



Published
Fruit preservation with bioethanol obtained from the fermentation of brewer’s spent grain with Saccharomyces carlsbergensis
Preservación de frutas con bioetanol obtenido a partir de la fermentación de cascarilla de cebada cervecera con Saccharomyces carlsbergensis
DOI:
https://doi.org/10.15446/rfnam.v73n3.85316Keywords:
Agro-wastes, Bioethanol, Fermentation, Fruit rottenness (en)Agro-residuos, Bioetanol, Fermentación, Podredumbre de frutos (es)
Downloads
Brewer’s Spent Grain (BSG) is renewable lignocellulosic biomass generated from the beer brewing process. It serves as a substrate for various biotechnological applications. BSG was used as the main substrate for bioethanol production with Saccharomyces carlsbergensis in submerged fermentation. Saccharification and fermentation studies were performed for the production of bioethanol. A sterilized fermenter was loaded with 50 g L-1 of BSG at 29±2 °C and an agitation speed of 180 rpm. pH was adjusted to 6.0 before the addition of 500 mL of yeast culture for 7 days under submerged and optimized conditions. The fermented product was concentrated using a rotary evaporator at 66±1 °C, and ethanol was qualitatively determined by the dichromate method. Bioethanol yield was 22%, with a specific gravity of 0.8 at 28 °C. Fourier-Transform Infrared Spectroscopy (FTIR) confirmed the presence of -CH3 stretch, -OHstretch and -CH2stretch in bioethanol. For the preservative test, Staphylococcus spp., Erwinia spp., Lactobacillus spp., Bacillus spp., Xanthomonas spp., Pseudomonas spp., Micrococcus spp. and Corynebacterium spp. were the bacteria isolated from fruits examined from different regions of Osun State. The genera of fungi isolated were Aspergillus, Colletotrichum, Penicillium, Fusarium, Alternaria, Rhizopus, Candida, Saccharomyces, Geotrichium and Pichia. Bioethanol produced from BSG inhibited the growth of microorganisms with zones of inhibition range from 7.0 mm to 11.5 mm, and thus, selected fruits were preserved. Hence, the fermentation technology of agro-industrial wastes with microorganisms can be adopted to convert waste biomass to useful resources.
Cascarilla de cebada cervecera (CCC) es una biomasa lignocelulósica renovable generada a partir del proceso de elaboración de la cerveza, que sirve como sustrato para diversas aplicaciones biotecnológicas. Se usó CCC como sustrato principal para la producción de bioetanol con Saccharomyces carlsbergensis en fermentación sumergida. Se realizaron estudios de sacarificación y fermentación para la producción de bioetanol, el fermentador esterilizado se cargó con 50 g L-1 de CCC a 29±2 °C y una velocidad de agitación de 180 rpm. El pH se ajustó a 6,0 antes de la adición de 500 mL de cultivo de levadura durante 7 días en condiciones sumergidas y optimizadas. El producto fermentado se concentró usando un evaporador rotatorio a 66±1 °C y el etanol se determinó cualitativamente por el método de dicromato. El rendimiento de bioetanol fue del 22% con un peso específico de 0,8 a 28 °C. La Espectroscopía Infrarroja por Transformada de Fourier (FTIR) confirmó la presencia de CH3, OH y CH2 en el bioetanol. Para el ensayo de preservación, Staphylococcus spp., Erwinia spp., Lactobacillus spp., Bacillus sp., Xanthomonas spp., Pseudomonas spp., Micrococcus spp. y Corynebacterium spp. fueron bacterias aisladas de frutas examinadas de diferentes regiones del estado de Osun. Los géneros de hongos aislados fueron Aspergillus, Colletotrichum, Penicillium, Fusarium, Alternaria, Rhizopus, Candida, Saccharomyces, Geotrichium y Pichia. El bioetanol producido a partir de CCC inhibió el crecimiento de microorganismos con zonas de inhibición comprendidas entre 7,0 mm y 11,5 mm conservando las frutas seleccionadas. Por lo tanto, se puede adoptar la tecnología de fermentación de desechos agroindustriales con microorganismos para convertir la biomasa residual en recursos útiles.
Recibido: 30 de marzo de 2020; Aceptado: 3 de agosto de 2020
ABSTRACT
Brewer's Spent Grain (BSG) is renewable lignocellulosic biomass generated from the beer brewing process. It serves as a substrate for various biotechnological applications. BSG was used as the main substrate for bioethanol production with Saccharomyces carlsbergensis in submerged fermentation. Saccharification and fermentation studies were performed for the production of bioethanol. A sterilized fermenter was loaded with 50 g L-1 of BSG at 29±2 °C and an agitation speed of 180 rpm. pH was adjusted to 6.0 before the addition of 500 mL of yeast culture for 7 days under submerged and optimized conditions. The fermented product was concentrated using a rotary evaporator at 66±1 °C, and ethanol was qualitatively determined by the dichromate method. Bioethanol yield was 22%, with a specific gravity of 0.8 at 28 °C. Fourier-Transform Infrared Spectroscopy (FTIR) confirmed the presence of -CH3 stretch, -OH stretch and -CH2stretch in bioethanol. For the preservative test, Staphylococcus spp., Erwinia spp., Lactobacillus spp., Bacillus spp., Xanthomonas spp., Pseudomonas spp., Micrococcus spp. and Corynebacterium spp. were the bacteria isolated from fruits examined from different regions of Osun State. The genera of fungi isolated were Aspergillus, Colletotrichum, Penicillium, Fusarium, Alternaria, Rhizopus, Candida, Saccharomyces, Geotrichium and Pichia. Bioethanol produced from BSG inhibited the growth of microorganisms with zones of inhibition range from 7.0 mm to 11.5 mm, and thus, selected fruits were preserved. Hence, the fermentation technology of agro-industrial wastes with microorganisms can be adopted to convert waste biomass to useful resources.
Keywords:
Agro-wastes, Bioethanol, Fermentation, Fruit rottenness.RESUMEN
Cascarilla de cebada cervecera (CCC) es una biomasa lignocelulósica renovable generada a partir del proceso de elaboración de la cerveza, que sirve como sustrato para diversas aplicaciones biotecnológicas. Se usó CCC como sustrato principal para la producción de bioetanol con Saccharomyces carlsbergensis en fermentación sumergida. Se realizaron estudios de sacarificación y fermentación para la producción de bioetanol, el fermentador esterilizado se cargó con 50 g L-1 de CCC a 29±2 °C y una velocidad de agitación de 180 rpm. El pH se ajustó a 6,0 antes de la adición de 500 mL de cultivo de levadura durante 7 días en condiciones sumergidas y optimizadas. El producto fermentado se concentró usando un evaporador rotatorio a 66±1 °C y el etanol se determinó cualitativamente por el método de dicromato. El rendimiento de bioetanol fue del 22% con un peso específico de 0,8 a 28 °C. La Espectroscopía Infrarroja por Transformada de Fourier (FTIR) confirmó la presencia de CH3, OH y CH2 en el bioetanol. Para el ensayo de preservación, Staphylococcus spp., Erwinia spp., Lactobacillus spp., Bacillus sp., Xanthomonas spp., Pseudomonas spp., Micrococcus spp. y Corynebacterium spp. fueron bacterias aisladas de frutas examinadas de diferentes regiones del estado de Osun. Los géneros de hongos aislados fueron Aspergillus, Colletotrichum, Penicillium, Fusarium, Alternaria, Rhizopus, Candida, Saccharomyces, Geotrichium y Pichia. El bioetanol producido a partir de CCC inhibió el crecimiento de microorganismos con zonas de inhibición comprendidas entre 7,0 mm y 11,5 mm conservando las frutas seleccionadas. Por lo tanto, se puede adoptar la tecnología de fermentación de desechos agroindustriales con microorganismos para convertir la biomasa residual en recursos útiles.
Palabras clave:
Agro-residuos, Bioetanol, Fermentación, Podredumbre de frutos.Wheat bran, rice bran, corn cob and wheat straw are examples of agricultural wastes available as carbon sources (Singh et al., 2012). Brewer's Spent Grain (BSG), a waste-product of the mashing process, is one of the initial operations in brewery industries to solubilize malt and grains to ensure adequate extraction of wort (Aliyu and Bala, 2011). BSG is available in larger quantities; approximately 85-90% of the total by-products generated during beer production; however, its central exploitation or disposition has been inadequate (Steiner et al., 2015). Several attempts have been made to use BSG in biotechnological processes, and this is achievable in various researches and industries by adopting solid-state fermentation (SFF) or submerged fermentation (SMF) since BSG contains basic nutrients required for microbial growth (Mussatto, 2014). BSG is a lignocellulosic material with 17% cellulose, 28% non-cellulosic polysaccharides, minerals, vitamins, proteins, amino acids, arabinoxylans, 28% lignin (Ivanova et al., 2017). Therefore, it has a perspective to be recycled and to become useful products. BSG has been utilized as a new and economical medium for the cultivation of microorganisms (Tan et al., 2020).
Agricultural residues are currently utilized for the production of bioethanol to decrease total dependence on forest woody biomass and continuous deforestation. Bioethanol is produced from different agro-wastes using some biotechnological methods such as fermentation with diverse microorganisms (Bušić et al., 2018). The use of carbon sources from renewable biomass is economical for the full exploitation of less expensive sources into the production of beneficial products (Saini et al., 2015). Hence, agro-industrial residues are an attractive alternative to costly raw materials. Bioethanol has been produced by converting sugars directly from BSG or indirectly through starch into alcohol via fermentation followed by distillation (Azhar et al., 2017). Ethanol produced from lignocellulosic biomass raises a global interest because it represents an excellent alternative to petroleum-derived energies and reduces food versus fuel confIict generated by first-generation ethanol (Awoyale and Lokhat, 2019; Prasad et al., 2019).
Ethanol is an excellent preservative agent that protects several surface fruits (external morphology) from microbial colonization (Dao and Dantigny, 2011). Hitherto, preservation of fresh fruits and vegetables is among the challenges of food products for commercial producers and distributors, particularly in middle-income, low or poor resource countries. Although many fresh fruits are in ideal conditions to hinder microorganisms from colonizing their integument, a lot of challenges in terms of post-harvest, storage, preservation of fruits or vegetables are rising up. It has generated the search of natural preservatives from plants or agro-wastes as an alternative and safer choice since they displayed little or no side effects (Sagar et al., 2018; Saeed et al., 2019). The bioethanol produced from agro-waste using S. carlsbergensis in submerged fermentation can be used as a preservative agent, which will not only help to reduce wastes in the environment but will preserve fruits or crops from post-harvest spoilage. Hence, this study aimed to produce bioethanol using BSG as a substrate with S. carlsbergensis in submerged fermentation and to assess the preservative potential of bioethanol on some selected fruits.
MATERIALS AND METHODS
Collection of brewer's spent grain and brewer's spent yeast
The brewer's spent grain and brewer's spent yeast were obtained from International Breweries Plc., Ilesha in Osun state, Nigeria. The town is located at longitude 7.6395°N and latitude 4.7588°E.
Collection of fruits from various locations in Osun State
Various types of fruits were collected from different locations in Osun State. This State was grouped into four (4) zones, A: Odeomu/Gbongan axis, B: Ife and its environment, C: Osogbo, and D: Ilesha and its environment. A total of 161 different fruits were collected as pineapple (Ananas comosus), orange (Citrus sinensis), African star apple (Chrysophyllum albidum), tomato (Solanum lycopersicum), banana (Musa acuminata), lime (Citrus aurantiifolia), pawpaw (Carica papaya), sour-sop (Annona muricata), watermelon (Citrullus lanatus), apple (Malus domestica), plantain (Musa paradisiaca) and almond (Prunus dulcis).
Source of S. carlsbergensis
S. carlsbergensis was isolated from brewer's spent yeast using serial dilution method. A loop full from 10-6 was cultured on Potato Dextrose Agar (PDA) and incubated at 25 °C for 48 h. The yeast was subcultured into another freshly prepared PDA and incubated at 25 °C for 48 h to get pure isolate of S. carlsbergensis. The pure isolate was transferred to yeast broth and incubated for 48 h.
Production of bioethanol from BSG
The method, according to Alam et al. (2009), was adopted for the production of bioethanol with a slight modification. The fermenter was sterilized using 3% v/v of hypochlorite. The sterile fermenter was loaded with 50 g L-1 of BSG at 29±2 °C and an agitation speed of 180 rpm. The pH was adjusted to 6.0 before the addition of 500 mL of yeast culture; the fermentation lasted 7 days under submerged and optimized conditions. The aeration and pH were kept stable during fermentation. The fermented product was concentrated using a rotatory evaporator at 66±1 °C, and ethanol was qualitatively determined by the dichromate method.
FTIR spectroscopic of bioethanol from BSG
Structural analysis of the functional group in bioethanol was determined using FT-IR spectroscopy (8400S, Shimadzu Scientific Instruments Inc.). BriefIy, bioethanol (1.0 µL) was placed on a fused KBr disc. This was placed on the cell holder, clamped loosely and fixed on the infrared (IR) beam. The running was done at 400 to 4000 per cm wavenumber.
Isolation and identification of microorganisms from fruits
The surface of fruit was sterilized with 1% v/v hypochlorite and rinsed with sterile distilled water to remove normal fIora of fruit and other possible microbial contaminants. Fruit samples were observed for 4-5 days to check any sign of spoilage. The rotten part of fruit was aseptically cut and transferred into sterilized peptone water. The sample was shaken vigorously and then allowed to stand for 30 min (Ajayi-Moses et al., 2019). Serial dilution was carried out up to10-4 and 10-5 dilution factor. An aliquot of 0.1 mL was aseptically transferred into Petri dish, and molten nutrient agar or PDA was then introduced. The plate solidified at room temperature (29±1 °C), then plates were incubated at 37 °C for 24 h and 2-3 days at 25 °C for bacteria and fungi, respectively. Discrete colonies were counted and recorded as colony-forming unit per gram (CFU g-1) for bacteria and spore-forming unit per gram (SFU g-1) for fungi. A pure colony was obtained by subcultured, and isolates were tentatively grouped according to their morphological, cultural, and staining characteristics. Biochemical tests such as the catalase test, production of hydrogen sulfide (H2S), indole, urease, methyl red, oxidase, coagulase, motility, methyl red, Voges-Proskauer, starch hydrolysis and sugars fermentation were carried out using the methods described by Olutiola et al. (2000). The results of the biochemical test were compared to Bergey's Manual of Systematic Bacteriology (Krieg et al., 2010). Fungi isolates were identified using cultural and microscopic observations, according to Barnett et al. (2000) and Samson et al. (2010).
In vitro antimicrobial activity of bioethanol and other preservatives against microorganisms
The antimicrobial activity of the bioethanol against spoilage microorganisms isolated from fruits was performed using agar well diffusion (CLSI, 2014). Suspension of test microorganisms was adjusted by the spectrophotometer to 0.5 McFarland standard. Sterile cotton swabs were dipped in the microbial suspension and spread on the surface of the agar plate. A sterilized cork borer was used for cutting wells in each plate. Bioethanol (50 μL) was introduced and sorbic acid (5.0 mg mL-1), ampiclox (5.0 mg mL-1) and terbinafine (5.0 mg mL-1) were implemented as positive controls against bacteria and fungi, while sterile distilled water was used as the negative control. All the plates were labeled appropriately and incubated at 37 °C during 24 h for bacteria and at 25 °C for 48 h for fungi. The diameter of zones of inhibition around wells was measured in millimeter (mm).
Preservation of fruits using bioethanol from BSG
Most prevalent as well as colonizing microorganisms (1.0×105 CFU mL-1 or SFU mL-1) were re-introduced to apparently healthy (absence of diseases, no wound symptoms or lesion) selected fruits (orange, watermelon, pineapple, and tomato). Each fruit was discretely remained in a sterile laminar hood without touching each other. After 24 h, bioethanol (20 mL) was applied on the surface fruits of the group A. The bioethanol was adjusted to 40% v/v since Kalathenos and Russell (2003) revealed that <30% v/v were rarely biocidal. Ethanol ≥70% v/v could cause damage to the fruit integument. Group B was treated with sorbic acid (5% w/v), group C was un-inoculated fruits with microorganism and group D was fruits inoculated with microorganisms but untreated with either bioethanol or sorbic acid. The fruit samples were observed for 7 days at room temperature (29 °C).
Statistical analysis
Data were presented as mean±standard deviation from three repetitions. Data obtained in this study were subjected to One-way Analysis of Variance (ANOVA) using Statistical Package for Social Sciences (SPSS) version 20 (USA). For bacteria and fungi count, the mean values were compared by Duncan's new multiple range test (MRT). Differences were considered significant at P<0.05.
RESULTS AND DISCUSSION
Yield and physicochemical parameters of bioethanol from BSG with S. cerevisiae
Table 1 shows the physicochemical parameters of bioethanol produced from BSG. The yield obtained was 22%, with a specific gravity of 0.8. These results contrast with the findings of Irfan et al. (2014). They reported bioethanol from sugarcane bagasse, rice straw and wheat straw by S. cerevisiae with a value of 77 g L-1, 62 g L-1, and 44 g L-1, respectively. Ingale et al. (2014) obtained ethanol of 17.1 g L-1 with a higher yield of 84% from banana pseudostem fermented by S. cerevisiae NCIM 3570. The higher proportion of ethanol in their study could be associated with two fungal strains of Aspergillus spp., which facilitated the maximal release of sugars to produce more ethanol. The combination of Aspergillus oryzae and S. cerevisiae NCYC479 produced the highest concentrations of ethanol (37 g L-1) in 10 days from BSG using consolidated bioprocessing (Wilkinson et al., 2017). Moodley and Gueguim Kana (2019) optimized pretreatment techniques to produce 25% more bioethanol from sugarcane leaf waste using S. cerevisiae BY4743. S. cerevisiae is one of the best yeast widely employed for commercial production of bioethanol. S. cerevisiae has been attractive for efficient consolidated bioprocessing and several biotechnological purposes because of its novel amylolytic enzyme combination, relatively high tolerance to osmotic stress and anaerobic conditions. Therefore, it is suitable for large-scale fermentation of agro-wastes into bioethanol (Cripwell et al., 2019). In the study of Wu et al. (2020), replacement of ethanol fermentation-associated regulatory gene in S. cerevisiae was reported to enhance ethanol production by a 5.30% increase in yield and 12.5% decrease in fermentation time when compared to the original strain. Another method that could increase the yield of ethanol is mixed substrates. Bolade et al. (2019) claimed that multi-substrates biomass of agro wastes increases the yield of bioethanol.
Table 1: Physicochemical properties of bioethanol produced from BSG.

BSG is interesting biomass with hydrolyzable fermentable sugars that can be converted to ethanol with different microorganisms through co-fermentation strategies (Rojas-Chamorro et al., 2020). Likewise, cassava peels, potato peels and millet husks with different microbial inoculants such as S. cerevisiae, Rhizopus nigricans, Aspergillus niger, Spirogyra africana showed great potential for bioethanol production (Chibuzor et al., 2016). The large proportion of lignocellulosic materials such as corncob, cornstalk, cornhusk, sugarcane bagasse and sugarcane bark that are creating environmental pollution, can be easily degraded by microorganisms and thus, serve as a substrate for renewable resources (bioethanol).
Figure 1 shows the transmittance and peak representing functional groups in the bioethanol. Table 2 shows various functional groups found in bioethanol, hydroxyl (OH stretch), methyl (CH3 stretch), and alkane (CH2 stretch). These functional groups in ethanol (alcohol) give it the biocide property, and it is responsible for the antisepsis, disinfection and can be used as a preservative agent (McDonnell and Russell, 1999). Hydroxyl group in ethanol (alcohol) acts as an antimicrobial agent against microbes except for alcohol-tolerant strains. It causes a partial breakdown of membrane function, inhibiting cell growth or protein synthesis, denaturation of proteins, and membrane damage, which lead to cell perturbations like ion leakage or loss of energy (Horinouchi et al., 2018).
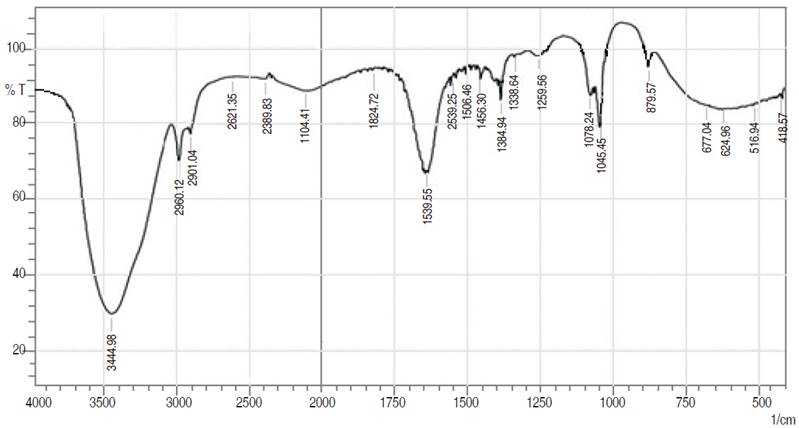
Figure 1: FTIR spectrum of bioethanol produced from BSG
Table 2: The functional group identified in bioethanol produced from BSG.
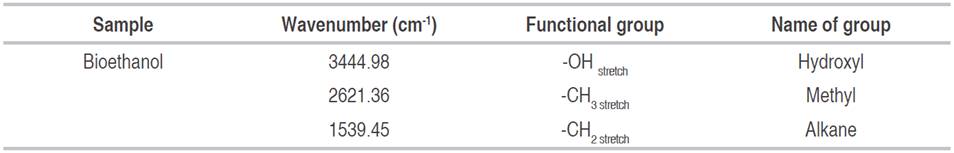
Total microbial load and occurrence (%) of microorganisms isolated from selected fruits
The colonies counted from each fruit are shown in Table 3.
Table 3: Bacteria and fungi count from fruits obtained from different regions in Osun State.
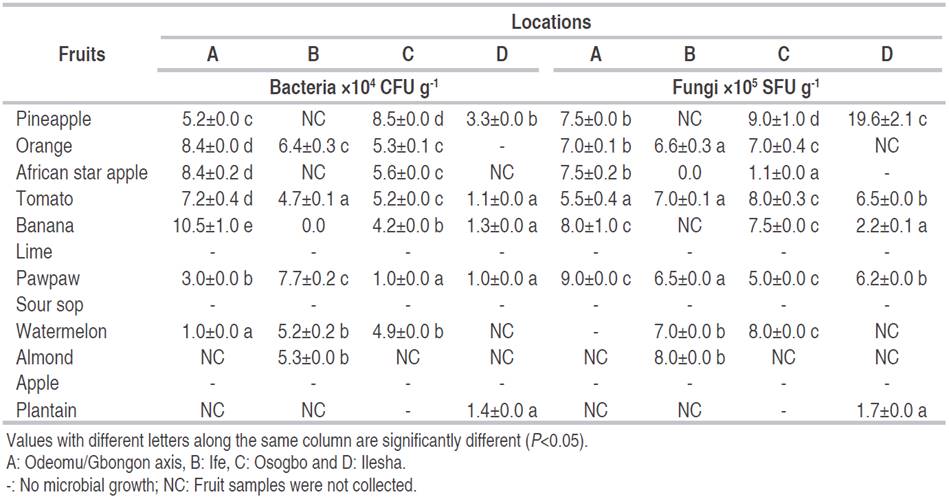
The highest colonies of bacteria count (1.05×105 CFU g-1) were recorded for bananas from Odeomu (A) market (P<0.05). Pineapple from Ilesha had the highest fungal count (1.96×106 SFU g-1). Watermelon from Odeomu (A) had the least bacterial load (P<0.05) of 1.0×104 CFU g-1, and African star apple collected from Osogbo (C) had the least fungal count of 1.1×105 SFU g-1. Ajayi-Moses et al. (2019) reported the highest bacterial count of 5.84×105 CFU g-1 for tomatoes and African star apple with the highest fungal count of 3.04×105 SFU g-1. Evaluation of microorganisms associated with fruits revealed that various bacteria and fungi in high densities could spoil fruits. Spoilage microorganisms can also colonize, enter and penetrate plant tissues at fruit development, either through calyx, stem or various specialized water and gas exchange structures of leafy matter. However, successful establishment requires the spoilage microbe to overcome multiple natural protective barriers and other factors (Erkmen and Bozoglu, 2016). Lime and soursop did not show any sign of spoilage, and no microorganisms were isolated (Table 3). Fruits like lime and soursop with higher pKa do not get spoilt but rather get dehydrated, exiting fruit juice, a process attributed to spoilage organisms affecting fruits (Czajkowski et al., 2011).
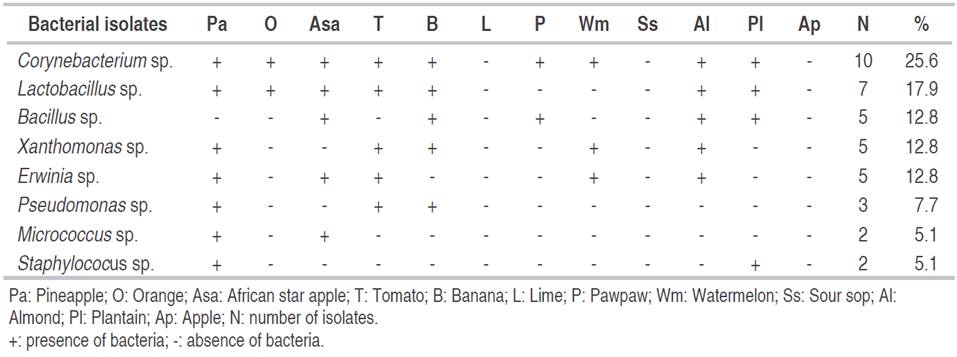
Table 4 shows the percentage of the occurrence of bacteria. The highest percentage of occurrence (25.6%) was obtained for Corynebacterium sp. followed by Lactobacillus sp. with 17.9%. The lowest bacteria percentage occurrence (5.1%) was obtained for species of Micrococcus and Staphylococus. Erwinia spp. were found to be associated with spoilage of pineapple, African star apple, tomato, watermelon and almond. The bacterium Erwinia carotovora subsp. carotovora is a highly effective spoilage microorganism that produces an increasing amount of pectolytic enzymes to degrade fruit tissues. It causes soft rot on fruits like oranges, tomatoes, banana, pineapple, and watermelon (Barth et al., 2009; Sharma et al., 2013). Besides, E. carotovora, several Pseudomonas spp., Corynebacterium, Xanthomonas campestris and lactic acid bacteria are important spoilage bacteria of fruits (Tournas, 2005; Erkmen and Bozoglu, 2016). Some spoilage microbes are capable of colonizing, creating lesions and damaged healthy plant tissues. The type of microbial spoilage in fruits is based on the pH, nutrient availability, water activity (aw), temperature, relative humidity, oxidation-reduction potential and content of biological structure of fruits (Erkmen and Bozoglu, 2016).
Table 4: Percentage of occurrence of bacteria isolated from different locations
Fungi with the highest percentage of occurrence was Aspergillus niger (13.9%), followed by Colletotrichum sp. and Penicillium digitatium with the same value of 11.1%. Pichia sp. has the lowest percentage of occurrence of 4.2% (Table 5). The highest occurrence of A. niger from examined fruits in Osun State is in concordances with findings of Mailafia et al. (2017). Researchers revealed that Aspergillus spp. had the highest occurrence in fruits like pineapple, watermelon, oranges, pawpaw, and tomatoes with a frequency of 38%, followed by Fusarium avenaceum with occurrence of 31% in pineapple, watermelon, oranges, pawpaw and tomatoes. In comparison, P. digitatum and R. stolonifera have the least frequency at the same value of 4% for tomato and orange. Other fungal species identified as agents of spoilage were Saccharomyces spp. (10%), F. solani (8%), and A. flavus (5%). Some of the fungi isolated from fruits were species of Penicillum, Aspergillus, Rhizopus, Fusarium and Mucor iriformis. Tafinta et al. (2013) and Ajayi-Moses et al. (2019) isolated similar microorganisms with varying prevalence from banana, pawpaw, orange, tomato, apple, pineapple, watermelon, cucumber, and African star apple. Some of these fungal isolates are known to be pathogenic due to the toxic secondary metabolites produced. Penicillium expansum and Botrytis cinerea are pathogenic spoilage microorganisms, which cause blue-rot, grey mold in African star apple, cherry, apple, tomato, pears and kiwi fruit called sour sop (Miedes and Lorences, 2004).
Table 5: Percentage of occurrence of fungi isolated from different locations

Likewise, molds of genera Rhizopus, Alternaria and Botrytis produce acidic compounds that cause fruit and vegetable rot with distorted color, texture and or taste (Tournas, 2005). Fernández-Cruz et al. (2010), Lewis and Goodrich-Schneider (2012) revealed different species of fungi that produce mycotoxins (aflatoxins, ochratoxin, patulin, fumonisin, alternariol, alternariol methyl ether, and altenuene) in fruits. The problem associated with mycotoxins in fruits includes economic loss, poor organoleptic properties, toxicities (acute to chronic), and a spectrum of effect (mild to severe), including carcinogenicity and death. Spoilage microorganisms exploit fruit components using their extracellular lytic enzymes, pectinases and hemicellulases to degrade fruit polymers to release intracellular constituents as nutrients for growth (Kalia and Gupta, 2006). Microorganisms from fruits secreted a wide variety of enzymes. Isolation and identification of novel strains of microorganism from fruits can serve as natural origin, a safer and cheaper alternative source of microbial enzymes as promising candidates for biotechnological uses and medical processes (Sharma et al., 2013; Garg et al., 2016).
Antimicrobial and preservative properties of bioethanol from BSG
To minimize wastage recorded on fruits and vegetables and to reduce economic losses associated with microbial spoilage, a reliable and supportive measure needs to be sourced. Bioethanol from BSG displayed zones of inhibition against bacteria with values ranging from 7.0 to 8.8 mm, sorbic acid was within 6.0 to 13.0 mm, and ampiclox was from 8.0 to 18.5 mm (Table 6). Zones of inhibition by bioethanol against fungi ranged from 7.5 to 11.5 mm, while sorbic acid was 7.5 to 13.0 mm and 10.0 to 17.0 mm for terbinafine (Table 6). Ethanol is used as a disinfectant and acts against microorganisms in two different ways: growth inhibition (bacteriostasis, fungistasis) or lethal action (bactericidal, fungicidal or virucidal effects) (Maris, 1995). Hence, bioethanol can be used to prevent the growth of spoilage microorganisms on fruits. Ethanol interacts with cell surface followed by penetration into cells and acts on the target site(s) of microorganisms (Maris, 1995). Inactivation of fungal spores, suppression of fungal growth or their germination on fruits by ethanol reflects the inhibitory potential of bioethanol in controlling fruit decaying by fungi and extending the shelf-life of food products (Dao and Dantigny, 2011).
Table 6: Zones of inhibition (mm) displayed by bioethanol and selective preservatives against #microorganisms isolated from fruits.
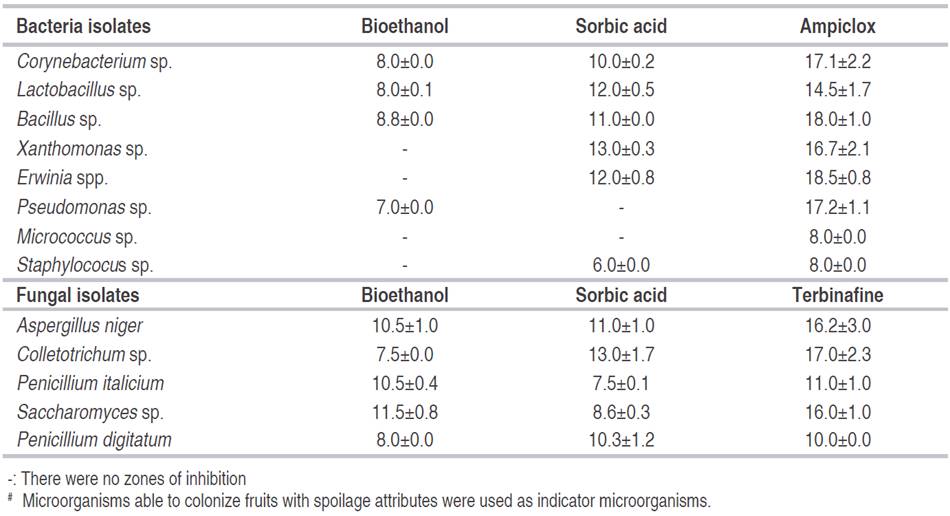
Table 7 shows different fruits preserved with bioethanol, sorbic acid, un-inoculated fruit with the microorganism, spoilt fruit and open spoilt fruit after 7 days. Dao and Dantigny (2011), found that oranges were protected from fungal infection for 30 days when exposed to 20-100% ethanol. The use of bioethanol as a preservative is important to prevent a higher loss of fruits since it has no side effects on humans. It will proffer the solution to longstanding spoilage of vegetables and fruits, which had been associated with different microorganisms. Ethanol exerts its most effective bactericidal action at ≥40 to 95% v/v against vegetative cells like chemical disinfectants (Kalathenos and Russell, 2003). Findings of Katsinis et al. (2008) established effective preservation with the synergistic effect of conventional chemical preservative (potassium sorbate) and ethanol, which improved the shelf life of bread by suppressing microbial growth (43.5% and 38.5% mold-free shelf life). Bioethanol displayed a higher zone of inhibition (11.5 mm) against Saccharomyces sp. than sorbic acid (8.6 mm). Sorbic acids had less or no inhibitory activity against S. cerevisiae, Saccharomycodes ludwigii, Zygosaccharomyces bailii, Zygosaccharomyces rouxii, Schizosaccharomyces pombe Brettanomyces spp., Pichia membranifaciens, Dekkerae spp, and Issatchenkia orientalis at 800 mg kg-1, which is much higher than the permissible limit in foods, but ethanol (>20% v/v) exerts a strong lethal effect on these molds (Loureiro and Malfeito-Ferreira, 2003).
Table 7: Evolution of fruits during 7-day treatment with bioethanol and sorbic acid

The mechanisms of resistance by yeasts to weak organic acids are by inducing the expression of H+ ATPases to regulate their cytosolic pH, using their plasma membrane components to modulate the influx of lipophilic weak organic acids (Ullah et al., 2012). ATP binding cassette (ABC) transporter (Pdr12) prevents anion accumulation or degrade sorbic acid to 1,3-pentadiene (Casas et al., 2004). Findings of Linares-Morales et al. (2018) revealed that chemical preservatives like organic acids in fruits and vegetables cause disruption of membrane permeability, reduce cell's internal pH, affects metabolic enzymes as well as protein synthesis. However, a microbial product like bioethanol can be encouraged in use as preservative agent since it strongly suppresses microbial activity on fruits. Microbial by-products are bio-protective or natural preservatives, and thus, exhibit antifungal activities. There is a growing interest in alternatives to preservation other than chemical or synthetic agents with different side effects (Leyva Salas et al., 2017). The microbial growth inhibition or killing action of bioethanol on microorganisms is an indication that; bioethanol produced by S. cerevisiae from renewable biomass (lignocellulosic wastes) is needed for commercial purposes.
CONCLUSION
The bioethanol produced from the fermentation of brewer's spent grain with S. carlsbergensis inhibited microorganisms associated with post-harvest spoilage of fruits. The presence of functional groups in bioethanol contributed to its bioactivity, which makes it a potential preservative agent to suppress the colonization of microbial spoilage on fruits, reaching inhibitions of 8.8 and 11.5 mm for bacteria and fungi, respectively. This study proffer solution to the underutilized BSG residue, which can be used for bioethanol and serve as a preservative agent for fruits or vegetables. However, modern facilities to increase the yield of bioethanol from agro wastes need to be considered in subsequent works.
REFERENCES
References
Ajayi-Moses OB, Ogidi CO and Akinyele BJ. 2019. Bioactivity of Citrus essential oils (CEOs) against microorganisms associated with spoilage of some fruits. Chemical and Biological in Technologies Agricultures 6(22): 15. doi: 10.1186/s40538-019-0160-5
Alam MZ, Kabbashi NA and Hussi SNIS. 2009. Production of bioethanol by direct bioconversion of oil-palm industrial effluent in a stirred-tank bioreactor. Journal of Industrial Microbiology and Biotechnology 36(6): 801-808. doi: 10.1007/s10295-009-0554-7
Aliyu S and Bala M. 2011. Brewer’s spent grain: a review of its potentials and applications. African Journal of Biotechnology 10(3): 324-331. doi: 10.5897/AJB10.006
Azhar SHM, Abdulla R, Jambo SA, Marbawi H, Gansau JA,Faik AAM and Rodrigues KF.
Yeasts in sustainable bioethanolproduction: a review. Biochemistry and Biophysics Reports 10: 52-61.doi: 10.1016/j.bbrep.2017.03.003
Awoyale AA and Lokhat D. 2019. Harnessing the potential of bio‐ethanol production from lignocellulosic biomass in Nigeria – a review. Biofuels, Bioproducts and Biorefining 13(1): 192-207. doi: 10.1002/bbb.1943
Barnett JA, Payne RW and Yarrow O. 2000. Yeast: Characteristics and identification. Third Edition. Cambridge University Press, London. pp 12-87.
Barth M, Hankinson TR, Zhuang H and Breidt F. 2009. Microbiological spoilage of fruits and vegetables pp. 135–183. In: Sperber WH and Doyle MP (eds.). Compendium of the microbiological spoilage of foods and beverages: Food Microbiology and Food Safety. Springer, New York. doi: 10.1007/978-1-4419-0826-1_6
Bolade DO, Ana GREE, Lateef SA and Sokan-Adeaga AA. 2019. Exploration of the Bioethanol Yield of Single and Multi-Substrate Biomass from Cassava Processing Wastes. The Journal of Solid Waste Technology and Management 45(3): 305-314. doi: 10.5276/JSWTM/2019.305
Bušić A, Marđetko N, Kundas S, Morzak G, Belskaya H, Ivančić Šantek M, Komes D, Novak S and Šantek B. 2018. Bioethanol production from renewable raw materials and its separation and purification: a review. Food Technology and Biotechnology 56(3): 289–311. doi: 10.17113/ftb.56.03.18.5546
Casas E, de Ancos B, Valderrama MJ, Cano P and Peinado JM. 2004. Pentadiene production from potassium sorbate by osmotolerant yeasts. International Journal of Food Microbiology 94(1): 93–96. doi: 10.1016/j.ijfoodmicro.2004.01.001
Chibuzor O, Uyoh EA and Igile G. 2016. Bioethanol production from cassava peels using different microbial inoculants. African Journal of Biotechnology 15(30): 1608-1612. doi: 10.5897/AJB2016.15391
CLSI - Clinical and Laboratory Standards Institute. 2014. Performance standards for antimicrobial susceptibility testing; twenty-fourth informational supplement. M100 – S24. Wayne, USA. pp. 1-16.
Cripwell RA, Rose SH, Favaro L and van Zyl W. 2019. Construction of industrial Saccharomyces cerevisiae strains for the efficient consolidated bioprocessing of raw starch. Biotechnology for Biofuels 12:201. doi: 10.1186/s13068-019-1541-5
Czajkowski MC, Perombelon JA, Veen JM and Van der W. 2011. Control of blackleg and tuber spoilage of potatoes caused by Pectobacterium and Dickeya species: a review. Plant Pathology 60(6): 999-1013. doi: 10.1111/j.1365-3059.2011.02470.x
Dao T and Dantigny P. 2011. Control of food spoilage fungi by ethanol. Food Control 22(3-4): 360-368. doi: 10.1016/j.foodcont.2010.09.019
Erkmen O and Bozoglu TF. 2016. Chapter 20: Spoilage of vegetables and fruits. pp 337-363. In: Food microbiology: principles into practice. doi: 10.1002/9781119237860.ch20
Fernández-Cruz ML, Mansilla ML and Tadeo JL. 2010. Mycotoxins in fruits and their processed products: analysis, occurrence and health implications. Journal of Advanced Research 1(2): 113-122. doi: 10.1016/j.jare.2010.03.002
Garg G, Singh A, Kaur A, Singh R, Kaur J and Mahajan R. 2016. Microbial pectinases: an ecofriendly tool of nature for industries. 3 Biotech 6: 47. doi: 10.1007/s13205-016-0371-4
Horinouchi T, Maeda T and Furusawa C. 2018. Understanding and engineering alcohol-tolerant bacteria using OMICS technology. World Journal of Microbiology and Biotechnology 34: 157. doi: 10.1007/s11274-018-2542-4
Ingale S, Joshi SJ and Gupte A. 2014. Production of bioethanol using agricultural waste: banana pseudo stem. Brazilian Journal of Microbiology 45(3): 885-892. doi: 10.1590/S1517-83822014000300018
Irfan M, Nadeem M and Syed Q. 2014. Ethanol production from agricultural wastes using Saccharomyces cerevisiae. Brazilian Journal of Microbiology 45(2): 457–465. doi: 10.1590/s1517-83822014000200012
Ivanova K, Denkova R, Kostov G, Petrova T, Bakalov I, Ruscova M and Penov N. 2017. Extrusion of brewers’ spent grains and application in the production of functional food. Characteristics of spent grains and optimization of extrusion. Journal of The Institute of Brewing 123(4): 544-552. doi: 10.1002/jib.448
Kalathenos P and Russell NJ. 2003. Ethanol as a food preservative. pp 196-217. In: Russell NJ and Gould GW. (eds.) Food preservatives. Springer, Boston. doi: 10.1007/978-0-387-30042-9_10
Kalia A and Gupta R. 2006. Fruit Microbiology. pp. 1-28. In: Hui YH (ed.). Handbook of fruit and fruit processing. First Edition. Blackwell publishing, United Kingdom. doi: 10.1002/9780470277737.ch1
Katsinis G, Rigas F and Doulia D. 2008. Synergistic effect of chemical preservatives with ethanol on the microbial shelf life of bread by factorial design. International Journal of Food Science and Technology 43(2): 208-215 doi: 10.1111/j.1365-2621.2006.01386.x
Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, Ludwig W and Whitman WB. 2010. Bergey’s Manual of Systematic Bacteriology. Second edition. Springer, New York. pp. 40-91. doi: 10.1007/978-0-387-68572-4
Lewis DC and Goodrich-Schneider R. 2012. Mycotoxins in fruit and fruit products. Proceedings of the Florida State Horticultural Society. 125:252–257.
Leyva Salas M, Mounier J, Valence F, Coton M, Thierry A and Coton E. 2017. Antifungal microbial agents for food biopreservation- A review. Microorganisms 5(3): 37. doi: 10.3390/microorganisms5030037
Linares-Morales JR, Gutiérrez-Méndez N, Rivera-Chavira BE, Pérez-Vega SB and Nevárez-Moorillón GV. 2018. Biocontrol processes in fruits and fresh produce, the use of lactic acid bacteria as a sustainable option. Frontiers in Sustainable Food System 2:50. doi: 10.3389/fsufs.2018.00050
Loureiro V and Malfeito-Ferreira M. 2003. Spoilage yeasts in the wine industry. International Journal of Food Microbiology 86(1-2): 23-503. doi: 10.1016/S0168-1605(03)00246-0
Mailafia S, Okoh GR, Olabode H and Osanupin R. 2017. Isolation and identification of fungi associated with spoilt fruits vended in Gwagwalada market, Abuja, Nigeria. Veterinary World 10(4): 393–397. doi: 10.14202/vetworld.2017.393-397
Maris P. 1995. Modes of action of disinfectants. Revue scientifique et technique (International Office of Epizootics) 14 (1):47-55. https://doc.oie.int/dyn/portal/index.seam?page=alo&aloId=27748
McDonnell G and Russell AD. 1999. Antiseptics and disinfectants: activity, action, and resistance. Clinical Microbiology Reviews 12(1): 147-179. doi: 10.1128/CMR.12.1.147
Miedes E. and Lorences EP. 2004. Apple (Malus domestica) and tomato (Solanum lycopersicum) fruits cell-wall hemicelluloses and xyloglucan degradation during Penicillium expansum infection. Journal of Agricultural and Food Chemistry. 52(26): 7957–7963. doi: 10.1021/jf048890f
Azhar SHM, Abdulla R, Jambo SA, Marbawi H, Gansau JA, Faik AAM and Rodrigues KF. 2017. Yeasts in sustainable bioethanol production: a review. Biochemistry and Biophysics Reports 10: 52–61. doi: 10.1016/j.bbrep.2017.03.003
Moodley P and Gueguim Kana EB. 2019. Bioethanol production from sugarcane leaf waste: Effect of various optimized pretreatments and fermentation conditions on process kinetics Biotechnology Reports 22: e00329. doi: 10.1016/j.btre.2019.e00329
Mussatto SI. 2014. Brewer’s spent grain: a valuable feedstock for industrial applications. Journal of the Science of Food and Agriculture 94(7): 1264-1275. doi: 10.1002/jsfa.6486
Olutiola PO, Famurewa O and Sonntag HG. 2000. An introduction to General Microbiology, a practical approach. Heldelberg, Nigeria. pp. 1-267.
Prasad RK, Chatterjee S, Mazumder PB, Gupta SK, Sharma S, Vairale MG, Datta S, Dwivedi SK and Gupta DK. 2019. Bioethanol production from waste lignocelluloses: A review on microbial degradation potential. Chemosphere 231: 588-606. doi: 10.1016/j.chemosphere.2019.05.142
Rojas-Chamorro JA, Romero-García JM Cara C, Romero I and Castro. E. 2020. Improved ethanol production from the slurry of pretreated brewers’ spent grain through different co-fermentation strategies. Bioresource Technology 296: 122367. doi: 10.1016/j.biortech.2019.122367
Saeed F, Afzaal M, Tufail T and Ahmad A. 2019. Use of natural antimicrobial agents: a safe preservation approach. In: Var I and Uzunhu S (eds.). Active antimicrobial food packaging. IntechOpen, London. doi: 10.5772/intechopen.80869
Sagar NA, Pareek S, Sharma S, Yahia EM and Lobo MG. 2018. Fruit and vegetable waste: bioactive compounds, their extraction, and possible utilization. Comprehensive Reviews in Food Science and Food Safety 17(3): 512-531. doi: 10.1111/1541-4337.12330
Saini JK, Saini R and Tewari L. 2015. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: concepts and recent developments. 3 Biotech 5: 337–353 doi: 10.1007/s13205-014-0246-5
Samson RA, Houbraken J, Thrane U, Frisvad JC and Andersen B. 2010. Fungi and indoor fungi. CBS-KNAW Fungal Biodiversity Centre, Utrecht. pp.1-390.
Sharma N, Rathore M and Sharma M. 2013. Microbial pectinase: sources, characterization and applications. Review in Environmental Science and Biotechnology 12: 45–60. doi: 10.1007/s11157-012-9276-9
Singh R, Kapoor V and Kumar V. 2012. Utilization of agro-industrial wastes for the simultaneous production of amylase and xylanase by thermophilic Actinomycetes. Brazilian Journal of Microbiology 43(4): 1545–1552.
Steiner J, Procopio S and Becker T. 2015. Brewer’s spent grain: source of value-added polysaccharides for the food industry in reference to the health claims. European Food Research and Technology 241(3): 303-315. doi: 10.1007/s00217-015-2461-7
Tafinta IY, Shehu K, Abdulganiyyu H, Rabe AM and Usman A. 2013. Isolation and identification of fungi associated with the spoilage of sweet orange (Citrus sinensis) fruit in Sokoto State. Nigerian Journal of Basic and Applied Sciences 21(3): 193-196. doi: 10.4314/njbas.v21i3.4
Tan YX, Mok WK and Chen WN. 2020. Potential novel nutritional beverage using submerged fermentation with Bacillus subtilis WX-17 on brewers’ spent grains, Heliyon 6(6): e04155. doi: 10.1016/j.heliyon.2020.e04155
Tournas VH. 2005. Spoilage of vegetable crops by bacteria and fungi and related health hazards. Critical Reviews in Microbiology 31(1): 33-44. doi: 10.1080/10408410590886024
Ullah A, Orij R, Brul S and Smits GJ. 2012. Quantitative analysis of the modes of growth inhibition by weak organic acids in Saccharomyces cerevisiae. Applied Environmental Microbiology 78: 8377–8387. doi: 10.1128/AEM.02126-12
Wilkinson S, Smart KA, James S and Cook DJ. 2017. Bioethanol production from brewers spent grains using a fungal consolidated bioprocessing (CBP) approach. Bioenergy Research 10(1): 146-157. doi: 10.1007/s12155-016-9782-7
Wu M, Li H, Wei S, Wu H, Wu X, Bao X, Hou J, Liu W and Shen Y. 2020. Simulating Extracellular Glucose Signals Enhances Xylose Metabolism in Recombinant Saccharomyces cerevisiae. Microorganisms 8(1): 100. doi: 10.3390/microorganisms8010100
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
CrossRef Cited-by
1. Marcos Barrera-León, Elí Terán-Cabanillas, Roberto de Jesús Avena-Bustillos, Feliznando Isidro Cárdenas-Torres, Bianca Anabel Amézquita-López, Mario Armando Gómez-Favela, David Moroni Alemán-Hidalgo, Mayra Arias-Gastélum. (2025). Transformation of Brewer’s Spent Grain Through Solid-State Fermentation: Implications for Nutrition and Health. Recycling, 10(5), p.170. https://doi.org/10.3390/recycling10050170.
2. Christopher Osazuwa, Oladipo Oladiti Olaniyi , Bamidele Juliet Akinyele, Felix Akinsola Akinyosoye . (2024). Poultry meat preservation with citric acid obtained from the fermentation of wheat straw by Aspergillus niger. Revista Facultad Nacional de Agronomía Medellín, 77(2), p.10729. https://doi.org/10.15446/rfnam.v77n2.105711.
3. Prachi Arora, Nidhi Godiyal, Anurag Mishra, Sakshi Tewari. (2026). Biofuels and Bioproducts from Lignocellulosic Materials. , p.139. https://doi.org/10.1016/B978-0-443-21527-8.00003-4.
4. Adeolu A. Awoyale, David Lokhat. (2021). Hybridization of Selected Nigerian Lignocellulosic Biomass Feedstocks for Bioethanol Production: Modeling and Optimization of Pretreatment and Fermentation Process Parameters Using Response Surface Methodology. Journal of Oleo Science, 70(7), p.1013. https://doi.org/10.5650/jos.ess21038.
5. Augreat Chiyangwa, Lee Moyo, Milton Manyangadze, Elizabeth Ticharwa, Lonestar Gonde. (2025). A concise review of wild loquate fruits/mazhanje beneficiation: Production of wine from uapaca kirkiana (wild loquate fruits/mazhanje). Cogent Food & Agriculture, 11(1) https://doi.org/10.1080/23311932.2025.2562174.
6. Vishal Ahuja, Shikha Chauhan, Sukhvinder Singh Purewal, Sanjeet Mehariya, Anil Kumar Patel, Gopalakrishnan Kumar, Mallavarapu Megharaj, Yung-Hun Yang, Shashi Kant Bhatia. (2024). Microbial alchemy: upcycling of brewery spent grains into high-value products through fermentation. Critical Reviews in Biotechnology, 44(7), p.1367. https://doi.org/10.1080/07388551.2023.2286430.
7. Zhengjie Xie, Meiling Dan, Guohua Zhao, Damao Wang. (2024). Recent advances in microbial high-value utilization of brewer’s spent grain. Bioresource Technology, 408, p.131197. https://doi.org/10.1016/j.biortech.2024.131197.
8. Gamze Nur Müjdeci, Kianoush Khosravi-Darani. (2022). Fruits and Vegetable Wastes. , p.315. https://doi.org/10.1007/978-981-16-9527-8_13.
9. Vanisree G S, P T Archana, Aparna K. (2025). Sustainable bioethanol synthesis from Queen sago seed shells via sequential acid hydrolysis and fermentation. Materials Research Express, 12(8), p.085502. https://doi.org/10.1088/2053-1591/adf695.
Dimensions
PlumX
Article abstract page views
Downloads
License
Copyright (c) 2020 Revista Facultad Nacional de Agronomía, Autores

This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.
The journal allows the author(s) to maintain the exploitation rights (copyright) of their articles without restrictions. The author(s) accept the distribution of their articles on the web and in paper support (25 copies per issue) under open access at local, regional, and international levels. The full paper will be included and disseminated through the Portal of Journals and Institutional Repository of the Universidad Nacional de Colombia, and in all the specialized databases that the journal considers pertinent for its indexation, to provide visibility and positioning to the article. All articles must comply with Colombian and international legislation, related to copyright.
Author Commitments
The author(s) undertake to assign the rights of printing and reprinting of the material published to the journal Revista Facultad Nacional de Agronomía Medellín. Any quotation of the articles published in the journal should be made given the respective credits to the journal and its content. In case content duplication of the journal or its partial or total publication in another language, there must be written permission of the Director.
Content Responsibility
The Faculty of Agricultural Sciences and the journal are not necessarily responsible or in solidarity with the concepts issued in the published articles, whose responsibility will be entirely the author or the authors.


















