



Published
Agronomic and physiological evaluation of eight cassava clones under water deficit conditions.
Evaluación agronómica y fisiológica de ocho clones de yuca sometidos a condiciones de déficit hídrico.
DOI:
https://doi.org/10.15446/rfnam.v73n1.75402Keywords:
Drought, Gas exchange, Growth, Manihot esculenta, Productivity (en)Sequía, Intercambio gaseoso, Crecimiento, Manihot esculenta, Productividad (es)
Cassava (Manihot esculenta Crantz) is an important crop in Africa, Asia, Latin America, and the Caribbean. This species grows adequately in drought conditions and is ideal to ensure food safety in marginal environments, such as soils with low fertility and low rainfall conditions. Complementary irrigation practices can be used, or genotypes with good yield potential can be identified against these conditions to enhance productivity in those environments. For this reason, this research aimed to study several physiological mechanisms and agronomical characteristics in eight cassava clones under prolonged water deficit. The experimental design was a split-plot where the principal plot was composed of three irrigation treatments (7, 15, and 21 days without irrigation, DWI) and the secondary plot by the eight clones. The results showed that cassava plants against drought conditions (21 DWI) decreased their growth, foliar expansion, and partially closed the stomata, avoiding water losses through transpiration. The variables of the net assimilation rate of CO2, yield, and biomass accumulation were not affected by the maximum drought period. The best clones under drought were Guajira, Guajira 3, Guajira 4, Concha Rosada, and MeVen 77-1. The frequency of irrigation 15 DWI increased the variables of gas exchange and vegetative growth. This behavior is due to the ability to tolerate complex conditions through morphological and physiological mechanisms, among them, long life leaf, stomatal control, and high photosynthetic potential.
La yuca (Manihot esculenta Crantz) es un cultivo importante en África, Asia, América Latina y el Caribe. Esta especie crece adecuadamente en condiciones de sequía y es ideal para garantizar la seguridad agroalimentaria en ambientes marginales, tales como suelos con baja fertilidad y bajas precipitaciones. Para ampliar la productividad en tales ambientes, se pueden utilizar prácticas de riego complementario o identificar genotipos con buen potencial de rendimiento frente a estas condiciones. Por ello, se planteó este experimento con el objetivo de estudiar algunos mecanismos fisiológicos y características agronómicas en ocho clones de yuca en condiciones de déficit hídrico prolongado. Se realizó un diseño experimental en parcelas divididas, donde la parcela principal estuvo conformada por tres niveles de riego (7, 15 y 21días sin riego, DSR) y la parcela secundaria por ocho clones de yuca. Los resultados muestran que las plantas de yuca frente a condiciones de sequía (21 DSR) disminuyeron su crecimiento, expansión foliar y cerraron parcialmente los estomas, evitando las pérdidas de agua por transpiración, sin afectar las variables de tasa de asimilación neta de CO2, rendimiento y acumulación de biomasa. Los clones que sobresalieron en este tratamiento fueron Guajira, Guajira 3, Guajira 4, Concha Rosada y MeVen 77-1. La frecuencia de riego 15 DSR afectó positivamente las variables de intercambio gaseoso y crecimiento vegetativo. Este comportamiento se debe a la capacidad de tolerar condiciones complejas mediante mecanismos morfológicos y fisiológicos, entre ellos, vida útil de la hoja, control estomático y alto potencial fotosintético.
Recibido: 8 de octubre de 2018; Aceptado: 23 de agosto de 2019
ABSTRACT
Cassava (Manihot esculenta Crantz) is an important crop in Africa, Asia, Latin America, and the Caribbean. This species grows adequately in drought conditions and is ideal to ensure food safety in marginal environments, such as soils with low fertility and low rainfall conditions. Complementary irrigation practices can be used, or genotypes with good yield potential can be identified against these conditions to enhance productivity in those environments. For this reason, this research aimed to study several physiological mechanisms and agronomical characteristics in eight cassava clones under prolonged water deficit. The experimental design was a split-plot where the principal plot was composed of three irrigation treatments (7, 15, and 21 days without irrigation, DWI) and the secondary plot by the eight clones. The results showed that cassava plants against drought conditions (21 DWI) decreased their growth, foliar expansion, and partially closed the stomata, avoiding water losses through transpiration. The variables of the net assimilation rate of CO2, yield, and biomass accumulation were not affected by the maximum drought period. The best clones under drought were Guajira, Guajira 3, Guajira 4, Concha Rosada, and MeVen 77-1. The frequency of irrigation 15 DWI increased the variables of gas exchange and vegetative growth. This behavior is due to the ability to tolerate complex conditions through morphological and physiological mechanisms, among them, long life leaf, stomatal control, and high photosynthetic potential.
Keywords:
Drought, Gas exchange, Growth, Manihot esculenta, Productivity.RESUMEN
La yuca (Manihot esculenta Crantz) es un cultivo importante en África, Asia, América Latina y el Caribe. Esta especie crece adecuadamente en condiciones de sequía y es ideal para garantizar la seguridad agroalimentaria en ambientes marginales, tales como suelos con baja fertilidad y bajas precipitaciones. Para ampliar la productividad en tales ambientes, se pueden utilizar prácticas de riego complementario o identificar genotipos con buen potencial de rendimiento frente a estas condiciones. Por ello, se planteó este experimento con el objetivo de estudiar algunos mecanismos fisiológicos y características agronómicas en ocho clones de yuca en condiciones de déficit hídrico prolongado. Se realizó un diseño experimental en parcelas divididas, donde la parcela principal estuvo conformada por tres niveles de riego (7, 15 y 21 días sin riego, DSR) y la parcela secundaria por ocho clones de yuca. Los resultados muestran que las plantas de yuca frente a condiciones de sequía (21 DSR) disminuyeron su crecimiento, expansión foliar y cerraron parcialmente los estomas, evitando las pérdidas de agua por transpiración, sin afectar las variables de tasa de asimilación neta de CO2, rendimiento y acumulación de biomasa. Los clones que sobresalieron en este tratamiento fueron Guajira, Guajira 3, Guajira 4, Concha Rosada y MeVen 77-1. La frecuencia de riego 15 DSR afectó positivamente las variables de intercambio gaseoso y crecimiento vegetativo. Este comportamiento se debe a la capacidad de tolerar condiciones complejas mediante mecanismos morfológicos y fisiológicos, entre ellos, vida útil de la hoja, control estomático y alto potencial fotosintético.
Palabras clave:
Sequía, Intercambio gaseoso, Crecimiento, Manihot esculenta, Productividad.Cassava is a tropical root crop and represents, due to its high content of starch, a basic daily source of dietary energy for more than 500 million people in Africa, Asia and Latin America (FAO, 2012). It has the ability to develop under complicated conditions on marginal lands. It is mostly grown on small-scale farms, usually intercropped with other vegetables and plantation crops. The roots (1-8 kg) mostly used for human consumption are processed into a wide variety of traditional foods. The cultivated cassava varieties contain cyanogenic glycosides (linamarin and lotaustralin) in concentrations that range 6-370 mg kg-1. It releases the toxic HCN upon the rupturing of the cells (e.g., during cutting, peeling, and processing) owing to the action of indigenous enzymes also present in the cassava root (Rawell and Kroll, 2003). The demand in the cultivation of cassava (Manihot esculenta Crantz) is increasing on the production of processed foods, animal feed, bioethanol, starch, and its derivatives (FAO, 2012).
However, loss in the yield of crops such as cereals, legumes, tubers and root crops due to the drought incidents borne out of global climate change are evidently demonstrated in several regions of the world (Daryanto et al., 2017). In tuber crops (potato and cassava), the storage root initiation is more sensitive to drought stress than vegetative developmental stages (Daryanto et al., 2017).
In Latin America, approximately 45% of the total area of cassava cultivation comes from areas with low water availability or where rains are sporadic (Tofiño et al., 2008). Despite this, the cassava plant is susceptible to drought at the beginning of the phenological stages because the water deficit causes a decrease in the vegetative growth, production, and quality of the roots. The drought is an important factor that directly infIuences various physiological processes of plants since water stress increases the overall resistance water vapor diffusion, by closing the stomata and reducing respiration and CO2 supply for photosynthesis (Nogueira, 1997).
In Venezuela, it is one of the crops within the sector of roots and tubers with the highest consumption per capita a year (8 kg) (Fedeagro, 2013).
At present, the information in Venezuela on the water requirements of cassava cultivation is limited. Current experience suggests that the plant requires moisture for sprouting and establishment of the crop, increasing its demand between the fourth and fifth months after planting (León et al., 2016, 2018). It has also been estimated that very frequent irrigation can cause excessive growth of the aerial part, reducing the production of radical biomass.
Therefore, the objective of this work was to evaluate eight cassava clones physiologically and agronomically against water deficit conditions in order to identify the cultivars with the best response and the appropriate irrigation frequencies in Maracay, Venezuela.
MATERIALS AND METHODS
Study location
The study was conducted in the Experimental Field of the National Center for Agricultural Research (CENIAP), Maracay, Aragua State, Venezuela, located in a Tropical Premontane Dry Forest zone, at 10°17'14" N and 67°36'02" W, and 480 m.a.s.l. The precipitation during the whole crop cycle was 1,208 mm.
The experiment was developed in a plot with loam-silty texture soil; the chemical analysis of the plot for phosphorus was 57 mg kg-1, potassium 111 mg kg-1, calcium 600 mg kg-1, magnesium 21 mg kg-1, organic matter 2.76%, pH 6.1 and hydraulic conductivity 0.13 mg kg-1 (Ceniap soil-water-plant analysis service unit).
Vegetal material
Eight clones of cassava were studied (Guajira, Guajira 1, Guajira 3, Guajira 4, MeVen 77-3, MeVen 77-1, Concha Rosada, and Bolívar 32), previously selected according to their favorable potential behavior against drought.
Description and design of the experiment
Under field conditions, an experimental trial was established with eight cassava clones, planted at 1 m between plants and 1 m between rows, three rows per genotype, three repetitions and six plants per row, guaranteeing irrigation and also the establishment and budding of the crop until the first 15 days. Irrigation was carried out by gravity, and water stress was applied by irrigation frequencies, which were made up of treatments 7, 15, and 21 days without irrigation (DWI).
Fertilization was applied at the time of sowing with a complete fertilizer (10-20-20 NPK) using a dose of 50 g per plant and was re-fertilized with urea three months after sowing with 50 g per plant. Weed control was manual and uniform throughout the plantation. During the investigation, there were no pests and diseases.
A design was made in split-plots, whose main plot consisted of three irrigation management (7, 15, and 21 days without irrigation, DWI) and the secondary plot with eight clones, with four replications. The clones were subjected to water stress from 21 to 111 days after sowing, at which time the dry period ended (precipitation greater than half of the potential evapotranspiration). The experimental unit (EU) consisted of irrigation treatments and clones, represented by 18 m2 per EU (18 plants per EU). The data were analyzed using an analysis of variance and Tukey's mean test, with a level of significance of 5%, using the Infostat program.
Physiological and agronomic evaluation
The evaluated variables were measured in weeks 7 and 17, at which time the three irrigation levels coincided.
Degree of greenness (spad). It was evaluated with the chlorophyll meter Spad 502 plus Minolta, taking the fifth fully expanded leaf, apex-base sense, in two plants and two leaves per plant for each experimental unit.
Photosynthesis (μmol CO 2 m -2 s -1 ), stomatal conductance (mmol CO 2 m -2 s -1 ), and transpiration (mmol H 2 O m -2 s -1 ). These variables were measured from 11:00 to 13: 00 h. An infrared gas analyzer equipment CI-340 Photosynthesis Systems (CID Inc, Washington, USA) was used, taking the fifth fully expanded leaf, apex-base sense, in two plants and two leaves per plant for each experimental unit.
Xylematic hydric potential (MPa). This variable was measured from 11:00 to 13:00 h. The last fully expanded sheet was taken, previously covered for approximately half an hour in the plant with aluminum foil bags, to be then taken to the pressure chamber according to the methodology described by Scholander et al. (1965). Two leaves were evaluated per clone and experimental unit.
Foliar area (cm 2 ). The fifth leaf, fully expanded, in the apex-base direction, was cut into three plants and taken to the laboratory, where they were then scanned and processed with the ImageJ program (Rincón Guerrero et al., 2012).
Number of leaves. The number of leaves of two plants of the central thread per experimental unit was counted, from the basal part of the plant to the apical stem.
Plant height (cm). This variable was measured in two plants of the central thread per experimental unit from the base to the apex of the stem.
Fresh aerial biomass (g). The aerial part was weighed in two plants of the central thread in the ninth month after the crop was established.
Dry biomass (g). The amount of dry matter per stem and root was measured during the ninth month. One plant of each clone was cut in each EU and taken to the laboratory. The stem and root samples were weighed once they were dried in an oven at 60 °C for 72 h.
Yield (g). Three plants of the central thread were evaluated in the ninth month, determining the following variables: number and mass of commercial roots (roots greater than 20 cm long and greater than 4 cm wide) and non-commercial roots.
RESULTS AND DISCUSSION
Physiological characterization of cassava clones
Degree of greenery (Spad). This variable did not show significant effects for the interactions and neither for the effects of the irrigation. However, there were significant differences between clones at the seventh week of evaluation in which the clone Guajira 3 stood out, while in week 17, the clone Guajira 4 showed the greatest value. This genotype was one of the worst in the behavior of this clone. Variable on the first evaluation date shows an unfavorable response and then adapts to the conditions of the experiment and manages to maintain higher values in this parameter (Table 1).
Table 1: Degree of greenness (spad) in eight clones of cassava in weeks 7 and 17 of evaluation.

On the other hand, it is possible that the clones against different water supplies maintained similar chlorophyll content, due to the strong association between the degree of greenness and the chlorophyll content (Barbosa, 2013).
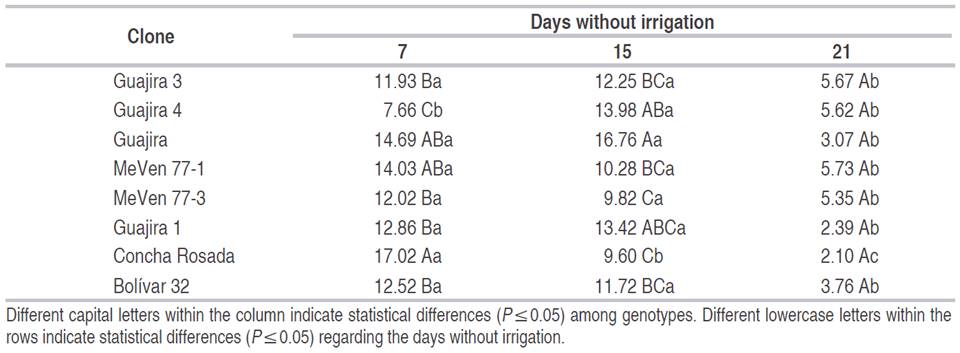
Rates of photosynthesis, transpiration, and stomatal conductance. For the photosynthesis variable, the irrigations did not show significant differences in week 17 of evaluation, but week 7 showed significant changes among the clones evaluated, highlighting the treatment 7 DWI Concha Rosada with 17.02 μmol CO2 m-2 s-1, 15 DWI Guajira with 16.76 μmol CO2 m-2 s-1, and Guajira 4 with 13.98 μmol CO2 m-2 s-1, respectively. Values that exceeded more than 14 μmol CO2 m-2 s-1 to the treatment present the worst behavior for this variable (21 DWI and 15 DWI Concha Rosada) (Table 2).
Table 2: Photosynthesis rate (μmol CO2 m-2 s-1) in week 7 of evaluation in three irrigation levels and eight cassava clones.
Same as the photosynthesis variable, the irrigations did not show significant differences for the stomatal conductance variable in weeks 7 and 17, with average values of 261 and 141 mmol CO2 m-2 s-1, respectively. This suggests that watering with a frequency of 21 days is sufficient to maintain adequate levels for these parameters (stomata partially open) since these were equally favored with 7 DWI, so that farmers can save water and make efficient use of this resource. This is explained because the response of cassava plants under conditions of abiotic stress is modulated by stomatal functions that allow reducing the loss of water by transpiration and by factors such as the canopy architecture and leaves orientation (León et al., 2016, San José and Mayobre, 1982).
In this regard, De Tafur (2002) found significant differences between cultivars and environments for the variable rate of assimilation of CO2, as well as a significant correlation between the same variable and dry root yield. This suggests that the rate of assimilation of CO2 is an important parameter to identify superior genotypes in the face of water stress due to drought. The rate of photosynthesis was 27 and 12 μmol CO2 m-2 s-1 for the dry and semi-arid environment, respectively. El-Sharkawy and De Tafur (2007) reported a similar trend, they refered to transpiration, values of 1 to 3 mol H2O m-2 s-1 in environments without stress due to drought, while in semiarid environments they reached values from 0 to 2 mmol H2O m-2 s-1.
The treatments that exceeded in the variable photosynthesis (7 DWI Concha Rosada and 15 DWI Guajira). They also exceeded in the transpiration rates in 1.83 and 1.75 mmol H2O m-2 s-1 to the treatment with 21 DWI, this treatment causes partially close of the stomata to avoid water losses (Table 3). On 17 DWI, Guajira 1 had similar behavior for all irrigation treatments, an outstanding characteristic of this clone against drought conditions. Even so, the high CO2 concentration reduced the negative effect of drought on transpiration, water use efficiency, all growth measures and harvest index, which gives indications that in the future, the crop could have different behavior with low rainfall and high concentrations of CO2 (Cruz et al., 2018).
Table 3: Transpiration (mmol H2O m-2 s-1) measured in weeks 7 and 17 of the evaluation in three irrigation levels and eight cassava clones.

On the other hand, León et al. (2014) found a different behavior for the variables of transpiration, stomatal conductance, and photosynthesis, in which no differences were observed between clones during the crop cycle, with values ranging between 1.27-1.61 mmol H2O m-2 s-1; 94.63-138.4 mmol CO2 m-2 s-1 and 16.63 μmol CO2 m-2 s-1, respectively.
Xylematic water potential. This variable had no significant interactions between factors and no statistically significant differences between the clones and irrigation factors for both evaluation dates. However, the values oscillated between -0.75 and -1.34 MPa, which indicates that the plants showed slight stress in the environmental conditions in which they were sown. Taiz and Zeiger (2006) indicate that the highest stresses are reached when the plants have water potentials less than -2 MPa.
In this aspect, El-Sharkawy (2007) showed that the leaf water potential in cassava suffers little variation with water stress with values ranging between -1.3 to -0.4 MPa. The capacity of cassava to maintain its water potential is due to the stomata closure and, therefore, to a considerable reduction of the stomatal conductance, which prevents the loss of water. Cassava plants conserve water under conditions of prolonged drought stress through a restriction in the formation of new leaves, production of smaller leaves, and leaf fall (Alves and Setter, 2004).
Similarly, in a study evaluating the leaf water potential in treatments without and with soil water stress in eight clones, they found values of -0.98 to -1.2 MPa and -0.97 to -1.19 MPa, respectively, without significant differences among clones evaluated, attributing these results to stomatal control to avoid water losses (El-Sharkawy, 2007; León et al., 2016).
Contrary to the above, San Jose and Mayobre (1982) observed differential changes in stomatal conductance and, therefore, in transpiratory rates, as well as a notable change in water potentials of leaves maintained by adequate turgor pressure.
Agronomic characterization of cassava clones
Leaf area. In none of the two evaluation dates, this variable showed significant effects of the clone interaction. In the same way it happened in week 7 for the irrigation factor, but it was significant in week 17, reaching the highest values (159.9 cm2) with the irrigation level of 7 DWI, then 15 DWI with 135.13 and 21 DWI with 118.9 cm2, respectively, since the greater frequency of water supply caused the plants to produce larger leaves. The lower frequency irrigation (21 DWI) caused a decrease for this variable of approximately 41.01 cm2 compared to 7 DWI, with the tendency to have the same behavior in the net assimilation rate variable of CO2.
In this aspect, San José and Mayobre (1982) indicated that the increase in the assimilation apparatus and in the assimilation rate might be due to leaf expansion, but in terms of specific leaf area. Likewise, the clone factor varied significantly in both evaluation dates, reaching the highest leaf area in week 7, the genotype Guajira 3 with 73.39 cm2, while in week 17, they were Guajira and Concha Rosada with 216.99 and 191.25 cm2, respectively. The tendency in time was to increase the leaf area; this last evaluation date (week 17) is approximately the time when this species reaches its maximum vegetative growth potential. This is possible because foliar expansion depends above all on cell expansion; the principles that govern those processes are similar.
Similar behavior is observed by Alves and Setter (2004), in which at 15 days after causing stress, the leaf area decreased from 200 cm2 in control to approximately 80 cm2 in the clones to which the volume of water was reduced. On the other hand, León et al. (2014) observed significant differences for the clone and evaluated months and interactions of the clones, highlighting the La Reina and Concha Rosada varieties, with values of 261 cm2 at the fourth month and 61 cm2 at the eighth month of evaluation. In this sense, the leaf area and the number of leaves decreased in the treatment to which the lower water demand of the cassava crop was supplied (25% of the field capacity of the soil). This reduction is attributed to the decrease and completion of production of new leaves, as well as foliar abscission under drought conditions (Helal et al., 2013).
Plant height. This variable was strongly infIuenced by the interaction irrigation of clones in week 17, whose highest rates were observed in all treatments of irrigation in the clones Guajira, Guajira 3, and Concha Rosada (Table 4), genotypes that can be recommended for seed production in dry environments. In this same sense, it is important to point out that clone MeVen77-3 had the lowest growth in the three irrigation treatments, which could be attributed to a low size characteristic of the genotype (Table 4).
Table 4: Plant height (cm) in week 17 of evaluation in three irrigation levels and eight cassava clones

Irrigation levels showed that the heights on 7 and 15 DWI were significantly higher (65.6 and 58.8 cm, respectively) in week 7 because they caused adequate moisture in the soil, allowing the plants to show an increase in the vegetative development.
On the other hand, the clones varied only for weeks 7 and 17 of evaluation, with the tendency to have the highest rates in clones Guajira 3, Guajira, and Concha Rosada (Table 5). In this regard, Montaldo (1996) show that plant height is one of the biggest indicators for agronomic management. In this sense, plants with intermediate and high heights are preferred, with values for plant height and first branching above 150 and 100 cm, respectively. Different investigations point out differences for this variable between different genotypes and evaluation dates, which indicates that it is a character strongly infIuenced by the genotype and environment (León et al., 2013, 2014; Barbosa, 2013; Rós et al., 2011; Marín et al., 2008). Otherwise, refer Albuquerque et al. (2009), who, in their research, did not obtain important differences between the clones and between the dates of measurement for the plant height variable with averages of 143 cm at seven months of culture and 208 cm in 13 months.
Table 5: Agronomic variables Leaf area (LA), number of leaves (NL), plant height (PH), fresh stem weight (FSW), and dry stem weight (DSW) on the aerial part of the plant in eight cassava clones.

Number of leaves. This variable showed significant effects neither for the clone interactions nor for the irrigation factor. On the other hand, the clones did not vary significantly in the seventh week of evaluation. In the opposite case, it occurred in week 17, significantly varying with the highest value being the Guajira 3 clone and with the worst behavior MeVen 77-1 (Table 5), which could translate into Guajira 3 being faster growing and, therefore, of more precocious behavior than the rest. In this aspect, San José and Mayobre (1982) reported in the Cuban clone, under savanna conditions, that partition mechanisms in the plant promoted the growth of the canopy and the development of new leaves with a distribution over time of the assimilated between leaves and other organs. Similarly, Ledent (2002) reports that, during a period of prolonged drought, the crop produces fewer leaves, and these are smaller and increases the retention of the already expanded leaves. When the drought ends, the plant begins to recover quickly and produces new leaves at the expense of the starch of the roots and stems. In the same order, Suárez and Mederos (2011) indicate that the total number of leaves produced by the plant and its longevity are varietal characteristics, deeply infIuenced by environmental conditions. In this regard, León et al. (2014) and Vandegeer et al. (2013) indicated that the loss of leaves in the cassava crop is a strategy of the plant to economize water. In spite of this, in their investigation, there were no significant differences between clones nor between interactions with the months, showing values of the number of leaves per plant that ranged from 181 to 305. Likewise, the number of leaves in cassava was significantly dropped in plants grown under water deficit conditions. At 150 DAP (120 days water withholding), number of leaves in water-deficit stressed plants was 15.34% lower than that in under well-watered plants (WW). It was evidently demonstrated that the number of fallen leaves was promoted by an adaptive period of water deficit, leading to decrease LAI parameter. LAI in both WW and water-stressed (WS) conditions increased in relation to the growing period. In addition, the reduction percentage of LAI in WS cassava at 150 DAP was 34.32% lesser than WW plants (Pipatsitee et al., 2019).
Fresh and dry biomass aerial and root. These variables did not present significant effects on the interactions. The same happened with the irrigation factor; however, the trend observed during the experiment was the decrease in the variables associated with vegetative growth on 21 DWI and, therefore, a decrease in fresh and dry aerial biomass.
On the other hand, the clones had a differential behavior (Table 5), in which Guajira 3 reached the highest value for aerial biomass, as well as the highest transport rates. Otherwise, it occurred with clones MeVen 77-1, Guajira 1, Guajira 4, Bolívar 32 and MeVen 77-3, which presented the lowest values for these variables, product of which during the cycle they reached the lowest vegetative growth rates, and therefore the least possibility of transferring the assimilated to the aerial part.
With regard to dry stem biomass, the genotypes that stood out were Guajira 3, Guajira, Concha Rosada, and Bolívar 32 (Table 5), which possibly was due to the fact that it was the clone that stood out in most of the physiological variables and agronomic, favoring a positive stem-root relationship towards all the elements that make up the plant. San José and Mayobre (1982) explain this behavior in a cassava cultivar in the humid savanna season, where they observed that a large part of the assimilated production was not due to the demand of the storage roots, but was due to the morphological development, production of new leaves and the canopy. Similarly, Barbosa (2013), El-Sharkawy et al. (2012) and León et al. (2018) observed a positive and significant correlation between the height of the cassava plant with the diameter of the canopy, diameter of the root and stem, aerial weight of the plants and root dry weight, which indicates that increases in vegetative parts, have a significant effect on cassava dry matter production. In this regard, Olasanmi (2010) observed that the genotypes that stood out against drought conditions were those that maintained a robust development in the roots, while the clones with the worst behavior showed a better development of other organs in favor of the roots.
Similar to this study, Helal et al. (2013) and El-Sharkawy (2007) observed that water stress reduced weight plant compared with the control with irrigation and with significant differences between stress irrigation treatments. The reduction in the weight plant was associated with a decrease in cell growth and foliar senescence under stress conditions (Bhatt and Srinivasa, 2005). This is attributed to thef reduction of the foliar area as a strategy of the plant to avoid water losses in the leaves due to a reduction in the photosynthetically active area (León et al., 2016).
Yield (Number of total roots, number of commercial roots, and weight of total and commercial roots). The variables associated with performance did not show statistically significant effects for the interactions between the evaluated factors nor for the irrigation factor. This could be attributed to the fact that these clones were adapted to drought conditions so that the plants were adjusted physiologically to avoid damages caused by water stress without detriment to the production variables.
In the same aspect, it can be pointed out that the clones varied in this investigation; the best cultivar clone evaluated is Guajira 3 (Table 6). The clone Guajira, despite having eight roots in total, they did not manage to form with a commercial size, decreasing the weight of commercial roots. This genotype was the fourth with the highest number of roots, so it could be a genotype with the ability to exploit its maximum yield potential in other environmental conditions, transferring its assimilated to the roots in the filling stage. It is important to point out that in general, all the evaluated clones presented the potential of yield, since they surpass the national yield (1 to 3 kg per plant) (Fedeagro, 2013), except the clone Guajira.
Table 6: Behavior of agronomic variables root dry weight (RDW), root fresh weight (RFW), weight of commercial root (WCR), weight of non-commercial root (WnCR), total number of roots (TNR), total number of commercial roots (TNCR) and total number of non-commercial roots in eight cassava clones.

Otherwise reported Caraballo and Velásquez (1997), in this case, the roots reached on average 2.9 kg per plant when the crop received at least 324 mm of water between the fourth and seventh month after sowing; when the availability of water was 156 mm, the weight of the roots reached 1.8 kg per plant. Also, several researchers observed on irrigation, clones and their interactions, that plants against drought decreased their yields, but some genotypes excelled against stressful conditions in different phenological phases, indicating that some genotypes adapted to dry environments can be selected, as well as materials with early development to avoid stress (Albuquerque et al., 2009; Barbosa, 2013; León et al., 2016; Rós et al., 2011;Vandegeer et al., 2013). Drought significantly decreased shoot dry weight, total root number, and root length by 84%, 30%, and 25%, respectively. A high adventitious root number was associated with increased shoot dry weight (r=0.44) under drought (Kengkanna et al., 2019).
Finally, it can be said that clone Guajira 3, in all irrigation treatments maintained high spad values, which could be associated with a high chlorophyll content, very important behavior, since, throughout the treatments with and without stress, it was possible to observe a superior behavior in the variables of photosynthesis and transpiration, which mean that their photoassimilates could be used efficiently for their vegetative growth (higher plant height and leaf area) and greater root yield, behavior that was contrary for the Bolívar 32 clone in the physiological and agronomic variables. Whereas, Concha rosada, despite not excelling in stress treatment (21 DWI), it was possible to observe higher rates of transpiration and photosynthesis at 7 DWI, which makes it a relevant clone in irrigation conditions or without stress by water deficit. About this aspect, leaf relative water content in cassava declined significantly upon a long period of water withholding, and regulated non-photochemical quenching, leading to chlorophyll degradation, reduced number of leaves and limited leaf area index and loss of storage root yield when compared with well-irrigated plants (Pipatsitee et al., 2019).
CONCLUSIONS
The clones interacted significantly with the irrigation in the variables of gas exchange and vegetative growth; so that the Guajira, Guajira 3, Guajira 4, Concha Rosada and MeVen 77-1 genotypes can be identified in seed production systems in dry environments.
The vegetative growth variables affected positively the fresh and dry stem-root biomass and yield, obtaining outstanding behavior the clones Guajira 3 and Guajira 1.
The treatment of 21 DWI negatively affected the vegetative growth of the cassava plants. However, the variables associated with gas exchange and yield did not vary in the irrigation treatments; therefore, it can be recommended for these experimental conditions to irrigate with a frequency of 21 days.
REFERENCES
References
Albuquerque JAA, Sediyama T, Silva AA, Sediyama CS, Alves JMA e Neto FA. 2009. Caracterização morfológica e agronômica de clones de mandioca cultivados no estado de Roraima. Revista Brasileira de Ciências Agrárias 4(4): 388-394. doi: 10.5039/agraria.v4i4a3
Alves AAC and Setter TL. 2004. Response of cassava leaf area expansion to water deficit: cell proliferation, cell expansion and delayed development. Annals of Botany 94(4): 605–613. doi: 10.1093/aob/mch179
Barbosa GM. 2013. Caracterização morfofisiológica de clones de mandioca em cândido sales-ba (Tesis Magister Scientarum). Universidad Estadual do Sudoeste da Bahia. Vitoria Da Conquista, Brasil. 114 p.
Bhatt RM and Srinivasa NK. 2005. Influence of pod load response of okra to water stress. Indian Journal of Plant Physiology 10(1): 54-59.
Caraballo L y Velásquez E. 1997. Respuesta de tres cultivares de yuca a diferentes condiciones hídricas y fechas de cosecha. Agronomía tropical 50(2): 267-284.
Cruz JL, LeCain DR, Alves AAC, Coelho Filho MA and Coelho EF. 2018. Elevated CO2 reduces whole transpiration and substantially improves root production of cassava grown under water deficit. Archives of Agronomy and Soil Science 64(12):1623-1634. doi: 10.1080/03650340.2018.1446523
Daryanto S, Wang L, Jacinthe PA. 2017. Global synthesis of drought effects on cereal, legume, tuber and root crops production: A review. Agricultural Water Management 179: 18-33. doi: 10.1016/j.agwat.2016.04.022
De Tafur SM. 2002. Capitulo 3 - Fisiología de la yuca (Manihot esculenta Crantz). En: Ceballos H and Ospina B (eds). La Yuca en el Tercer Milenio: Sistemas Modernos de Producción, Procesamiento, Utilización y Comercialización. CIAT, Cali. pp. 4-45.
El-Sharkawy MA. 2007. Physiological characteristics of cassava tolerance to prolonged drought in the tropics: Implications for breeding cultivars adapted to seasonally dry and semiarid environments. Brazilian Journal of Plant Physiology 19(4): 257-286. doi: 10.1590/S1677-04202007000400003
El-Sharkawy M and De Tafur SM. 2007. Genotypic and within canopy variation in leaf carbon isotope discrimination and its relation to short-term leaf gas exchange characteristics in cassava grown under rain-fed conditions in the tropics. Photosynthetica 45(4): 515-526. doi: 10.1007/s11099-007-0089-y
El-Sharkawy MA. 2012. Stress-tolerant cassava: The role of integrative ecophysiology-breeding research in crop improvement. Open Journal of Soil Science 2(2): 162-186. doi: 10.4236/ojss.2012.22022.
FAO. 2012. Base de datos. En: http://www.fao.org/statistics/databases/es/ ; consulta: junio 2012
Fedeagro. 2013. Base de datos. Confederacion de Asociaciones de Productores Agropecuarios. En : http: //www.fedeagro.org/produccion/ ; consulta: junio 2014
Helal NAS, Eisa SS and Attia A. 2013. Morphological and chemical studies on influence of water deficit on cassava. World Journal of Agricultural Sciences 9(5): 369-376. doi: 10.5829/idosi.wjas.2013.9.5.1801
Kengkanna J, Jakaew P, Amawan S, Busener N, Bucksch A and Saengwilai P. 2019. Phenotypic variation of cassava root traits and their responses to drought. Applications in Plant Sciences 7(4): e01238. doi: 10.1002/aps3.1238
León R, Polanco D, Zárraga P, Zambrano M, Ramos E, Perdomo D y Marín A. 2013. Caracterización morfológica y agronómica del banco de germoplasma de yuca (Manihot esculenta, Crantz) en el campo experimental de Fagro-UCV. Revista de la facultad de Agronomía de la UCV 39(2): 93-104.
León R, Pérez M, Gutiérrez M, Rodríguez A, Fuenmayor F y Marín C. 2014. Caracterización eco-fisiológica de cuatro clones de yuca (Manihot esculenta Crantz) en el Campo Experimental del INIA-CENIAP. Agronomía Tropical 64(1-2): 97-105.
León R, Pérez M, Fuenmayor F, Gutiérrez M, Rodríguez A, Rodríguez G y Marín C. 2016. Evaluación fisiológica y agronómica de clones promisorios de yuca (Manihot esculenta Crantz) sometidos a condiciones de estrés por sequía. Revista UNELLEZ de Ciencia y tecnología 34: 50-57
León R, Pérez M, Fuenmayor F, Rodríguez A, Rodríguez G y Marín C. 2018. Calidad de las raíces en cuatro clones de yuca (Manihot esculenta Crantz) y efecto del régimen de riego. Bioagro 30(1): 87-91
Ledent JF. 2002. Déficit hídrico y crecimiento de las plantas: Respuestas al déficit hídrico. Comportamiento morfo-fisiológico. Université Catholique de Louvai. Lovaina, Bélgica.
Marín A, Perdomo D, Albarrán JG, Fuenmayor F y Zambrano C. 2008. Evaluación agronómica morfológica y bioquímica de clones élites de yuca a partir de vitro plantas. Interciencia 33(5): 27-35.
Montaldo A. 1996. La yuca frente al hambre del mundo tropical. Primera Edición. Anauco, Maracay. 570 p.
Nogueira R. 1997. Expressões fisiológicas da aceroleira (Malpighia emarginata D.C.) em condições adversas (Tese Doutorado). Universidade Federal de São Paulo. São Carlos, Brasil. 207 p.
Olasanmi O. 2010. Cassava drought tolerance mechanisms re-visited: Evaluation of drought tolerance in contrasting cassava genotypes under water stressed environments. (Doctorate thesis). University of Ibadan. Ibadan, Nigeria. 150 p.
Pipatsitee P, Eiumnoh A, Praseartkul P, Ponganan N, Taota K, Kongpugdee S and Cham-Um S. 2019. Non-Destructive Leaf Area Estimation Model for Overall Growth Performances in Relation to Yield Attributes of Cassava (Manihot esculenta Cranz) under Water Deficit Conditions. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 47(3): 580-591. doi: 10.15835/nbha47311487
Rincón Guerrero N, Olarte Quintero MA y Pérez Naranjo JC. 2012. Determinación del área foliar en fotografías tomadas con una cámara web, un teléfono celular o una cámara semiprofesional. Revista Facultad Nacional de Agronomía Medellín 65(1): 6399-6405.
Rós AB, Silva ACS, de Araújo HS e Narita N. 2011. Crescimento, fenologia e produtividade de cultivares de mandioca. Pesquisa Agropecuária Tropical 41(4): 552-558. doi: 10.5216/pat.v41i4.11075
Rawel H and Kroll J. 2003. The importance of Cassava (Manihot esculenta Crantz) as the main staple food in tropical countries. Deutsche Lebensmittel-Rundschau. 99(3): 102-111.
San José JJ and Mayobre F. 1982. Quantitative growth relationship of cassava (Manihot esculenta Crantz). Crop development in a savanna wet season. Annals of Botany 50(3): 309-316. doi: 10.1093/oxfordjournals.aob.a086370
Scholander PF, Bradstreet ED, Hemmingsen EA and Hammel HT. 1965. Sap pressure in vascular plants: negative hydrostatic pressure can be measured in plants. Science 148(3668): 339–346. doi: 10.1126/science.148.3668.339
Suárez L y Mederos V. 2011 Apuntes sobre el cultivo de la yuca (Manihot esculenta Crantz). Tendencias actuales. Cultivos Tropicales 32(3): 27-35.
Taiz L y Zeiger E. 2006. Fisiología Vegetal. Universitat Jaume I. Castello de la Plana, España.
Tofiño A, Ceballos H y Romero M. 2008. Posibilidades de expansión del cultivo de yuca (Manihot esculenta Crantz) en el Caribe Seco Colombiano a partir de investigación multidisciplinaria. Actualidades Biológicas 30(88): 15-27.
Vandegeer R, Miller RE, Bain M, Gleadow RM and Cavagnaro TR. 2013. Drought adversely affects tuber development and nutritional quality of the staple crop cassava (Manihot esculenta Crantz). Functional Plant Biology 40: 195–200. doi: 10.1071/FP12179
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
CrossRef Cited-by
1. Amparo Rosero, Jorge-Iván Lenis, Rommel León, Hernando Araujo, Jorge García, Alfonso Orozco, Remberto Martínez, Martha Montes, Víctor De la Ossa, Carina Cordero, Sandra Salazar, Nelson Morante, Luis-Fernando Delgado, Hernán Ceballos. (2026). Enhancing Year-Round Cassava Production and Processing in Colombia Through Varieties with Stable Root Dry Matter Content. Plants, 15(3), p.489. https://doi.org/10.3390/plants15030489.
2. Ofelda Peñuelas Rubio, Leandris Argentel Martínez, Jorge Gonzalez Aguilera, Bruna Izabel Krewer, Jessica Ferrerira Diniz, Pedro Paulo Vilela Barros, José Aristeu Alfonso Junior, Rafael Felippe Ratke, Alan Mario Zuffo, Jaime Garatuza Payán. (2023). EFFECTS OF PLANTING POSITION AND Azospirillum brasilense APPLICATION ON THE SPROUT AND AGRONOMIC TRAITS OF CASSAVA (Manihot esculenta Crantz). Agricultura, Sociedad y Desarrollo, 20(2) https://doi.org/10.22231/asyd.v20i2.1415.
3. Oluwadamilare Eludire, Oluwaseun Faloye, Michael Alatise, Ayodele Ajayi, Philip Oguntunde, Tayo Badmus, Abayomi Fashina, Oluwafemi Adeyeri, Idowu Olorunfemi, Akinwale Ogunrinde. (2025). Evaluation of Evapotranspiration Prediction for Cassava Crop Using Artificial Neural Network Models and Empirical Models over Cross River Basin in Nigeria. Water, 17(1), p.87. https://doi.org/10.3390/w17010087.
4. Jimmy A. Ocaña-Reyes, Richard Paredes-Espinosa, Astrid Quispe-Tomas, Henry Díaz-Chuquizuta, Zoila L. Ore-Aquino, Alex I. Agurto-Piñarreta, W. Michel Paz Monge, Roiser H. Lobato-Galvez, José G. Ruiz Reyes, José W. Zavala-Solórzano, Hugo Alfredo Huamani Yupanqui, Giannfranco Egoávil-Jump, Ceila P. Lao Olivares. (2024). Tillage Systems Modify the Soil Properties and Cassava Physiology During Drought. Agronomy, 14(12), p.3041. https://doi.org/10.3390/agronomy14123041.
5. Charles Orek. (2025). Evaluation of cassava genotypes (Manihot esculenta Crantz) for drought tolerance and susceptibility under water deficit conditions. Journal of Agricultural, Food Science and Biotechnology, 3(1), p.48. https://doi.org/10.58985/jafsb.2025.v03i01.65.
6. Marcelo de Almeida Silva, Samara Zanetti, Hariane Luiz Santos, Laís Fernanda Melo Pereira. (2021). Nutritional status, yield components, and yield of cassava as influenced by phenological stages and water regimes. Journal of Plant Nutrition, 44(19), p.2912. https://doi.org/10.1080/01904167.2021.1927084.
7. Ridwan Diaguna, Suwarto, Edi Santosa, Arief Hartono, Gatot Pramuhadi, Nunung Nuryartono, Roza Yusfiandayani, Tri Prartono, Magdi Abdelhamid. (2022). Morphological and Physiological Characterization of Cassava Genotypes on Dry Land of Ultisol Soil in Indonesia. International Journal of Agronomy, 2022, p.1. https://doi.org/10.1155/2022/3599272.
8. Maltase Mutanda, Assefa B. Amelework, Nzumbululo Ndou, Sandiswa Figlan. (2025). Drought Stress in Cassava (Manihot esculenta): Management Strategies and Breeding Technologies. International Journal of Plant Biology, 16(4), p.112. https://doi.org/10.3390/ijpb16040112.
9. Sanket J. More, Kirti Bardhan, Velumani Ravi, Ratnakumar Pasala, Ashish K. Chaturvedi, Milan Kumar Lal, Kadambot H. M. Siddique. (2023). Morphophysiological Responses and Tolerance Mechanisms in Cassava (Manihot esculenta Crantz) Under Drought Stress. Journal of Soil Science and Plant Nutrition, 23(1), p.71. https://doi.org/10.1007/s42729-023-01127-4.
Dimensions
PlumX
Article abstract page views
Downloads
License
Copyright (c) 2020 Revista Facultad Nacional de Agronomía Medellín

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
The journal allows the author(s) to maintain the exploitation rights (copyright) of their articles without restrictions. The author(s) accept the distribution of their articles on the web and in paper support (25 copies per issue) under open access at local, regional, and international levels. The full paper will be included and disseminated through the Portal of Journals and Institutional Repository of the Universidad Nacional de Colombia, and in all the specialized databases that the journal considers pertinent for its indexation, to provide visibility and positioning to the article. All articles must comply with Colombian and international legislation, related to copyright.
Author Commitments
The author(s) undertake to assign the rights of printing and reprinting of the material published to the journal Revista Facultad Nacional de Agronomía Medellín. Any quotation of the articles published in the journal should be made given the respective credits to the journal and its content. In case content duplication of the journal or its partial or total publication in another language, there must be written permission of the Director.
Content Responsibility
The Faculty of Agricultural Sciences and the journal are not necessarily responsible or in solidarity with the concepts issued in the published articles, whose responsibility will be entirely the author or the authors.


















