



Publicado
Efecto de aportes superficiales de biomasa vegetal en la temperatura, humedad y dinámica de nemátodos en el suelo en época seca en Santander de Quilichao (Departamento del Cauca)
Palabras clave:
Suelo, cobertura arbórea, nemátodos, biomasa, factores ambientales. (es)Soils, trees cover, nematodes, biomass, decomposition, environmental factors. (en)
|
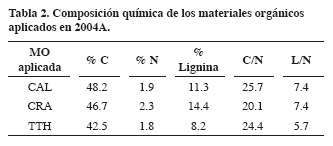
Efecto de aportes superficiales de biomasa vegetal en la temperatura, humedad y dinámica de nemátodos en el suelo en época seca en Santander de Quilichao (Departamento del Cauca).1 Jorge Fernando Navia Estrada,2 Edmundo Barrios,3 Marina Sánchez4 Compendio | Abstract | Introducción | Materiales y Métodos Resultados y Discusión | Conclusiones | Bibliografía En la estación experimental del CIAT (Santander de Quilichao- Cauca, 3°06'N y 76°31'W) se exploró el efecto de aplicaciones superficiales de biomasa vegetal (6 t ms ha-1 semestre-1), después de la siembra de fríjol. El ensayo constó de cuatro tratamientos, tres coberturas de árboles de características contrastantes (Calliandra-CAL; Cratylia-CRA y Tithonia -TTH) y el testigo CON; dispuestos en un DBCA con 5 repeticiones. Las variables evaluadas fueron ambientales (humedad y temperatura del suelo) y biológicas (abundancia y dinámica de los nemátodos). Bajo cobertura se incrementó la humedad en 3% y se reguló la temperatura del suelo de 0.4 a 0.6°C menor que CON. El comportamiento de los nemátodos fue muy dinámico y estuvo relacionado con las condiciones ambientales, siendo favorecido en épocas de humedad y por la dinámica de descomposición relacionada con el tipo de material. En general, la cobertura modificó el microambiente afectando la población de organismos, generando cambios en la aireación, estructura y ciclaje de nutrientes en el suelo por aporte de materia orgánica a través de la hojarasca regulando su disponibilidad en los sistemas productivos. Palabras claves: Suelo, cobertura arbórea, nemátodos, biomasa, factores ambientales. Effect of superficial contributions of vegetal biomass in the temperature, humidity and dynamics of nematodes in the soil at a season dry time in Santander Quilichao (Department of the Cauca). In the experimental station at CIAT (Santander de Quilichao- Cauca, to 3°06'N and 76°31'W), the effect of surface application of vegetal biomass was explored (6 t dm ha-1 semester-1), after planting of bean. The experiment consisted of four treatments, three cover of the trees that had contrasting characteristics (Calliandra-CAL; Cratylia-CRA and Tithonia - TTH) and the control- CON; arranged in a CBRD with 5 repetitions. The evaluated variables were: environmental (humidity and temperature of the soil) and biological (abundance and dynamics of the nematodes). Under cover the humidity in a 3% was increased and the soil temperature was regulated from the 0.4 to 0.6°C smaller than the CON treatment. The behavior of the nematodes was very dynamic and was related to the environmental conditions, being favored at wet seasons and by the dynamics of decomposition related to the type of material. In general, the cover modified the microenvironment affecting the population of organisms, generating changes in the aireation, structures and nutrients cycling in the soil by contribution of organic matter through mulch which regule its availability in the productive systems. Key words: Soils, trees cover, nematodes, biomass, decomposition; environmental factors. Una de las principales fuentes de materia orgánica del suelo es el tejido vegetal. En condiciones naturales la biomasa aérea y las raíces de árboles, arbustos, hierbas proveen residuos orgánicos. La cantidad y calidad de la MOS está influenciada por la naturaleza de las adiciones superficiales o incorporadas de diferentes residuos orgánicos, sumada a las condiciones ambientales propias de cada región (Swift y Woomer, 1993). La tasa de descomposición de la MOS está en función de diversas características y procesos incluyendo la composición química o calidad del material orgánico, la temperatura y humedad del sustrato y la composición de la comunidad descomponedora (Heal et al., 1997; Neher et al., 2003). Su velocidad de descomposición, por lo general, es alta hasta que desaparecen los compuestos de fácil degradación (proteínas, almidones y azúcares). Las biomoléculas de celulosa, hemicelulosa y lignina se degradan más lentamente, pero brindan barreras de protección al suelo y posibilidades de almacenamiento o acumulación de materia orgánica (Wardle y Lavelle, 1997). Generalmente los aportes orgánicos a partir de leguminosas son mejores debido a que tienen mayores contenidos de N y relaciones C/N más bajas; aunque existen excepciones como Tithonia diversifolia (Barrios y Cobo, 2004). El contenido de N se usa frecuentemente como el principal criterio de selección de aportes orgánicos como fuente de nutrientes, pero se han propuesto otros índices para predecir las tasas de descomposición y liberación de N, que incluyen las relaciones Lignina/N o polifenol/N y (lignina+polifenol)/N (Barrios et al., 1997; Vanlauwe et al., 1997; Wachendorf, et al., 1997; Cobo et al., 2002). Dentro de la comunidad de los organismos del suelo, los macroorganismos se encargan de desmenuzar los restos vegetales y animales para que los microorganismos los descompongan y los conviertan en fuente de nutrientes para las plantas (Lavelle y Spain, 2001). Los microorganismos son los agentes primarios para la inmovilización de nutrientes, debido a que los utilizan en el crecimiento y reproducción (Gupta y Yeates, 1997). Estudios recientes confirman los efectos de los invertebrados del suelo en la estructura, en la alteración de los patrones de la actividad microbiana y la influencia en la dinámica de la materia orgánica y ciclaje de nutrientes (Lee y Pankhurst, 1992; Lavelle y Spain, 2001). Varios estudios indican que la abundancia y composición de los nemátodos del suelo son una medida indirecta de la comunidad microbiana del suelo y pueden proveer información adicional de los procesos (Freckman y Casswell, 1985; Ingham, 1994; Gupta y Yeates, 1997; Coleman et al., 1999). Los nemátodos son muy sensibles a las prácticas de manejo del suelo (Yeates et al., 1993; Yeates y Bird, 1994), pues actúan como consumidores de microflora (bacterias, hongos) y como saprófitos, principalmente, mediando la descomposición y liberación de nutrientes a las plantas (Freckman y Ettema, 1993). Actualmente ha aumentado el interés por profundizar en el conocimiento del papel de los nemátodos en procesos de los agroecosistemas tales como ciclaje de nutrientes, en el control biológico y como indicador de manejo de suelos que afectan la abundancia, diversidad y función de esta población y por ende en el ecosistema donde actúa (Yeates y Bird, 1994; Ekschmitt et al., 1999). La temperatura afecta oviposición, reproducción, movilidad, desarrollo y supervivencia y la mayoría de los nemátodos herbívoros se tornan inactivos entre 5 y 15 °C y mayores de 35° C (Freckman, 1982) y con relación a los niveles de humedad, el óptimo para su desarrollo está entre 40% y 60 % de la capacidad de campo de los suelos; pero, cuando el suelo se encuentra demasiado seco, las poblaciones disminuyen considerablemente y solo sobreviven los huevos y en condiciones de anegación, por la falta de oxígeno y algunas toxinas generadas por organismos anaeróbicos, se reducen las poblaciones (Freckman, 1982, Freckman y Ettema, 1993). Para avanzar en dicha dirección, en el presente trabajo se exploró el efecto de aplicaciones superficiales al suelo de biomasa vegetal, con características contrastantes, sobre la temperatura y humedad del suelo y su relación con la dinámica y abundancia de nemátodos. El ensayo se realizó en la Estación experimental del CIAT, municipio de Santander de Quilichao, departamento del Cauca (3°06'N y 76°31'W). El sitio presenta una altitud de 990 m temperatura promedia anual de 23°C, precipitación anual promedia de 1800 mm/año, con dos temporadas lluviosas (marzo-junio; septiembre-diciembre), humedad relativa 78% y evapotranspiración potencial de 1711 mm/año. Los suelos son profundos, con un pH de 4.5 a 4.9; de baja fertilidad natural, y están clasificados como Ultisoles con características Palehumult ortóxico, arcilloso, caolinítico, isohipertérmico. La investigación hace parte del macroproyecto “Sistema Integrado de Manejo de la Fertilidad del suelo, de Enfermedades y Plagas” destinado a estudiar el manejo indirecto de hongos patógenos del suelo favoreciendo organismos benéficos como los nemátodos de vida libre. El experimento persiguió estudiar el impacto de la aplicación de biomasa vegetal, de diferentes calidades, sobre los nemátodos. El ensayo se diseñó en bloques completos al azar (BCA), con 4 tratamientos y 5 repeticiones por tratamiento (Tabla 1). Los bloques se localizaron con base en un plano georreferenciado, teniendo en cuenta las diferencias de fertilidad detectadas en el suelo, con base en los niveles de porcentaje de carbono (% C), potasio (K), fósforo (P), calcio (Ca) y magnesio (Mg). El área de cada parcela fue de 30 m2, 5 surcos separados 0.60 m, el número de plantas por surco fue alrededor de 80. La variedad utilizada fue fríjol (Phaseolus vulgaris) V-70 (arbustiva, susceptible a Macrophomina). En la parcela control no se aplicó ningún material orgánico. En todas las parcelas se realizaron las mismas labores de manejo del cultivo del fríjol. La tasa de descomposición de Calliandra en Pescador (Cauca) y en el invernadero de CIAT (Palmira) fue menor que la de Cratylia (0.009), y Thitonia (0.037) por la textura de las hojas (Cobo et al., 2002). El contenido de carbono (C) en CAL fue más alto (Tabla 2), mientras que en TTH fue más bajo; CRA presentó más alto contenido de N, mientras en los otros dos materiales fue similar; el contenido de lignina fue más alto en CRA y más bajo en TTH. Los tres materiales presentaron altas relaciones de C/N, por encima de 20, lo que hace que la descomposición no sea tan rápida; aunque en TTH la más baja relación L/N facilitará la descomposición en el tiempo.
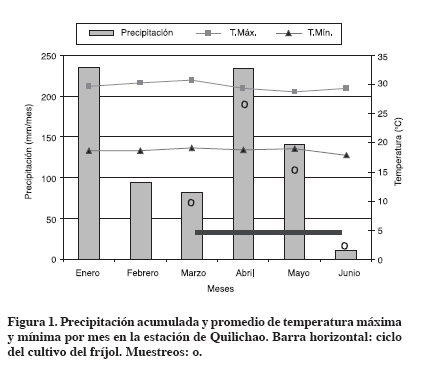
Variables evaluadas Durante el semestre 2004-A la temperatura se tomó a las 8 a.m. (mínima) y a la 1 p.m (máxima). La humedad se tomó semanalmente en cada una de las repeticiones a una profundidad de 5 cm y se expresó como humedad gravimétrica. También la humedad se midió cada vez que se realizó el muestreo de suelos para efectuar las correcciones de humedad a base seca de los datos tomados. Los datos de precipitación y temperatura máxima y mínima diarias de la estación experimental de Santander de Quilichao del CIAT, se relacionaron con los datos de microambiente generados en el experimento. Se realizaron cuatro muestreos (10 cm) para estimar la abundancia y la dinámica de la población de nemátodos. Los tiempos de muestreo fueron antes de la siembra (0), a las 3, 6 después de aplicados los aportes y 11 semanas (a la cosecha). Se tomaron porciones de suelo en el surco, cerca a las plantas (rizósfera), así como también entre los surcos. La muestra de suelo rizosférico se tomó en los 3 surcos centrales de cada parcela y para suelo no-rizosférico se tomó de los dos entresurcos centrales (21 submuestras). Para estimar la población (abundancia) de nemátodos se empleó la técnica de centrifugado (Metodología ASA-Ingham, 1994): se pesaron 300 g y se lavó la muestra a través del tamiz #18 de (1000μm), dentro de un balde, agitando por un minuto. Se pasó la suspensión por tamiz #500, con 25μm, sin permitir el taponamiento, aplicando golpes laterales. Luego se pesó de 16 a 18 g del suelo retenido en el tamiz #500 por tubo y se completó con agua hasta 1 cm antes del tope y se llevó a la centrífuga. Se centrifugaron los tubos por 4 minutos a 1800 rpm y se decantó la mitad del sobrenadante, sin perturbar el pellet (sacar 15 cm3 de la solución sobrenadante). Se añadieron 15 cm3 de una solución de agua azucarada al 90%. Se selló con papel encerado (microfilm) y agitó con mezclador tipo vortex . Se centrifugó a 1800 rpm por 1 minuto. Se pasó el sobrenadante por el tamiz #20 con 53μm, colocado dentro de un recipiente con agua en un ángulo de 45° y este líquido lavado se pasó a una caja petri. Luego la caja de Petri se colocó por pocos segundos en baño María (60°C), para relajar los nemátodos. Después se pasó al estereoscopio para el conteo de cada repetición. La abundancia se determinó en kg de suelo peso seco (No. individuos kg-1 ss). Se realizó análisis de varianza para observar las diferencias entre tratamientos con respecto a la humedad y temperatura. También se usaron pruebas de LSD para separar los promedios de los tratamientos en cada variable evaluada utilizando un ≤ 5%. Todos los análisis se realizaron por medio del paquete estadístico SAS (SAS Institute Inc., 1999 - 2000). Situación climática del ensayo En el semestre 2004-A se presentó menos precipitación (467mm) que en años anteriores (superiores a 600 mm). Los meses más lluviosos fueron enero y abril, mes que coincidió con el segundo muestreo; en todas las épocas de muestreo se presentaron diferentes comportamientos de la precipitación (Figura 1).
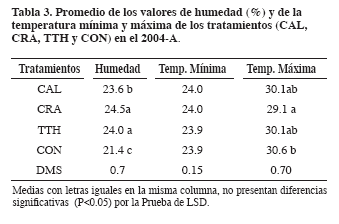
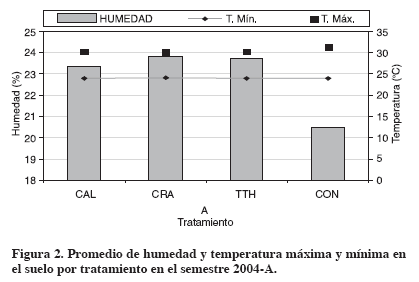
La temperatura máxima disminuyó en los meses de alta precipitación y la mínima en los meses secos. Las temperaturas máxima y mínima oscilaron en forma similar durante el semestre, presentando un rango entre 28.7°C y 30.7°C y de 17.8°C a 19.1°C respectivamente. La temperatura al renovar la masa del aire del suelo influye en la generación de diferentes gradientes de temperatura lo que produce intercambios de flujos de aire especialmente en la capa superficial (Lavelle y Spain, 2001); además, la infiltración de la precipitación en el suelo también genera procesos de renovación del aire a través del desplazamiento del agua por medio de los espacios porosos durante el proceso de redistribución, por lo tanto, el aire del suelo es enriquecido por el O2, que ha sido disuelto por el agua infiltrada e intercambiada más tarde por el aire del suelo (Schnurer et al., 1986; Amézquita, 1994). Comportamiento de la temperatura y humedad del suelo En el comportamiento de la humedad (Tabla 3, Figura 2) se presentó efecto significativo de los aportes orgánicos; siendo los más húmedos CRA y TTH. La cobertura con CRA en general fue la más regular o uniforme, por la biomasa y las grandes hojas el grado de cobertura permaneció mayor tiempo, también, la parte leñosa demora en descomponerse, lo que favoreció en esta época seca, y especialmente a finales de junio, la regulación de la humedad.
En la temperatura mínima no se presentaron efectos significativos por la aplicación de aportes orgánicos. Mientras en la temperatura máxima se presentaron efectos significativos, ya que con CRA fue menor y menos variable en el periodo, seguida de CAL y TTH con similar comportamiento. La temperatura fue más uniforme con Cratylia porque la cobertura permaneció más tiempo; TTH y CAL estuvieron por encima, posiblemente porque el material orgánico estaba en descomposición y se elevó la temperatura por mayor actividad de los organismos del suelo. El tratamiento CON siempre estuvo por encima de los demás, a excepción de la época de cosecha que presentó condiciones de humedad por encharcamiento (junio) (Figura 2).
Con los aportes orgánicos aumenta la humedad y a través del perfil se reduce la temperatura del suelo cuando se aplican materiales de poda de árboles (Scholes et al., 1994). La descomposición de los residuos ubicados en la superficie es a menudo más baja que cuando se incorporan en el perfil de suelo, debido a que la humedad es inferior a la óptima y el contacto de los residuos con el suelo favorece la descomposición. Por ejemplo, Amézquita, 1994, comentó que en un suelo desnudo a 1 cm la temperatura fue de 33°C y con cobertura de tamo de maíz fue de 22°C y en perfil hasta los 20 cm fue de 22°C, mientras sin cobertura fue de 27°C. Los residuos vegetales sobre el suelo producen un efecto aislante, disminuyendo sensiblemente la cantidad de calor que pudiera incidir en los ascensos de temperatura (Woomer et al., 1994; Amézquita, 1994). En este estudio los más rápidos en mineralizarse fueron TTH y CRA y el más lento CAL. La alta descomposición incrementa la actividad de la biomasa microbiana (Phiri et al., 2003; Cobo et al., 2002) aumenta la disponibilidad de nutrientes y la regulación de los gradientes de temperatura y humedad del suelo (Palm, et al., 2001; Barrios y Cobo, 2004). En cuanto a la calidad de la cobertura, los residuos que tienen relación C/N alta, mayor de 20, generan menos energía y las temperaturas se mantienen constantes y el carbono se perderá en forma de dióxido de carbono, a menos que estén combinados con fuentes de N a causa de la inmovilización neta por microorganismos (Swift y Woomer, 1993). En general, la actividad de los organismos desintegradores es baja, debido a que las producciones de CO2 se reducen al mínimo (Myers et al., 1994). También, los materiales ricos en N, presentan cociente bajo de C/N, por tener alto contenido inicial de N y estimulan la tasa de descomposición y son los que más rápidamente se descomponen (Swift y Woomer, 1993). Abundancia y dinámica de la población de nemátodos En términos generales hubo mayor abundancia en el surco que
en el entresurco (Tabla 4); aunque similar comportamiento debido a la
mayor actividad en la rizosfera. El tratamiento TTH fue el de mayor
abundancia y más alta variabilidad entre los materiales de aportes
orgánicos; mientras CRA fue un poco superior a CAL no presentando
diferencias significativas y CON fue el más bajo (Tabla 4). Con relación al surco, TTH fue la más
abundante (311 ind kg-1 ss); posiblemente por ser el material de más
rápida descomposición; seguida de CRA (211 ind
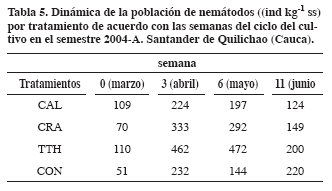
Schnurer et al., 1986, en condiciones similares al estudio, registraron que en épocas secas disminuye la abundancia y aparecen más los herbívoros, ya que al disminuir el potencial del agua se reduce la difusión de solutos y el llenado de poros por agua se incrementa en aireación (Villenave et al., 2003). También en verano se incrementan las tasas de reproducción y los tiempos de generación serían más cortos y aparecerían especies como los bacteríovoros por el incremento de la tasa de descomposición (Yeates, 1979). El total de nemátodos aumentó con la precipitación en todos los tratamientos con aportes, como se registró en la tercera semana (Tabla 5) lo cual afirma lo registrado por varios autores (Yeates, 1979; Ferris y Matute, 2003; Papatheodorou et al., 2004).
En todos los tratamientos se incrementó la población a partir de la primera semana (Tabla 5), por los efectos de la descomposición y de la siembra del fríjol; después de la sexta semana disminuyó drásticamente la población debido al incremento de la temperatura y a la disminución de la precipitación, generando condiciones de humedad muy bajas, las cuales son contrastantes con las requeridas por los nemátodos (Ferris et al., 2001). También se observó el efecto de las aplicaciones de los aportes orgánicos en el tiempo con base en la descomposición; debido a que entre la tercera y sexta semana hubo más abundancia, principalmente en los muestreos abril (3) y mayo (6) que mostraron muy significativas diferencias. En el material de rápida descomposición (TTH) se presentó mayor abundancia que en los otros dos y con CAL se incrementó en el tiempo por ser un material de lenta descomposición. En marzo (semana 0), época donde aún no se ha sembrado y no se ha aplicado el material, el tratamiento TTH fue el más abundante y CAL se comportó mejor que CRA. En abril (semana 3), cuando TTH se ha descompuesto más del 30%, presentó mayor abundancia y CRA mostró mejor comportamiento que CAL. Además, CON superó a CAL, posiblemente por la lenta descomposición del material. En mayo (semana 6), época de floración, los organismos disminuyeron en todos los tratamientos, pero TTH siguió conservando el mejor comportamiento. En la época de recolección a finales de junio (semana 11), antecedieron tres semanas con alta sequía y temperatura, por lo tanto, es posible que estas condiciones hayan llevado a que CON presentara la población más abundante, aunque TTH mostró mejor comportamiento que los otros dos aportes. Papatheodorou et al., 2004 presentaron, con aportes orgánicos, variaciones en la temperatura hasta de 1.4°C y en la humedad en 2.3%. Los cambios en la composición y abundancia de los nemátodos es consecuencia de relaciones fuertes con la microflora edáfica y de cambios fuertes en la humedad y temperatura que afectan la estructura y función de la comunidad microbial y los nemátodos por ser sensibles a esos cambios son indicadores tempranos en los cambios de la MOS (Ferris et al., 2001; Berkelmans et al., 2003; Ferris y Matute, 2003; Neher et al, 2003; Papatheodorou et al., 2004; Wang et al., 2004). Los aportes orgánicos al entrar en descomposición liberan sustancias tóxicas (amoníaco, ácidos fórmico, acético, propiónico y butírico) para muchas especies de nemátodos (Freckman, 1988; Agrios, 1999; Williamson et al., 2005). Por lo tanto la composición química de los residuos de cosecha determina el tamaño y la composición de las comunidades microbianas y faunísticas (Freckman y Ettema, 1993; Freckman y Caswell, 1985; Berkelmans et al., 2003; Sánchez de P., 2003; Williamson et al., 2005). A medida que varía la temperatura en el ambiente y en el suelo se favorecieron diferentes tipos de microorganismos que actúan en diversos procesos de descomposición; además, en sitios húmedos el estado de descomposición es más rápido (Schnurer et al., 1986; Swift y Woomer, 1993). Bongers y Ferris (1999), mostraron respuestas contrastantes de la comunidad de nemátodos a ensayos experimentales, por ejemplo en pasturas no se presentó ningún efecto significativo por cambios de humedad. En otros sitios la temperatura fue más importante para la estructura de la comunidad y la humedad presentó efecto predominante en la abundancia; en otros experimentos se presentaron efectos significativos con respecto a la humedad y épocas de muestreo; lo cual es muy similar con lo reportado en este estudio. Los aportes orgánicos de lenta descomposición (Cratylia y Calliandra) presentaron significativamente efecto en la humedad. Cratylia presentó significativamente menos temperatura máxima por el mayor grado de cobertura. En la dinámica con base en la descomposición la abundancia fue mayor entre la tercera y sexta semana de acuerdo con la calidad del material, Tithonia por la rápida descomposición fue en la tercera semana y en Calliandra fue a partir de la sexta semana por su lenta descomposición. Agrios, G. 1999. Fitopatología. 2 ed. México. Limusa. 838 p Amézquita, E. 1994. Residuos orgánicos superficiales (“mulch”), su importancia en el manejo de suelos. En: Sociedad Colombiana de la Ciencia del Suelo, congreso 7, octubre, 1994. Bucaramanga. Memorias. pp. 9 - 15. Barrios, E.; Kwesiga, F.; Buresh, R.; Sprent, J. 1997. Ligth fraction soil organic matter and available nitrogen following trees and maize. Soil Sci Soc Am J. 61: 826 - 831. –––––.; Cobo, J. 2004. Plant growth, biomass
production and nutrient accumulation by slash/mulch Berkelmans, R.; Ferris, H.; Tenuta, M.; Bruggen, A. 2003. Effects of long - term crop management on nematode trophic levels other tan plant feeders disappear after 1 year of disruptive soil management. Appl Soil Ecol. 23: 223 - 235 Bongers, T.; Ferris, H. 1999. Nematode community structure as a bioindicator in environmental monitoring. Trend Evol Ecol. 14:224-228. Cobo, J.; Barrios, E.; Kass, D.; Thomas, R. 2002. Decomposition and nutrient release by green in tropical hillside. Plant Soil. 240: 331 - 342. Coleman, D.; Blair, J.; Elliot, E.; Wall, D. 1999. Soil invertebrates In: Roberts et al (Eds.). Standard Soil Methods for Long-term ecological research. pp 349 - 377. Ekschmitt, K.; Bakonyi, G.; Bongers, M.; Bongers, T.; Bostrom, S.; Dogan, H.; Harrison, A.; Kallimanis, A.; Nagy, P.; O´Donnell, A.; Sohlenius, B.; Stamou, G.; Wolters, V. 1999. Effects of the nematofauna on microbial energy and matter transformaion rates in European grassland soils. Plant Soil. 212: 45-61. Ferris, H.; Matute, M. 2003. Structural and functional succession in the nematode fauna of a soil food web. Appl Soil Ecol. 23: 93-110. Ferris, H.; Bongers, T.; de Goede de, R. G. 2001. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Appl Soil Ecol. 18: 13-29. Freckman, D.; Ettema, C. 1993. Assessing nematode comunities in agroecosystems of varying human intervention. Agric Ecosys Envi. 45: 239 - 261. –––––; Caswell, E. 1985. The ecology of nematodes in agroecosystems. Ann Rev Phytopath. 23: 275-296 –––––. 1988. Bacterivorous nematodes and organic - matter decomposition. Agric Ecosyst Env. 24: 195-217 –––––. 1982. Nematodes in soil ecosystems. The University of Texas Press. 198 p. Gupta, V.; Yeates, G. 1997. Soil microfauna as bioindicators of soil health. In: Pankhurst, C; Gupta, V. (Eds.). Biological Indicators of Soil health. CABI, Wallingford. pp 201-233. Heal, O.; Anderson, J.; Swift, M. 1997. Plant litter quality and decomposition: an historical overview In: Giller, K.; Cádish, G.; (Eds) Driven by Nature. CABI, Wallingford. pp 3-30. Ingham, R. 1994. Nematodes. In: American Methods of Soil análisis. pp. 459-490. Lavelle, P.; Spain, A. V. 2001. Soil Ecology. Netherlands: Kluver. 654 p. Lee, K.; Pankhurst, C. 1992. Soil organism and sustainable productivity. Austr J Soil Res. 30:855-892 Myers, R.; Palm, C.; Cuevas, E.; Gunatilleke, I.; Brussaard, M. 1994. The Synchronization of nutrient mineralisation and plant nutrient demand. En: Woomer, P.; Swift, M (eds). The biological managemet of tropical soil fertility. Wiley, Chichester. pp 81-116. Neher, D.; Barbercheck, M.; El - Allaf, S.; Anas, O. 2003. Effects of disturbance and ecosystem on decomposition. Appl Soil Ecol. 23: 165 - 179. Palm, C.; Giller, K.; Mafongoya, L.; Swift, M. 2001. Management of organic matter in the tropics: translating theory into practice. Nut Cycl Agroecosist. 61: 63-75. Papatheodorou, E.; Argyropoulou, M.; Stamou, G. 2004. The effects of large and small scale differences in soil temperature and moisture on bacterial functional diversity and the community of baceriovorous nematodes. Appl Soil Ecol. 25: 37-49. Phiri, S.; Rao, I.; Barrios, E.; Sing., B. 2003. Plant grow mycorrhizal association, nutrient uptake and phosphorus dynamics in a volcanic-ash soil in Colombia as affected by the establishment of Tithonia diversifolia. J Sust Agric. 21 (3): 43-61. Sánchez, M de P. 2003. Actividad biológica en la rizosfera del maracuyá en diferentes sistemas de manejo, estados de desarrollo y condiciones fitosanitarias. Tesis Doctoral, Universidad Politécnica de Madrid. 247 p. Scholes, R.; Dalal, R.; Singer, S. 1994. Soil physics and fertility of water, temperature and texture. In: Woomer, P.; Swift, M. (eds). The biological management of tropical soil fertility. Wiley, Chichester. pp 117 - 135. Schnurer, J.; Clarholm, M.; Bostrom, S.; Rosswall, T. 1986. Effects of moisture on soil microorganisms and nematodes: A field experiment. Micr Ecol. 12: 217-230. Swift, M.; Woomer, P.; 1993. Organic matter and the sustainability of agricultural systems: Definition and measurement. In: Woomer, P.; Swift, M. (eds) the biological managemet of tropical soil fertility. Wiley, Chichester. pp 1-12 Vanlauwe, B.; Diels, J.; Sanginga, N.; Merckx, R. 1997. Residues quality and decomposition: An unsteady relationship. In: Cádish, G; Giller, (Eds) Driven by Nature. CABI, Wallingford. pp 157 - 166. Villenave, C.; Cadet, P.; Planchon, O.; Esteve, M.; Lapetite, J. 2003. Transport of free-living nematodes by runoff water in a Sudano-Sahelian Area. Appl Soil Ecol. 23: 85-91. Wachendorf, C.; Irmler, U.; Blume, H. 1997. Relationships between litter fauna and chemical changes of litter during decomposition under different moisture conditions. In: Cadish G, Giller, K (Eds) Driven by Nature. CABI, Wallingford. pp, 135 - 144. Wang, K.; Mcsorley, R.; Marshall, A.; Gallear, N. 2004. Nematode community changes associated with decomposition of Crotalaria juncea amendment in litterbags. Appl Soil Ecol. 27(1):31-45. Wardle, D.; Lavelle, P. 1997. Linkages between soil biota, plant litter quality and decomposition. In: Cadish, G; Giller K (Eds) Driven by Nature. CABI, Wallingford. pp 107 - 124. Williamson, W.; Wardle, D.; Yeates, G. 2005. Changes in soil microbial and nematode communities during ecosystem decline across a long-term chronoquence. Soil Biol Biochem. 37(7):1289-1301. Woomer, P.; Martin, A.; Resck, D.; Scharpenseel, H. 1994. The importance and management of soil organic matter in the tropics. In: Woomer, P.; Swift, M.(eds). The biological managemet of tropical soil fertility. Wiley, Chichester. pp 47-79 Yeates, W.; Bird, A. 1994. Some observations on the influence of agricultural practices on the nematode faunae of some South Australian soils. Fund Appl Nemat 17. (2): 133 - 145. –––––., Bongers, T.; Goede, R.; Freckman, D.; Georgieva, S. 1993. Feeding habits in soil nematode families and Genera - An Outline for Soil Ecologists. J Nemat. 25 (3): 315 - 331. –––––. 1979. Soil nematodes in terrestial ecosystems. J Nemat. 11(3), 213-227.
|


Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2006 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.





















