras de RNA total extraídas en cada punto de la curva de crecimiento normal. Como control de la eficiencia de los iniciadores en la amplificación del cDNAse incluyó una muestra de DNA de Mycobacterium bovis, y para evaluar su especificidad se incluyó una muestra de RNA de Mycobacterium smegmatis.
|
||||
|
|
||||
|
64
|
||||
|
|
||||
|
|
|||
|
ANÁLISIS TRANSCRIPCIONAL DE LA REGIÓN RvD1 DE M. bovis
|
|||
|
|
|||
|
y se prolongó hasta el día 22, a partir del cual el cultivo entró en fase estacionaria (figura 2). En cada uno de los puntos medidos se tomaron 50 mL de cultivo de cada una de las dos réplicas, y a partir de ellos se obtuvo un RNA total de buena calidad, el cual fue analizado cada vez por medio de electrofo-resis en gel de agarosa y por lectura espectrofotó-metrica a 260 nm.
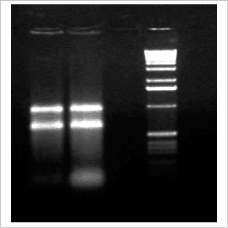
Con el kit RNA total de CorpoGen se logró extraer RNA total de muy buena calidad, tal como se puede ver en la figura 3, en donde se aprecian en forma clara las bandas del RNA ribosomal 23S y 16S, indicando la integridad y alta concentración del ácido nucleico extraído y con mínima contaminación con DNA.
Cinética de transcripción de ORF1, 2, Rv2024 y sigA a lo largo de la curva de crecimiento. La PCR
en tiempo real con DNA genómico de M. bovis, permitió demostrar que todos los iniciadores fueron igualmente eficientes en la amplificación. Las curvas de disociación de los amplificados presentaron las temperaturas de disociación (Tm) esperadas para cada ORF, reproducibles con cada dilución de DNA. De esta manera el ORF2 presentó un Tm de 81,6 °C, el ORF1 de 81,8 °C, el Rv2024 de 84,8 °C y el sigA de 87 °C.
La RT PCR en tiempo real permitió determinar que la transcripción del control sigA fue constitutiva a lo largo de la curva, de esta forma este transcrito pudo ser empleado como normalizador para la cuantifica-ción de ORF1, ORF2 y Rv2024. En cuanto a la cinética de transcripción de ORF1, ORF2 y Rv2024, analizada por RT-PCR en tiempo real, se estableció que al igual que el control de transcripción sigA, estos ORF se transcribieron en forma constitutiva en todos los ocho puntos de la curva de crecimiento normal. La
|
especificidad de las amplificaciones se comprobó con la observación de las curvas de las temperaturas de disociación de cada uno de los diferentes ORF, las cuales fueron idénticas a las generadas por la amplificación de DNA genómico en la PCR en tiempo real. Estos hallazgos indican que ORF1, ORF2 y Rv2024 se transcriben en forma constitutiva a lo largo de todas las etapas de la curva de crecimiento (figura 4).
Cuando se analizaron los datos de la cuantifica-ción con el método del ACt, se observó que los valores más bajos correspondían al ORF1; es decir, se transcribió en cantidad similar al normalizador, y de esta forma generó diferencias pequeñas entre los Cts del normalizador y de este gen; mientras que los ACt del ORF2 y de Rv2024 fueron más grandes, por ende su nivel de transcripción fue menor en relación con sigA (figura 5).
Los resultados de la cuantificación por el método del ACt, se corroboraron mediante el análisis de va-rianza a una vía, el cual determinó que algún ORF presentó transcripción diferencial a lo largo de la curva de crecimiento. Mediante la prueba de comparación múltiple de Tukey-Kramer se estableció que el gen ORF 1 se transcribió en mayor cantidad que ORF2 y Rv2024
|
||
 |
|||
 |
|||
|
Figura 3. RNA extraído con el kit RNA Total 2002 de CorpoGen.
Carril 1: extracción del día 10 de crecimiento.
Carril 2: extracción del día 15.
Carril 4: marcadores de peso molecular de 100 pb.
|
|||
|
Figura 2. Curva de crecimiento de M. bovis bajo condiciones normales (densidad óptica y UFC).
|
|||
|
|
|||
|
65
|
|||
|
|
|||
|
|
|||
|
REVISTA COLOMBIANA DE BIOTECNOLOGÍA VOL.VI No. 2 Diciembre 2004 62-66
|
|||
|
|
|||
 |
 |
||
|
Figura 4. Cts de amplificación de ORF1, ORF2, Rv2024 y SigA en RT-PCR en tiempo real.
|
|||
|
|
|||
 |
Figura 6. Anova de ACts de ORF1, ORF2 y Rv2024 en RT-PCR en tiempo real, a lo largo de la curva de crecimiento. Los valores más altos de ACt corresponden a un menor nivel de expresión y viceversa (en comparación con el gen normalizador sigA).
|
||
|
CONCLUSIONES
Los hallazgos obtenidos por medio de este estudio permiten afirmar que los tres marcos de lectura abiertos (ORF) predichos por el análisis bioinformático de la región RvD1 de Mycobacterium bovis: ORF1, ORF2 y Rv2024, sí se transcriben y, al parecer, lo hacen de manera constitutiva, ya que se detectó la presencia de estos transcritos por la técnica de RT-PCR en tiempo real, a partir del RNA total extraído a partir de todos los puntos de la curva de crecimiento normal.
AGRADECIMIENTOS
Los autores agradecen a las doctoras Clarissa Granja y Marcela Salazar, por sus aportes a este proyecto y a Colciencias por su financiación. Contrato RC 048-2002. No 6570-05-11417
BIBLIOGRAFÍA
Gordon, S. V., Brosch, R.; Billault, A.; Garnier, T.; Eiglmeier, K.; Cole, S. T. 1999. Identification of variable regions in the genomes of tubercle bacilli using bacterial artificial chro-mosome arrays. Molecular Microbiology. 32: 643-655.
Hu, Y.; Coates, A. 1999. Transcription of two sigma 70 homologue genes, sigA and sigB, in stationary phase Mycobacterium tuberculosis. Journal of Bacteriology. (181)1: 469-476.
O'Reilly, L. M. 1995. The epidemiology of Mycobacterium bovis infection in animals and man: a review. Tubercle and Lung Diseases. 76; 1-46.
Rodríguez, J. G.; Mejía, G. A., Del Portillo, P.; Patarroyo, M. E.; Murillo, L. A. 1995. Species-specific identification of Mycobacterium bovis by PCR. Microbiology. 141: 2131-2138.
|
|||
|
Figura 5. ACts de ORF1, ORF2 y Rv2024 en RT-PCR en tiempo real. Los valores más altos de ACt corresponden a un menor nivel de expresión y viceversa (en comparación con el gen norma-lizador sigA).
|
|||
|
(p < 0.001). Mientras que ORF2 y Rv2024 se transcribían en la misma cantidad (p > 0,05) (figura 6).
El hecho de que el ORF1 se transcriba en mayor proporción a lo largo de la curva de crecimiento, puede deberse a que éste es un gen independiente de ORF2 y de Rv2024, los cuales al parecer podrían transcribirse en forma de operón, en razón a su similitud en los niveles y en la tendencia de la cinética de transcripción. El paso siguiente será determinar si en realidad ORF1, ORF2 y Rv2024 se traducen a proteínas y, de ser así, si lo hacen bajo una organización en forma de operón o si son ORF independientes que se transcriben bajo el control de sus propios promotores. Los resultados de esta investigación permitirán más adelante determinar la función que desempeña la región RvD1 de M. bovis, su posible papel en la patogénesis y en la interacción huésped-patógeno en la tuberculosis bovina y humana, o su utilización para la generación de nuevos inmunógenos y métodos diagnósticos o como blanco de antibióticos.
|
|||
|
|
|||
|
66
|
|||
|
|
|||
Cómo citar
APA
Tibatá R., V. M., González, C. E., Rodríguez, J. G. & del Portillo, P. (2004). Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis. Revista Colombiana de Biotecnología, 6(2), 62–66. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527
ACM
[1]
Tibatá R., V.M., González, C.E., Rodríguez, J.G. y del Portillo, P. 2004. Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis. Revista Colombiana de Biotecnología. 6, 2 (jul. 2004), 62–66.
ACS
(1)
Tibatá R., V. M.; González, C. E.; Rodríguez, J. G.; del Portillo, P. Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis. Rev. colomb. biotecnol. 2004, 6, 62-66.
ABNT
TIBATÁ R., V. M.; GONZÁLEZ, C. E.; RODRÍGUEZ, J. G.; DEL PORTILLO, P. Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis. Revista Colombiana de Biotecnología, [S. l.], v. 6, n. 2, p. 62–66, 2004. Disponível em: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527. Acesso em: 26 mar. 2026.
Chicago
Tibatá R., Víctor Manuel, Clara Eugenia González, Juan Germán Rodríguez, y Patricia del Portillo. 2004. «Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis». Revista Colombiana De Biotecnología 6 (2):62-66. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527.
Harvard
Tibatá R., V. M., González, C. E., Rodríguez, J. G. y del Portillo, P. (2004) «Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis», Revista Colombiana de Biotecnología, 6(2), pp. 62–66. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527 (Accedido: 26 marzo 2026).
IEEE
[1]
V. M. Tibatá R., C. E. González, J. G. Rodríguez, y P. del Portillo, «Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis», Rev. colomb. biotecnol., vol. 6, n.º 2, pp. 62–66, jul. 2004.
MLA
Tibatá R., V. M., C. E. González, J. G. Rodríguez, y P. del Portillo. «Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis». Revista Colombiana de Biotecnología, vol. 6, n.º 2, julio de 2004, pp. 62-66, https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527.
Turabian
Tibatá R., Víctor Manuel, Clara Eugenia González, Juan Germán Rodríguez, y Patricia del Portillo. «Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis». Revista Colombiana de Biotecnología 6, no. 2 (julio 1, 2004): 62–66. Accedido marzo 26, 2026. https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527.
Vancouver
1.
Tibatá R. VM, González CE, Rodríguez JG, del Portillo P. Análisis transcripcional de la región genética RvD1 de Mycobacterium bovis. Rev. colomb. biotecnol. [Internet]. 1 de julio de 2004 [citado 26 de marzo de 2026];6(2):62-6. Disponible en: https://revistas.unal.edu.co/index.php/biotecnologia/article/view/527
Descargar cita
Visitas a la página del resumen del artículo
678
Descargas
Los datos de descargas todavía no están disponibles.
Licencia
Derechos de autor 2004 Revista Colombiana de Biotecnología

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Esta es una revista de acceso abierto distribuida bajo los términos de la Licencia Creative Commons Atribución 4.0 Internacional (CC BY). Se permite el uso, distribución o reproducción en otros medios, siempre que se citen el autor(es) original y la revista, de conformidad con la práctica académica aceptada. El uso, distribución o reproducción está permitido desde que cumpla con estos términos.
Todo artículo sometido a la Revista debe estar acompañado de la carta de originalidad. DESCARGAR AQUI (español) (inglés).















