



Publicado
Descomposición y liberación de nutrimentos desde las raíces finas de los sistemas agroforestales de cacao con sombras de poró y laurel, en Turrialba, Costa Rica
Palabras clave:
Agroforestería, Theobroma cacao, Erythrina poeppigiana, Cordia alliodora, recirculación de nutrimentos, descomposición de raíces finas. (es)Agroforestry, Theobroma cacao, Erythrina poeppigiana, Cordia alliodora, nutrients retranslocate, fine roots decomposition. (en)
Descomposición y liberación de nutrimentos desde las raíces finas de los sistemas agroforestales de cacao con sombras de poró y laurel, en Turrialba, Costa Rica.1 Compendio | Abstract | Introducción | Materiales y Métodos Discusión y Conclusiones | Agradecimientos | Bibliografía Se usó el método de bolsas de descomposición para estudiar la pérdida de biomasa y liberación de N, P, K, Ca y Mg desde las raíces finas (<2 mm) en los sistemas agroforestales de cacao con sombras de poró y laurel. Se muestrearon bolsas a las 1, 2, 4, 6, 12, 18 y 25 semanas en las épocas seca y lluviosa. Las tasas de descomposición y liberación de nutrimentos fueron bien descritas por un modelo exponencial doble. Se observó alta relación entre las tasas de pérdida de biomasa y liberación de N de las tres especies. Las tasas de liberación de nutrimentos fueron K > Mg > P > N > Ca. Después de una semana de incubación cerca del 90% del K fue liberado mientras que el Ca tendió a la acumulación. Las raíces de poró fueron las de más rápida descomposición y liberación de nutrimentos, excepto de K. El poró sería la especie de sombra adecuada para sistemas de producción que requieran rápida recirculación de nutrimentos. El laurel exhibió tasas lentas de descomposición y liberación de nutrimentos. Palabras claves: Agroforestería, Theobroma cacao, Erythrina poeppigiana, Cordia alliodora, recirculación de nutrimentos, descomposición de raíces finas. Decomposition and nutrient release from fine roots of agroforestry systems of cocoa with Erythrina and Cordia in Costa Rica. The litter bag methodology was used to study biomass decay and nutrient release of N, P, K, Ca, and Mg from fine roots (<2mm) in cocoa plantations shaded with poro (Erythrina) and laurel (Cordia) in Turrialba, Costa Rica. Samplings at 1, 2, 4, 6, 12, 18, and 25 weeks in the dry and rainy periods were done. Decomposition and nutrient release rates were well described by a double exponential model. Relation between biomass decay and N release from the fine roots of the three species was observed. The nutrient release rates were : K > Mg > P > N > Ca. After the first week of the incubation period, about 90% of the K was released while Ca tended to accumulate. Poro roots had the fastest decomposition and nutrient release rates except for K which was similar for all of the three species. Poro would be the most suitable specie for systems where a fast recirculation of nutrients is desirable. On the contrary, laurel exhibited slow decomposition and nutrient release rates. Key words: Agroforestry, Theobroma cacao, Erythrina poeppigiana, Cordia alliodora, nutrients retranslocate, fine roots decomposition. La descomposición de la materia orgánica juega papel importante en el ciclo de los nutrimentos en ecosistemas naturales y agroecosistemas. El proceso permite que recirculen y que sean potencialmente reutilizables los nutrimentos acumulados en la biomasa muerta. La descomposición es afectada principalmente por factores abióticos como la temperatura y la humedad, por lo tanto en el trópico donde la temperatura es relativamente alta y constante, la humedad es el factor climático que más influye sobre la descomposición (Anderson y Swift, 1983; Fabiao et al., 1985). También influyen las características fisico-químicas del material vegetal, dureza física, contenido de lignina, contenido de nutrimentos, substancias secundarias, masa y tamaño de partículas ; además de los organismos descomponedores presentes en el medio edáfico y en la biomasa acumulada sobre el suelo (Anderson y Swift, 1983). Aunque la descomposición de las raíces finas y de la hojarasca son la principal fuente vegetal de recirculación de nutrimentos, en algunos ecosistemas se ha estimado mayor aporte de materia orgánica al suelo a partir de la descomposición de las raíces finas (Persson, 1979). En sistemas agroforestales de cacao (Theobroma cacao) la proporción de materia orgánica aportada por las raíces finas (diámetro < 2 mm) es menor aunque faltan estudios que estimen la ventaja comparativa de la descomposición bajo el suelo versus la descomposición de la hojarasca (Muñoz y Beer, 2001). En los sistemas de producción agrícola donde es muy baja la diversidad de especies y de sistemas radiculares aumenta la probabilidad de pérdidas por lavado y/o lixiviación de los nutrimentos provenientes de la descomposicion. Ante esta situación se vuelve importante conocer la dinámica del proceso de descomposición con el objetivo de diseñar agroecosistemas y afinar las estrategias de manejo de los ya existentes de manera que se pueda obtener máxima eficiencia en el uso y mínima pérdida de los nutrimentos. Por consiguiente, el conocimiento de la tasa de descomposición de las raíces finas en los sistemas agroforestales es un factor clave ya que al analizarlas en conjunto con variables como tasas de mineralización, periodos de máxima absorción de las especies, producción y biomasa total de raíces finas y hojarasca, permitirán conocer la dinámica de los nutrimentos en un sistema y por lo tanto tratar de asegurar su sostenibilidad (Fassbender, 1993). La descomposición de raíces finas en sistemas agroforestales ha sido poco estudiada. El presente estudio tuvo como principal objetivo aportar conocimientos sobre la dinámica de la liberación de nutrimentos a partir de la descomposición de las raíces finas en los sistemas agroforestales de cacao con sombras de laurel (Cordia alliodora) y poro (Erythrina poeppigiana) en Turrialba, Costa Rica. El sitio experimental está localizado en Turrialba, Costa Rica a una altura de 600 m, 21.5˚C y una precipitación de 2660 mm año-1 . El suelo se clasifica como Andic Eutropept, franco, mezclado, isohipertérmico en la taxonomía americana y como Eutric Cambisol en el sistema FAO/UNESCO. La pluviosidad y tensión de humedad en el suelo fueron medidos in situ y reportados en el estudio de tasas de producción de biomasa de raíces finas (Muñoz y Beer, 2001) conducido simultáneamente con este estudio y se usaron los mismos periodos (alta y baja precipitación) definidos en ese estudio. Se usó el mismo diseño experimental de Muñoz y Beer, 2001. En el estudio se modificó el método de bolsas de descomposición (Steen, 1985; Noorwijk, 1992). Al inicio de cada periodo se recolectaron raíces vivas de cada especie (diámetro <2 mm) de las áreas de borde de cada parcela y se identificó el origen para colocarlas nuevamente en la parcela correspondiente. Las raíces se lavaron y secaron al ambiente por 72 horas. Antes de colocarlas dentro de cada bolsa se humectaron para que recuperaran flexibilidad y así evitar pérdidas por manipuleo. Dentro de cada bolsa se colocaron 0.6 g (peso fresco) de raíces (0.3 g de cacao y 0.3 g de poró o laurel, de acuerdo con el sistema). Se utilizaron bolsas de malla plástica de abertura variable (máx 5 mm) que tuvieron la ventaja de que al tirar del extremo para recuperarlas la abertura tendió a cerrarse y así se evitó la pérdida de fragmentos de raíz. Para colocar las bolsas en el suelo, se abrió una ranura de 15 cm de profundidad por 3 cm de ancho. Luego, con la ayuda de una regla plástica, se empujó la bolsa hasta el fondo de la ranura, finalmente se abrió otra ranura paralela a la primera y se empujó el suelo hacia la ranura que contenía la bolsa con raíces, de esta manera se logró buen contacto entre el suelo y las raíces. Las bolsas enterradas se cubrieron con hojarasca para simular las condiciones naturales. Para calcular la cantidad de materia seca de las raíces que se colocaron en las bolsas de descomposición, se secaron a 65°C hasta peso constante (± 0.001g), muestras de raíces de cada especie con el mismo peso fresco que se colocó en cada bolsa. En el periodo de alta precipitación la cantidad de materia seca colocada en las bolsas fue de 0.22, 0.24 y 0.16 g de raíces de cacao, laurel y poró, respectivamente. En el periodo seco, 0.25 g de materia seca de cada especie. El mismo día de recolectadas las muestras se lavaron con agua corriente sobre un tamiz de 0.5 mm de malla, se separaron por especie y se colocaron en el horno hasta peso constante. Posteriormente se pesaron y molieron para realizar análisis de tejidos. Para analizar las concentraciones de N, P, K, Ca y Mg, se hizo una muestra compuesta de las raíces de cada especie en cada tratamiento (Sistema), en cada bloque y en cada fecha de muestreo. En cada fecha de muestreo se calculó un factor de corrección para suelo adherido a las raíces (Muñoz y Beer, 2001). El contenido de N se determinó por el método de Kjehldal con digestión en ácido sulfúrico titulado con ácido bórico ; el contenido de P por colorimetría utilizando longitud de onda de 660 nm. El K, Ca y Mg se determinaron por espectrofotometría de absorción atómica en la solución de la digestión nitroperclórica (Díaz-Romeu y Hunter, 1982). Los resultados de biomasa y de nutrimentos se presentaron con base en el peso del material libre de cenizas (biomasa corregida * % de nutrimento = cantidad de nutrimento). Se utilizó el procedimiento NLIN de SAS para ajustar las medias del peso seco remanente de las raíces de cada especie y el contenido de nutrimentos en cada fecha, expresados como porcentaje del peso inicial de la muestra, al modelo de regresión exponencial doble (Wieder y Lang, 1982).
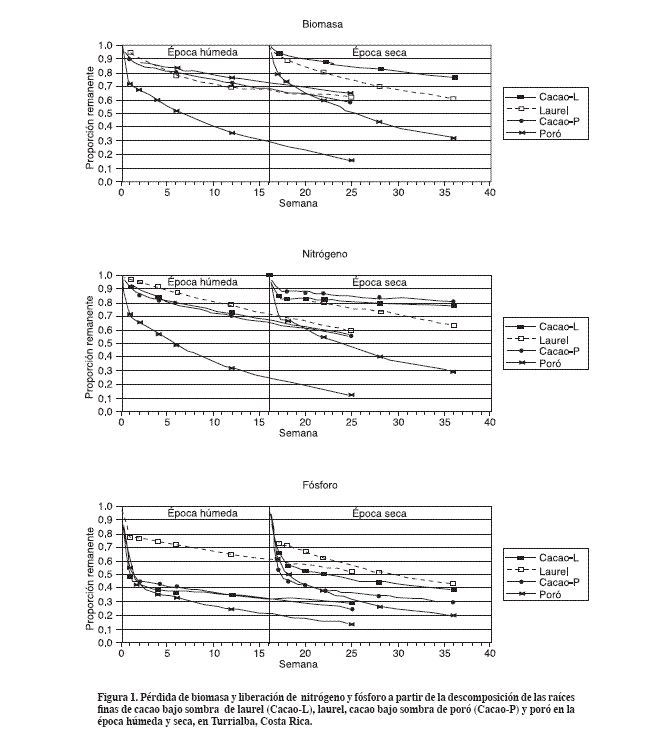
Resultados En general se observaron dos etapas en la descomposición de biomasa y liberación de nutrimentos: una rápida (1ª semana) y otra lenta (después de la segunda semana del periodo de incubación). Este comportamiento justifica el uso del modelo exponencial doble (Wieder y Lang, 1982) excepto en el caso del Ca, el cual por estar muy relacionado con el desarrollo de las poblaciones de hongos (Anderson y Swift, 1983), posiblemente se ajustaría más a un modelo de crecimiento poblacional. La no existencia de una temporada seca marcada en la zona hizo que los patrones de descomposición y liberación fueran relativamente similares en las dos épocas estudiadas. Para efectos de comparación entre los periodos seco y húmedo se tomó como punto final del estudio la semana 18 en los dos periodos, a pesar de que el último muestreo en el periodo húmedo se realizó a las 25 semanas de incubación. Biomasa En la época húmeda, el modelo exponencial doble explicó bien la pérdida de peso de las raíces finas causada por la descomposición (Figura 1). Al cabo de 18 semanas de descomposición en el periodo húmedo se observaron pérdidas de biomasa de 34% en las raíces de cacao bajo poró, 28% en cacao bajo laurel, 73% en poró y 34% en laurel. El modelo también explicó satisfactoriamente la pérdida de biomasa de los cuatro materiales durante las 18 semanas del periodo seco. El cacao en los dos sistemas perdió 23% de la biomasa, mientras que el poró y el laurel perdieron 68 y 39% respectivamente. La pérdida de biomasa fue menor que durante la época húmeda. Teniendo en cuenta que durante el periodo de estudio la humedad del suelo siempre fue mayor en el sistema cacao-poró (Muñoz y Beer, 2001), la pérdida de peso por descomposición en las raíces de cacao bajo poró sólo fue 6% mayor que la del cacao bajo laurel en la época húmeda y no se presentó diferencia con la época seca. El poró presentó la descomposición más rápida de las tres especies, en los dos periodos, y el cacao tendió a presentar descomposición más lenta que el laurel (Figura 1).
Nitrógeno El modelo exponencial doble explicó muy bien la liberación de N desde los cuatro materiales durante las épocas húmeda y seca del estudio (Figura 1). La mayor liberación de N se observó en la primera semana de las dos épocas, en las que se liberó el 29% del N inicial en poró, y el 6 y 15% en cacao en los periodos húmedo y seco, respectivamente, en cacao bajo laurel; el cacao bajo poró liberó el 10% de N en los dos periodos. El laurel liberó 3 y 15% en los periodos húmedo y seco respectivamente (Figura 1). A partir de la segunda semana, se observó una etapa de liberación más lenta. En esta etapa el aumento de humedad del suelo durante la época húmeda, parece haber acelerado la liberación de N desde todos los materiales excepto en el poró cuya liberación fue muy similar en las dos épocas (Figura 2).
Fósforo En la etapa de liberación rápida (1a semana), en los dos periodos, se observó la menor liberación de P desde las raíces de laurel, 22 y 27% en los periodos húmedo y seco, respectivamente. Durante esta etapa en el periodo húmedo, el cacao en los dos sistemas liberó 53% de P, mientras que en el periodo seco liberó 46% en el sistema cacao-poró y 34% en el sistema cacao-laurel. El poró presentó liberación de P similar en los periodos seco y húmedo (Figura 2). En la etapa de liberación lenta tendió a ser uniforme en todas las especies por lo que se mantuvieron las diferencias establecidas durante la primera semana de incubación (Figura 1). Potasio En las dos épocas la mayor tasa de liberación de K ocurrió en la primera semana, entre el 82 y 92% del contenido inicial de todas las especies y luego prácticamente se detuvo (Figura 2). Calcio El Ca presentó un comportamiento tendiente más hacia acumulación que a liberación, a pesar de esto el modelo exponencial doble se ajustó a los datos y describió el comportamiento de la liberación de Ca. En las raíces de laurel se observó acumulación de Ca hasta la cuarta semana en los periodos húmedo y seco, y en las de cacao bajo poró en la época seca. En el cacao bajo laurel en los dos periodos y en el cacao bajo laurel en el periodo húmedo se observó estabilidad en el contenido de Ca . El poró en los dos periodos mostró una tasa de liberación de Ca casi constante hasta el final del periodo de estudio (Figura 2). Magnesio En el laurel y cacao en los dos sistemas y durante los dos periodos, la etapa de liberación rápida del Mg se extendió hasta la cuarta semana, observándose tendencia a mayor liberación en la época húmeda, mientras que en el poró la etapa de liberación rápida en los dos periodos se registró en la primera semana. Durante la etapa de liberación rápida en la época seca, el cacao en los dos sistemas liberó 42% y el laurel 35%. En la época húmeda, el cacao bajo laurel y el laurel liberaron casi 54% de Mg y el cacao bajo poró 59% (Figura 2). El poró, en la etapa de liberación rápida, en los dos periodos, liberó 64% en la primera semana. Posteriormente, durante la etapa de liberación lenta, la tasa tendió a hacerse similar (Figura 2). En el trabajo se estudió la liberación de los nutrimentos desde las raíces en descomposición pero no se estudiaron los procesos de fijación y mineralización que son de vital importancia para conocer el balance, disponibilidad y sostenibilidad en el uso de los nutrimentos de un sistema. También se debe tener en cuenta que si se presentara relocalización de nutrimentos antes de la muerte de las raíces finas, en este estudio podría presentarse sobreestimacion ya que se usaron raíces vivas recién cortadas. Aunque en estudios realizados en pinos en Australia se encontró poca evidencia de que esto pueda ocurrir (Nambiar, 1987). Otro aspecto a tener en cuenta es el que al colectar las raíces que se usan en los estudios con bolsas de descomposición se disturban las asociaciones de organismos en la rizosfera lo que puede producir subestimación de las tasas de descomposición (Dornbush et al., 2002). A pesar de los inconvenientes citados este estudio provee una estimación de las tasas relativas de liberación de nutrimentos a partir de la descomposición de las raíces finas de las especies estudiadas. En estudios anteriores de descomposición de raíces y liberación de nutrimentos se ha reportado un patrón de liberación con una etapa rápida al inicio y lenta al final del proceso, pero generalmente en estos estudios de larga duración, 2 años o más, los intervalos de muestreo tambien son largos (2 meses o más) (King et al., 1997), en este estudio los intervalos de muestreo fueron cortos al inicio y se ampliaron hacia el final, esta es una ventaja ya que se pudo describir bien el inicio del proceso donde se encuentra la parte más dinámica y tal vez más importante debido a la relativamente gran liberación de todos los nutrimentos, excepto Ca. La relativamente rápida descomposicion de la biomasa y la liberación de nutrimentos en la etapa inicial del estudio, seguidas por otra época de descomposición y liberación lenta, podrían explicarse biológicamente como el resultado de la liberación rápida del citoplasma y las estructuras contenidas en él, seguidas por una descomposición lenta de las membranas, paredes celulares y el contenido que existe entre ellas. La liberación de N, P y Mg siguió el patrón de liberación inicial rápida y posteriormente lenta a partir de la segunda semana (Figura 1 y Figura 2). El 84% del K fue liberado en la primera semana del periodo de incubación en todas las especies estudiadas, demostrando alta movilidad y escasa incorporación en estructuras celulares. En estudios con raíces de pino en la zona templada (King et al., 1997) y árboles leguminosos en el trópico (Lehmann et al., 1995) también se ha hecho la misma observación. La liberación rápida y la lixiviación del K no parece ser un problema en el sitio ya que es insignificante al compararla con la cantidad de K que entra al sistema por fertilización, precipitación y las reservas del suelo (Imbach et al., 1989). El Ca presentó tendencia a la acumulación excepto en las raíces de poró (Figura 2), sugiriendo que este material en descomposición no es un sustrato estable para la formación de colonias de hongos los cuales son los responsables de movilizar Ca hacia los materiales en descomposición (Anderson y Swift, 1983). En estudios de lixiviación de nutrimentos en este mismo sitio se reportaron datos de 27 y 7 kg ha-1 año de Ca lixiviado en los sistemas cacao/poró y cacao/laurel respectivamente (Imbach et al., 1989), concordando con la rápida liberación de Ca observada en este estudio desde las raíces de poró (Figura 2). También se reportó tendencia a la estabilidad del Ca en la hojarasca de cacao y laurel y rápida liberación desde la hojarasca de poró (Vilas Boas, 1990). En un estudio de lixiviación en el mismo sitio se reportaron 20 y 6 kg ha-1 año de Mg lixiviado en los sistemas cacao/poró y cacao/laurel respectivamente y se estimó que si se dejara de aplicar fertilizante al sistema cacao/poró el Mg se agotaría en 30 años (Imbach et al., 1989). A pesar de que en este sitio estas pérdidas no presentan problema al compararlas con las entradas de Mg al sistema, este hecho se debe tener en cuenta cuando se incorporen especies de rápida descomposición como Erythrina poeppigiana en sistemas agroforestales de otras regiones donde las pérdidas por lixiviación podrían ser un problema. El rápido flujo de nutrimentos desde las raíces de poró al iniciarse la descomposición podría ser contraproducente en sitios donde haya posibilidad de pérdidas de nutrimentos por escorrentía y/o lixiviación. El estudio de las tasas de mineralización de los nutrimentos liberados desde las raíces finas en descomposición y el conocimiento del momento de la ocurrencia de la época de máxima absorción, posiblemente el mes de junio en este sitio (Muñoz y Beer, 2001), posibilitarían ajustar la fecha de la poda del poró para que los nutrimentos liberados por los residuos y la posible biomasa de raíces finas muertas estuvieran disponibles para el cacao y de esta forma afinar las actividades de manejo del cultivo. La relativamente lenta descomposicion de las raíces de laurel apoya el reporte de que la biomasa de raíces finas del sistema cacao/laurel fue mayor que la del sistema cacao/poró (Muñoz y Beer, 2001) .
El autor agradece al CATIE y en especial al doctor John Beer por su apoyo y guía para lograr el título de Magister Scientiae. El apoyo financiero a este proyecto fue dado por GTZ, CATIE, y el gobierno holandés como oferente de la beca de estudio del autor. Anderson, J. M.; Swift, M. J. 1983. Decomposition in tropical forest. En: Sutton S. L.; Whitmore, T. C. ; y Chadwic, A. C. (eds.) Tropical Rain Forest: ecology and management. Oxford. Blackwell. pp. 287-309 Díaz-Romeu, R. 1982. Metodología de muestreo de suelos, análisis químico de suelos y tejido vegetal y de investigaciones en invernadero. CATIE. Serie de materiales de enseñanza N° 12. 61p. (Mimeografiado). Dornbush, M. E.; Isenhart, T.M.; Raich, J.W. 2002. Quantifying fine-root decomposition: an alternative to buried litterbags. Ecology, Vol 83:2985-2990. Fabiao, A. ; Persson, H. A. ; Steen, E. 1985. Growth dynamics of superficial roots in portuguese plantations of Eucaliptus globolus Labill, studied with a mesh bag Technique. Plant Soil 83 :233-242 Fassbender, H. W. 1993. Modelos edafológicos de sistemas agroforestales. 2 ed. CATIE (Serie de materiales de enseñanza N° 29). 491p. Imbach, A.C. ; Fassbender, H.W. ; Borel, R. ; Beer, J. ; Bonnemann, A. 1989. Modeling agroforestry systems of cacao (Theobroma cacao) with laurel (Cordia alliodora) and poró (Erythrina poppigiana) in Costa Rica. IV. Water balances, nutrient inputs and leaching. Agrofor Syst 6:37-48. King, J. S.; Allen, L.H.; Dougherty, P. y Strain, B.R. 1997. Decomposition of roots in loblolly pine: Effects of nutrient and water availability and root size class on mass loss and nutrient dynamics. Plant Soil 195 1:171-184. Lehmann, J.; Schroth, G.; Zech, W. 1995. Decomposition and nutrient release from leaves, twigs and roots of three alley-cropped tree legumes in central Togo. Agrofor Syst 29:21–36. Muñoz, F.; Beer, J. 2001. Fine root dynamics of shaded cacao plantations in Costa Rica. Agrofor Syst 51: 119-130. Nambiar, E.K.S. 1987. Do nutrients retranslocate from fine roots? Can J For Res 17:913-918. Noorwijk, M. Van. 1992. Roots: length, biomass, production and mortality. En: Anderson, J.M.; Ingram, J.S.I. (ed) TSBF Methods handbook. Appendix D. pp. 127-138. Persson, H. 1979. Fine-root production, mortality and decomposition in forest Ecosystems. Vegetatio 41(2): 101-109 Steen, E. 1985. Root and rhizome dynamics in a perennial grass crop during an annual growth cycle. Swed J Agric Res 14: 93-97. Vilas Boas, O. 1990. Descomposición de hojarasca y mineralización del nitrógeno de la materia orgánica del suelo bajo cuatro sistemas agroforestales, en Turrialba, Costa Rica. Tesis M. Sc. CATIE. 152 p. Wieder, R.K.; Lang, G.E. 1982. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 63: 1636–1642.
|


Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2006 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.






















