



Publicado
Aspectos del polimorfismo genético en Drosophila pseudoobscura, D. willistoni y D. nebulosa
Aspects of genetic polymorphism in Drosophila pseudoobscura, D. willistoni and D. nebulosa
Palabras clave:
Drosophila, polimorfismo cromosómico, poblaciones naturales, México, chromosomal polymorphism, natural populations (es)Drosophila, chromosomal polymorphism, natural populations, Mexico. (en)
Aspectos del polimorfismo genético en Drosophila pseudoobscura, D. willistoni y D. nebulosa
Aspects of genetic polymorphism in Drosophila pseudoobscura, D. willistoni and D. nebulosa
Víctor Manuel Salceda Sacanelles
Departamento de Biología, Instituto Nacional de Investigaciones Nucleares. Carretera México-Toluca S/N, La Marquesa, Ocoyoacac, México, 52750. (Autor para correspondencia:vmss@nuclear.inin.mx)
REC: 28-06-07. ACEPT.: 18-04-08
RESUMEN
En México el polimorfismo cromosómico en poblaciones naturales de Drosophila se estudió en D. pseudoobscura, D. willistoni y D. nebulosa. En la primera, más ampliamente analizada, se reconocen 27 inversiones correspondientes al 67.5% de la variabilidad para esta característica, identificando cuatro razas. Diferentes recolectas detectaron nuevas inversiones que permitieron reestructurar el árbol filogenético para la característica. Con base en el alto número de estas nuevas inversiones y otras evidencias se postuló como arreglo ancestral el Tree Line (TL) y se sugiere como centro de origen de la especie la región en que colindan los estados de Michoacán y Zacatecas. Se encontraron gradientes geográficos para algunas inversiones tanto en localidades distantes como micro-geográficas. Un estudio de recolectas semestrales por ocho años con las otras dos especies señaló alto grado de diversidad para la frecuencia de inversiones, 64.3% para D. willistoni y 52.9% para D. nebulosa. En ambas especies se cuantificó el número promedio de inversiones por hembra de 2.57 para D. willistoni y de 1.57 para D. nebulosa, que las sitúan dentro del área marginal de la distribución de la especie. Sin embargo, conservan alto grado de diversidad que aparentemente no les correspondería debido a su marginalidad.
Palabras claves: Drosophila; polimorfismo cromosómico; poblaciones naturales; México.
ABSTRACT
In Mexico, chromosomal polymorphism in Drosophila has been studied in D. pseudoobscura, D. willistoni and D. nebulosa. The first species is better studied with respect to the number of populations analyzed (close to 60), in which there were recognized up to 27 inversions corresponding to the 67.5 % of the total variability for this characteristic, identifying four races. With all of these evidences we rebuilt phylogenetic tree and postulate as ancestral gene arrangement the sequence Tree Line (TL) and suggesting as origin center of the species the surrounded area between Michoacán and Zacatecas states. Geographical gradients for several inversions at both macro and micro-geographic scales were also detected. The other species were both studied for a seasonal changes during eight years. As a result, it was found a high variability degree for the inversions number, 64.3% of the total variability for this parameter in D. willistoni and 52.9 % in D. nebulosa. Average number of inversions carried by a female was 2.57 in D. willistoni and 1.57 in D. Nebulosa, which set as marginal with respect to the distribution. Nevertheless, they show a considerable variability not expected for a marginal population.
Key words: Drosophila; chromosomal polymorphism; natural populations; Mexico.
INTRODUCCIÓN
La existencia de segmentos invertidos en los cromosomas de Drosophila fue detectada por primera vez por Sturtevant (1917). Pruebas del orden invertido en los marcadores genéticos entre D. melanogaster y D. simulans fueron presentadas por Sturtevant y Plunkett (1926). Painter (1934) y Bridges (1935), trabajando con los cromosomas politécnicos de D. melanogaster, sentaron las bases para una serie de estudios en elevado número de especies. Tan (1935) y Koller (1936) en D. pseudoobscura y Sturtevant y Dobzhansky (1936) en poblaciones de esta especie y de D. persimilis a lo largo de la distribución geográfica.
Los estudios se ampliaron a nivel mundial en poblaciones naturales de especies de Drosophila. Dubinin y Tiniakov (1945) con D. funebris, Pavan (1946) con D.nebulosa, Carson y Stalker (1947) con D. robusta, Dobzhansky (1950) con D. willistoni, Goldschmidt (1956) con D. subobscura y Brncic (1962) con D. flavopilosa.
En los cromosomas politénicos de las glándulas salivales de las larvas de Drosophila es posible detectar el polimorfismo para inversiones de secciones cromosómicas, especialmente para inversiones paracéntricas. Las inversiones se generan por dos rupturas, independientes y simultáneas en el mismo cromosoma, acompañadas de un giro de 180°, con la posterior reunión de las regiones involucradas, la ocurrencia del evento es de origen único. La inversión se perpetúa por herencia y puede ser sujeta a eventos similares. Una representación del fenómeno es:
Secuencia original ABCDEFGH
Puntos de ruptura AB/CDEF/GH, seguido del giro de 180°
Secuencia final invertida ABFEDCGH
Las nuevas inversiones pueden ser independientes o incluidas en las previas, por ejemplo:
ABCDEFGHIJ ® ABDECFHGJI independiente
ABCDEFGH ® ACBEDFGH incluida
Existen dos tipos de inversiones, paracéntricas y pericéntricas. Estas últimas involucran el centrómero entre los dos brazos del cromosoma. Las primeras involucran únicamente un brazo cromosómico, y son las de mayor presencia e importancia al considerar los estudios acerca de los polimorfismos cromosómicos. Existen dos maneras de distinguir las inversiones, cuando el cromosoma o individuo es homocigoto se diferencia banda por banda y se compara con el patrón “Standard” o ST. En cromosomas o individuos heterocigotos se distinguen las configuraciones mediante evidencia fotográfica (Kastritsis y Crumpacker, 1966 y 1967; Olvera et al., 1979).
En muchas especies de Drosophila el polimorfismo cromosómico es de carácter adaptativo, lo que le confiere importancia ecológica y evolutiva. En este caso el alelo menos frecuente debe estar presente entre 1% o 5% como valores mínimos.
La investigación se propuso analizar el polimorfismo cromosómico en tres especies de Drosophila (D. pseudoobscura, D. willistoni y D. nebulosa), habitantes naturales y exclusivas del continente americano. El estudio hizo hincapié en la distribución en México.
MATERIALES Y MÉTODOS
Las recolectas se realizaron durante una semana por localidad. Las moscas se capturaron en recipientes plásticos (25 cm de diámetro por 15 cm de alto) con frutas fermentadas y se transportaron en grupos de 20- 25 hembras. En el laboratorio cada hembra se separó en frascos de un cuarto de litro con alimento fresco. Luego de ocho días se pasó a un nuevo frasco de cultivo y al cultivo original se agregaron gotas de solución concentrada de levadura de cerveza fresca para sobrealimentar las larvas y obtener glándulas salivales grandes.
Las larvas se colocaron en portaobjetos con solución al 2% de aceto-orceína. Las glándulas salivales se tiñeron por 3-5 minutos. Se analizaron con ayuda de un atlas fotográfico y figuras de algunos reportes, identificándose la(s) inversión(es) presente(s). Se tomaron notas hasta completar la muestra obtenida en la naturaleza. Se calcularon las frecuencias relativas de cada inversión y se construyeron las tablas.
RESULTADOS Y DISCUSIÓN
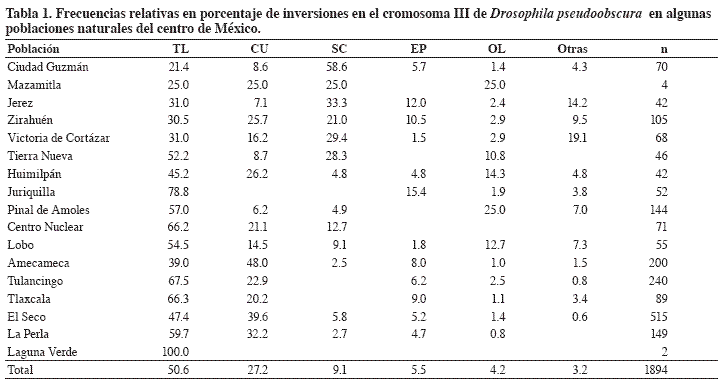
D. pseudoobscura es una especie del Neártico que habita bosques templados de pino-encino-abeto y que se distribuye desde la Columbia Británica en Canadá, el oeste de Estados Unidos, México, Guatemala y una colonia en las cercanías de Bogotá, Colombia (Dobzhansky, 1963). El tercer par de cromosomas es más polimórfico (Dobzhansky y Epling, 1944), ya que en el cromosoma X sólo se conoce la inversión denominada “sex-ratio” (proporción sexual). En el resto de los cromosomas no se tiene conocimiento de polimorfismo. Las inversiones constituyen eventos mutacionales sin relevancia. Para el cromosoma III se conocen 40 tipos estructurales y se pueden añadir 3-4 aún no descritas (Olvera et al., 1979) y relacionadas entre sí como inversiones traslapadas constituyendo una serie filogenética (Figura 1). Con referencia al polimorfismo cromosómico se han reportado en México frecuencias relativas para diferentes inversiones (Tabla 1) en aproximadamente 60 poblaciones (Dobzhansky,1939 y 1948; Dobzhansky et al., 1975; Levine et al.,1975; Espinoza-Velázquez y Salceda,1981; Olvera et al., 1982; Salceda, 2001; Olvera et al.,2005;Guzmán et al.,2005; Salceda et al.,2007a y b).
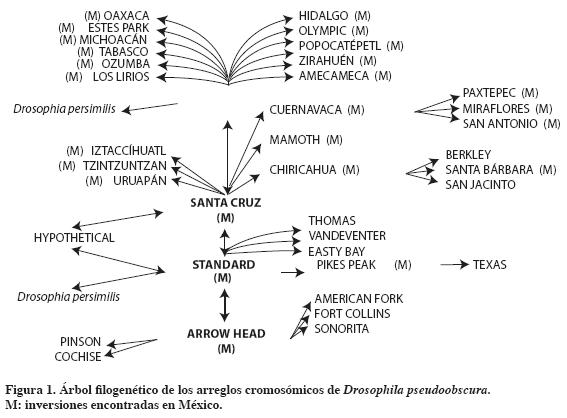
Todas las poblaciones muestran alto grado de variabilidad en arreglos cromosómicos presentes en cada población. Hoy se sabe que existen 27 inversiones, la mayoría endémicas. Esto representa el 67.5% del total de la variabilidad para este parámetro en la especie y permitió la elaboración del árbol filogenético actualizado.
El aporte de las investigaciones en poblaciones mexicanas puede notarse al comparar el árbol filogenético presentado por Dobzhansky y Epling (1944) con el elaborado por Olvera et al. (1979). En el trabajo de 1944 al separar la especie D. persimilis quedó D. pseudoobscura representada por una sola raza. Posteriormente Dobzhansky (1948) concluyó que constaba de siete razas que dependen de las frecuencias relativas de los diferentes componentes presentes en cada región. En México, pese a tener menor área respecto a Estados Unidos y en relación con la distribución de esta especie, coexisten cuatro razas.
La amplia distribución de la inversión Tree Line (TL), así como las altas frecuencias relativas en la mayoría de las poblaciones y junto con ser precursora de 12 de los arreglos conocidos (30% del total), sirve de enlace con la especie hermana D. miranda. Se apoya así la hipótesis de que TL puede ser el arreglo ancestral de la especie. Estudios moleculares como el de Schaeffer et al. (2003) apoyan esta hipótesis. Asimismo el elevado número de inversiones presentes en algunas localidades, entre 13 y 17, particularmente en la zona entre los estados mexicanos de Michoacán y Zacatecas, llevan a postular como centro de origen a Michoacán (Olvera et al., 1979), pero se requieren más estudios en la región.
En cuanto a aspectos ecológicos se han reconocido gradientes geográficos en la zona central de México (Guzmán et al.,1993 y 2005), en la zona sur (Olvera et al., 2005) y en cuatro zonas (Salceda y Espinosa-Velázquez, 2006a y b). Los últimos autores demostraron que los gradientes se presentaron con diferencias menores de 20 km entre localidades.
En la dinámica del polimorfismo cromosómico se han detectado cambios temporales en algunas poblaciones. Espinoza-Velázquez y Salceda (1981), tras un seguimiento anual de las frecuencias relativas de los arreglos en la localidad de Los Lirios, cercana a Saltillo, Coahuila, detectaron que tres arreglos, Pikes Peak (PP), Olympic (OL) y Tree Line (TL) eran los principales componentes y presentaban frecuencias semejantes a lo largo del estudio. En las localidades de Estados Unidos sólo dos inversiones fueron los principales componentes y las frecuencias inversas conforme transcurre el año, constituyendo un patrón cíclico. En el seguimiento realizado por más de 25 años en Tulancingo, Hidalgo, Amecameca, estado de México, y Zirahuén, Michoacán (datos en proceso), las inversiones participantes fueron Tree Line (TL), Cuernavaca (CU) y Santa Cruz (SC).
D. willistoni es una especie neotropical ampliamente distribuida en América del Sur, que se extiende desde el sur de Florida y centro de México hasta Buenos Aires y está presente en las Antillas y varias islas caribeñas (Townsend, 1958). En México Patterson y Mainland (1944) la recolectaron en los estados de Tamaulipas, Veracruz, Morelos, Guerrero, Michoacán y Jalisco, pero según nuestra experiencia debe habitar en otros, al menos en Tabasco. La especie hace parte del grupo willistoni de importancia ecológica, genética y evolutiva; se le considera la especie más polimórfica del género Drosophila, con 70 inversiones descritas (Valente y Morales, 1985). Dobzhansky (1950) describió el mapa cromosómico fundamental de la especie y posteriormente en colaboración con investigadores brasileños inició descripciones de inversiones en buen número de localidades. Entre otros trabajos tenemos los de Da Cunha et al. (1950) relacionando polimorfismo cromosómico con adaptación, Da Cunha y Dobzhanskysky (1954) relacionando el polimorfismo con el medio ambiente, Dobzhansky (1957) refiriéndose a poblaciones continentales e insulares, Townsend (1952 y 1958) con referencia a poblaciones marginales y caribeñas, Valente (1980 y 1981) relacionando la variabilidad cromosómica con ambientes y recursos tróficos. No ha faltado el estudio de la variabilidad mediante técnicas bioquímicas (Ayala et al., 1970).
En México los estudios de esta índole son incipientes. Salceda (2001), en una población tabasqueña de 19 hembras, detectó 27 inversiones repartidas en los cinco brazos del genoma politécnico, tres inversiones A del cromosoma XL con frecuencia de 15.8 %, E del IIR con 28.9 % y B del III con 21.0 %. Las restantes 24 alcanzaron frecuencias inferiores al 10 % (Tabla 2).
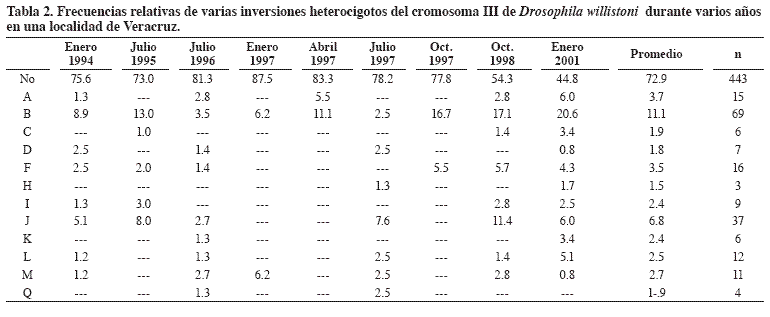
Salceda (2005), en un análisis preliminar en cinco localidades del oriente de México con tamaños de muestra desde cinco hasta 114 hembras, entre 202 hembras detectó siete inversiones en el cromosoma XL, seis en el XR, siete en el IIL, seis en el IIR y 12 en el III (54.3% de las conocidas). En un estudio que se prolongó por ocho años en una localidad y haciendo un seguimiento semestral, Salceda (2006) detectó en 319 hembras y 3.190 cromosomas 45 inversiones (64.3%) en los cinco brazos del cariotipo. El número de inversiones portadas por hembra se localizó entre 1.9 y 3.7, lo que incidió en los valores representativos de zonas marginales de la distribución de la especie. Como la zona cubierta por los estudios fue de 400 km y el polimorfismo cromosómico es de índole adaptativo se deben estudiar más localidades, considerando la alta diversidad ecológica existente en México.
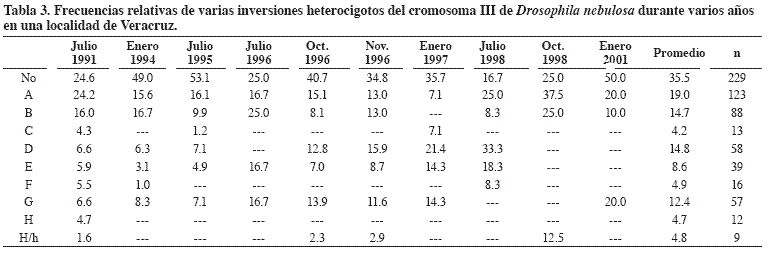
D. nebulosa, especie neotropical que se extiende desde Buenos Aires hasta México y Texas (Patterson y Mainland, 1944), es el miembro más común del grupo willistoni en México, donde se ha recolectado en los estados de Coahuila, Durango, Guerrero, Jalisco, México, Michoacán, Morelos, Nuevo León, Oaxaca, Puebla, San Luis Potosí y Veracruz. El mapa cromosómico de D. nebulosa fue descrito por Pavan (1946) señalando la existencia del polimorfismo en los brazos XL y III. En el brazo XL se encuentra una inversión en la extremidad del cromosoma sexual y es semejante a la “sex-ratio” de D. pseudoobscura sin presentar las consecuencias de la misma. El cromosoma III presenta 17 inversiones paracéntricas en general de pequeño tamaño y son el producto de ocho secciones invertidas. La especie ha sido poco estudiada a este respecto. Da Cunha (1953) reportó la frecuencia de inversiones heterocigotas en diez poblaciones naturales en Brasil, incluyendo otro parámetro de estudio, el número promedio de inversiones por hembra. Valente et al. (1996) reportaron el hallazgo de una nueva inversión en el segundo cromosoma (IIR).
En poblaciones mexicanas Salceda (2005) estudió ocho años una localidad de Veracruz (Tabla 3), analizó 1.270 cromosomas y correspondieron 635 para cada uno de los dos pares cromosómicos en los que existe polimorfismo. Las frecuencias fueron similares en cada muestreo y las inversiones de mayor frecuencia relativa fueron: A (19.0%), D (14.8%), B (14.7%) y G (12.4%). La inversión IIL se detectó con una frecuencia promedio de 12.5%. Recolectas recientes en la costa oriental de México representativas de seis localidades se encuentran en proceso de análisis. El número promedio de inversiones por hembra fue de 1.57.
AGRADECIMIENTOS
El autor desea expresar su agradecimiento a las autoridades del ININ por el continuo apoyo a este proyecto. Al CONACyT (Consejo Nacional de Ciencia y Tecnología de México), por el apoyo financiero que permitió parcialmente la realización de recolectas y desarrollo del proyecto mediante convenio 31736-N. Muy particularmente a la doctora Creucí María Caetano por la amable invitación al evento y a las autoridades de la Universidad Nacional de Colombia, Sede Palmira, por cubrir los gastos que me permitieron la asistencia al II Simposio Latinoamericano de Citogenética y Evolución (II SLACE, 15-18 agosto de 2007). A todas las personas que de una u otra forma colaboraron al desarrollo de esta investigación. El escrito fue mejorado gracias a los comentarios y sugerencias de un revisor anónimo.
BIBLIOGRAFÍA
1.Ayala, F.J; Mourao, C.A; Perez-Salas, S; Richmond R.C; Dobzhansky, T. 1970. Enzyme variability in the Drosophila willistoni group. I. Genetic differentiation among sibling species. Proc Natl Acad Sci USA, 67: 225-232.
2.Bridges, C.B. 1935. Salivary chromosome maps, with a key to the banding of the chromosomes of Drosophila melanogaster. J Hered 26: 60-64.
3.Brncic, D. 1962. Chromosomal structure of populations of Drosophila flavopilosa studied in larvae collected in their natural breeding sites. Chromosoma 13: 183-195.
4.Carson, H.L; Stalker, H.D. 1947. Gene arrangements in a natural population of Drosophila robusta, Sturtevant. Evolution 1: 113- 123.
5.Da Cunha, A.B; Brncic, D; Salzano, F.M. 1953. A comparative study of chromosomal polymorphism in certain South American species of Drosophila. Heredity 7: 193-302.
6.Da Cunha, A.B; Burla, H; Dobzhansky, T. 1950. Adaptive chromosomal polymorphism in Drosophila willistoni. Evolution 4: 212-225.
7.Da Cunha, A.B; Dobzhansky, T. 1954. A further study of chromosomal polymorphism in Drosophila willistoni in its relation to the environment. Evolution 8: 119-134.
8.Dobzhansky, T. 1948. Chromosomal variation in populations of Drosophila pseudoobscura which inhabit Northern Mexico. Am Nat, 82: 97-106.
9.Dobzhansky, T. 1950. The chromosomes of Drosophila willistoni. J Hered 41: 156-158.
10.Dobzhansky, T. 1957. Genetics of natural populations. XXVI. Chromosomal variability in island and continental populations of Drosophila willistoni from Central America and West Indies. Evolution 11: 280-293.
11.Dobzhansky, T; Sturtevant, A.H. 1938. Inversions in the chromosomes of Drosophila pseudoobscura. Genetics 23: 28-64.
12.Dobzhansky, T; Epling, C. 1944. Contributions to the genetics, taxonomy and ecology of Drosophila pseudoobscura and its relatives. Carnegie Institution of Washington, Publication 554, 183p.
13.Dobzhansky, T; Hunter, A.S; Pavlovsky, O; Spassky, B; Wallace, B. 1963. Genetics of natural populations. XXXI. Genetics of an isolated marginal population of Drosophila pseudoobscura. Genetics, 48: 91-103.
14.Dubunin, N.P; Tiniakov, G.G. 1945. Seasonal cycles and the concentration of inversions in populations of Drosophila funebris. Am Nat 79: 570-582.
15.Dobzhansky, T ; Felix, R; Guzmán, J ; Levine, L; Olvera, O; powell, J.R; de la Rosa, M.E; Salceda, V.M. 1975. Population genetics of Mexican Drosophila. I. Chromosomal variation in natural populations of Drosophila pseudoobscura from Central Mexico. J Hered 66: 203-205..
16.Espinoza-Velázquez, J; Salceda, V.M. 1981. Inversion polymorphism of a Drosophila pseudoobscura population from Saltillo, Coah. Mexico. Southwest Nat 25 (4): 479-483.
17.Goldschmidt, E. 1956. Chromosomal polymorphism in a population of Drosophila subobscura from Israel. J Genet 54: 474-481.
18.Guzmán, J; Olvera, O; de la Rosa, M.E; Salceda, V.M. 1993. East-West distribution of inversion polymorphism in Drosophila pseudoobscura. Southwest Nat 38: 52-57.
19.Guzmán, J; Salceda, V.M; Olvera, O; Levine, L. 2005. Geographical gradients in Mexican populations of Drosophila pseudoobscura. Rev Int Contam Ambient 21 (Supl 1): 21-25.
20.Kastritsis, C.D; Crumpacker, D.W. 1966. Gene arrangements in the third chromosome of Drosophila pseudoobscura. I. Configurations with tester chromosomes. J Hered 57: 150-158.
21.Kastritsis, C.D; Crumpacker, D.W. 1967. Gene arrangements in the third chromosome of Drosophila pseudoobscura. II. All possible configurations. J Hered 58: 112-129.
22.Koller, P.C. 1936. Structural hybridity in Drosophila pseudoobscura. J Genet 32: 79-84.
23.Levine, L; Olvera, O; Powell, J.R; Rockwell, R.F; de la Rosa, M.E; Salceda, V.M; Anderson, W.W.; Guzmán, J. 1995. Studies on Mexican Drosophila pseudoobscura. In: Levine, L. (ed). Genetics of Natural Populations: The Continuing Importance of Theodosius Dobzhansky. Columbia University Press. p 109- 139.
24.Olvera, O; de la Rosa, M.E; Salceda, V.M; Gaso, M.I; Guzmán, J. 1982. Population genetics of Mexican Drosophila. VII. Inversion polymorphism in populations of Drosophila pseudoobscura from Southern Mexico. Southwest Nat 27: 361-364.
25.Olvera, O; Powell, J.R; de la Rosa, M.E; Salceda, V.M; Gaso, M.I; Guzmán, J; Anderson, W.W; Levine, L. 1979. Population genetics of Mexican Drosophila. VI. Cytogenetic aspects of the inversion polymorphism in Drosophila pseudoobscura. Evolution 33: 381-393.
26.Olvera, O; Salceda, V.M; Levine, L; Guzmán, J. 2005. Chromosomal polymorphism of Drosophila pseudoobscura from Southern Mexico. Rev Int Contam Ambient 21, (Supl. 1).27-30.
27.Painter, T.S. 1934. A new method for the study of chromosomal aberrations and the plotting of chromosome maps in Drosophila melanogaster. Genetics 19: 175-188.
28. Pavan, C. 1946. Chromosomal variation in Drosophila nebulosa. Genetics 31: 546-557.
29. Patterson, J.T; Mainland, G.B. 1944. The Drosophilidae of Mexico. The University of Texas Publication No. 4445, 101p.
30. Salceda, V.M. 2001. Inversion polymorphism of Drosophila pseudoobscura (Drosophilidae) from the Baja California Peninsula. Dros Inform Serv 84: 95-99.
31. Salceda, V.M. 2001. Inversion polymorphism in a natural population of Drosophila willistoni from Tabasco, Mexico. Dros Inform Serv 84: 99-100.
32. Salceda, V.M. 2005a. Chromosomal polymorphism in natural populations of Drosophila willistoni from Eastern Mexico. Preliminary report. Dros Inform Serv 88: 30-33.
33. Salceda, V.M. 2005b. Inversion polymorphism in a natural population of Drosophila nebulosa. A ten year record. Rev Int Contam Ambient 21 (Supl.1): 31-34.
34. Salceda, V.M. 2006. Chromosomal polymorphism in anatural population of Drosophila willistoni (Diptera: Drosophilidae) from Laguna Verde, Veracruz, Mexico. An eight-year record. Southwest Nat 51 (3): 320-325.
35. Salceda, V.M; Espinoza-Velázquez, J. 2006a. Gradientes geográficos para inversiones del cromosoma III de Drosophila pseudoobscura (Diptera: Drosophilidae) de México. Folia Entomol Mex 45 (1): 17-26.
36. Salceda, V.M; Espinoza-Velázquez, J. 2006b. Microgeographic variation of inversions in natural populations of Drosophila pseudoobscura. Genetika 38(2): 97-106.
37. Salceda, V.M; Guzman, J; Olvera, O; Levine, L. 2007a.Chromosomal variation in natural populations of Drosophila pseudoobscura inhabiting Northern Mexico. Southwest Nat 52(3): 430-435.
38. Salceda, V.M; Guzmán, J. and Olvera, O. 2007b. Inversion polymorpism in some natural populations of Drosophila pseudoobscura from Central Mexico. Genetika 39(3): 343-354.
39. Schaeffer, S.W; Goetting-Minesky, M.P; Kovacevic, M; Peoples, J.R; Graybill, J.L; Miller, J.M; Kim, K; Nelson, J.G; Anderson, W.W. 2003. Evolutionary genomics of inversions in Drosophila pseudoobscura: Evidence for epistasis. Proc Natl Acad Sci USA 100: 8319-8324.
40. Sturtevant, A.H. 1917. Genetic factors affecting the strength of linkage in Drosophila. Proc Natl Acad Sci USA 22: 448-452.
41. Sturtevant, A.H; Dobzhansky, Th. 1936. Inversions in the third chromosome of wild races of Drosophila pseudoobscura, and their use in the study of the history of the species. Proc Natl Acad Sci USA 22: 448-450.
42. Sturtevant, A.H.; Plunkett, C.R. 1926. Sequence of corresponding third chromosome genes in Drosophila melanogaster and D. simulans” Biol Bull 50: 56-64.
43. Tan, C.C. 1935. Alivary gland chromosomes in the two races of Drosophila pseudoobscura. Genetics 20: 392-402.
44. Townsend, J.I. 1952. Genetics of marginal populations of Drosophila willistoni. Evolution 6: 428-442.
45. Townsend, J.I. 1958. Chromosomal polymorphism in Caribbean Islands populations of Drosophila willistoni. Proc Natl Acad Sci USA 44: 38-42.
46. Valente, V.L.S. 1980. Observacoes sobre diferencas no polimorfismo cromossômico de linhagens de Drosophila willistoni em paralelo com exploracao de diferentes recursos tróficos. Ciencia e Cultura 32 (Supl.) 7: 668.
47. Valente, V.L.S. 1981. polimorfismo cromossômico e variabilidade ambiental em populacoes naturais de Drosophila willistoni. Arch Biol Med Exp 14 (1):87.
48. Valente, V.L.S; Morales, N.B. 1985. New inversions and qualitative description of inversion heterozygotes in natural populations of Drosophila willistoni inhabiting two different regions in the State of Rio Grande do Sul, Brazil. Rev Bras Genet 7 (1): 167-173.
49. Valente, V.L.S; Bonorino, C.B; Goñi, B. 1996. Photomap of Drosophila nebulosa Sturtevant, with description of a new inversion inpopulations from Uruguay. Braz J Genet 19 (1): 93-96.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2008 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.





















