



Publicado
Correlaciones genotípicas, fenotípicas y ambientales, y análisis de sendero en tomate de árbol (Cyphomandra betacea Cav. Sendt.)
Genotypic correlations, phenotypic, environmental and path analysis in tomato tree (Cyphomandra betacea Cav. Sendt.)
Palabras clave:
Análisis de sendero, correlación ambiental, correlación fenotípica, correlación genética, Cyphomandra betacea (es)Cyphomandra betacea, environmental correlation, genetic correlation, phenotypic correlation, path analysis (en)
Descargas
1I.A; M.Sc en Ciencias Agrarias. Línea Fitomejoramiento. Universidad Nacional de Colombia. Palmira, Valle del Cauca, Colombia.
2I.A.; M.Sc.; Ph.D; Genética y Mejoramiento de Plantas. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia.
Palmira, Valle del Cauca, Colombia, A.A 237. 3I.A.; M.Sc.; Ph.D. Facultad de Ciencias Agrícolas. Universidad de Nariño. Pasto,
Nariño, Colombia. 4I.A. Universidad de Nariño. Pasto, Nariño, Colombia.
*Autor para correspondencia: lklagoss@unal.edu.co.
En el estudio se evaluaron diferentes componentes de producción y se estimaron las correlaciones fenotípicas, genéticas y ambientales, considerando nueve variables relacionadas con el tamaño y la calidad del fruto de tomate de árbol (Cyphomandra betacea Cav. Sendt.); igualmente se establecieron los efectos directos e indirectos de las variables componentes de calidad sobre el peso del fruto. Para el efecto se utilizaron los datos existentes de 81 híbridos (72 híbridos interpoblacionales y nueve testigos) dispuestos en tres repeticiones, en condiciones del departamento de Nariño, Colombia. Los resultados obtenidos indicaron que las correlaciones genotípicas fueron superiores a las fenotípicas y ambientales. El peso de fruto (PF) presentó las mayores correlaciones genéticas con el peso de semilla por fruto (rG = 0.97) y con el peso de pulpa más semilla (rG = 0.92). El análisis de sendero con base en correlaciones genotípicas mostró que el peso de pulpa más semilla fue la variable que tuvo el mayor efecto directo sobre el PF (1.353). Esto demuestra que una selección por peso de fruto da como resultado un aumento en peso de pulpa más semilla. Teniendo en cuenta las correlaciones fenotípicas, este análisis permitió establecer que los efectos directos de peso de semilla de fruto (PSF) y peso de pulpa más semilla (P + S) (1.166 y 0.743, respectivamente) son los que más contribuyen al PF.
Palabras clave: Análisis de sendero, correlación ambiental, correlación fenotípica, correlación genética, Cyphomandra betacea.
The study different production components were evaluated and phenotypic, genetic and environmental correlations were estimated, considering nine characters related to the size and quality of tomato tree fruit (Cyphomandra betacea Cav Sendt.), also settled the direct and indirect effects of the component variables of quality on fruit weight. To effect the existing data of 81 hybrids (72 interpopulation hybrids and nine witnesses) arranged in three replications, under the Department of Nariño, Colombia were used. The results indicated that genotypic correlations were higher than phenotypic and environmental. The fruit weight (PF) presented the highest genetic correlations with seed weight per fruit (rG = 0.97) and with more seed pulp weight ( rG = 0.92 ) . Path analysis based on genotypic correlations showed that more seed pulp weight was the variable that had the most direct effect on the PF (1.353). This shows that a selection of the fruit weight results in an increase in weight of pulp more seed. Given the phenotypic correlations, this analysis established that the direct effects of seed weight of fruit (PSF) and more seed pulp weight (P + S ) (1.166 and 0.743 , respectively ) are the largest contributors to PF.
Key words: Cyphomandra betacea, environmental correlation, genetic correlation, phenotypic correlation, path analysis.
En Colombia, el tomate de árbol (Cyphomandra betacea Cav. Sendt.) es una de las frutas con mayor potencial para el mercado debido a sus características nutricionales y organolépticas, además es una alternativa económicamente viable para los agricultores de la región Andina de Colombia. Según el Plan F rutícola N acional, l a s uperficie q ue ocupa actualmente este cultivo en el país es de 9.223 ha, de las cuales el departamento de Nariño participa con el 9.08% (760 ha) (Agronet, 2011). Para 2014 se proyecta en Colombia un incremento en la superficie sembrada con este cultivo de aproximadamente 10.000 ha, para generar 7.823 empleos directos (Tafur, 2006). Su potencial frutícola está determinado por su adaptación a condiciones tropicales; no obstante este gran potencial se ve afectado por la condición semisilvestre del cultivo y por la falta de soporte tecnológico, ya que su desarrollo se basa fundamentalmente sobre el conocimiento empírico, como resultado del esfuerzo de los agricultores. Esto se refleja en la falta de una oferta de genotipos mejorados que suplan las necesidades del productor y ayuden a resolver los problemas fitosanitarios del cultivo en la zona andina del departamento de Nariño (Lagos, 2008).
En Colombia y Ecuador, los principales países productores de la fruta, no existen variedades comerciales, sino poblaciones locales que tradicionalmente se han venido multiplicando y seleccionando por parte de los cultivadores. En consecuencia, este cultivo se caracteriza por la gran heterogeneidad en las formas y tamaños de las frutas entre y dentro de una misma plantación, resultado de las hibridaciones y de las mezclas del material genético a través del tiempo (Lobo, 2000).
Esta especie aún se encuentra en proceso de domesticación y los cultivares existentes han sido producto de la selección empírica por parte de los agricultores. Las poblaciones de estos cultivares exhiben un alto grado de vulnerabilidad a problemas sanitarios debido a la co-evolución natural de las poblaciones con los patógenos, al bajo nivel de selección al que ha sido sometida la especie y a que las semillas que siembran los agricultores tienen un origen desconocido (Lagos, 2008).
El trabajo del fitomejorador para identificar los individuos o cultivares que reúnan simultáneamente características deseables no es fácil, ya que muchos de estos caracteres se encuentran asociados positiva o negativamente. Las asociaciones entre los caracteres de interés en fitomejoramiento se evalúan por medio de correlaciones fenotípicas, genotípicas y ambientales. La correlación fenotípica es estimada directamente de los valores medios fenotípicos de campo, siendo, por tanto, el resultado de causas genéticas y ambientales. La correlación genotípica, en cambio, corresponde a la porción genética de la correlación fenotípica y es empleada para orientar programas de mejoramiento por ser la única de naturaleza heredable (Cruz, 2001; Cruz y Regazzi, 1997; Falconer y Makay, 1996; Vencovsky y Barriga, 1992; Mariotti, 1986; Hallauer y Miranda, 1981). Sin embargo, los coeficientes de correlación, a pesar de ser de gran utilidad en la cuantificación de la magnitud y dirección de las influencias de factores en la determinación de caracteres complejos, ofrecen sólo una importancia relativa de los efectos directos e indirectos de esos factores. La solución que se plantea para esta limitación es realizar un análisis de sendero, ya que por medio de él se desdoblan las correlaciones estimadas en efectos directos e indirectos (Falconer y Mackay, 1996; Cruz y Regazzi, 1997).
De acuerdo con lo anterior, en esta investigación se buscó identificar las variables que más influyen en la calidad del fruto y el rendimiento del tomate de árbol, para ser utilizadas como criterios de selección en programas de mejoramiento de esta especie, con el objeto de satisfacer las demandas de los productores y del sector agroindustrial. Mediante las variables peso de fruto (PF), peso de pulpa más semillas (P + S), contenido de jugo (CJ), pH, grados brix (BX), acidez titulable (AC), índice de madurez (IM), peso de semilla de fruto (PSF) y número total de semilla (NTS) se conceptualizaron las correlaciones fenotípicas, genotípicas y ambientales que fueron descompuestas mediante un análisis de sendero.
Localización
El trabajo se realizó en tres sitios considerados como repeticiones para efectos del diseño experimental. El primero, en el municipio de Pasto, corregimiento de la Pradera, a 01° 19' 33.3" N y 77° 19' 18.9" O, con una temperatura promedio de 18 °C y 1980 m.s.n.m.; el segundo en el municipio de Ipiales, vereda Yanalá, a 0° 52' 23" N y 77° 33' 3.8" O, con una temperatura promedio de 12 °C y 2730 m.s.n.m.; y el tercero en el municipio de Tangua, vereda El Placer, a 1° 8' 10.08" N y 77° 26' 3.41" O, con una temperatura promedio de 16 °C y 2000 m.s.n.m.
Diseño experimental
El diseño experimental utilizado en esta investigación fue el de bloques completos al azar, con tres repeticiones (sitios de recolección de frutos) y 81 tratamientos que correspondieron a 72 híbridos interpoblacionales y nueve testigos (Cuadro 1). La parcela experimental consistió en seis plantas, a una distancia de 2.5 m entre ellas, la distancia entre parcelas experimentales fue de 2.5 m y la parcela útil de cuatro plantas centrales que ocupaban 25 m2.
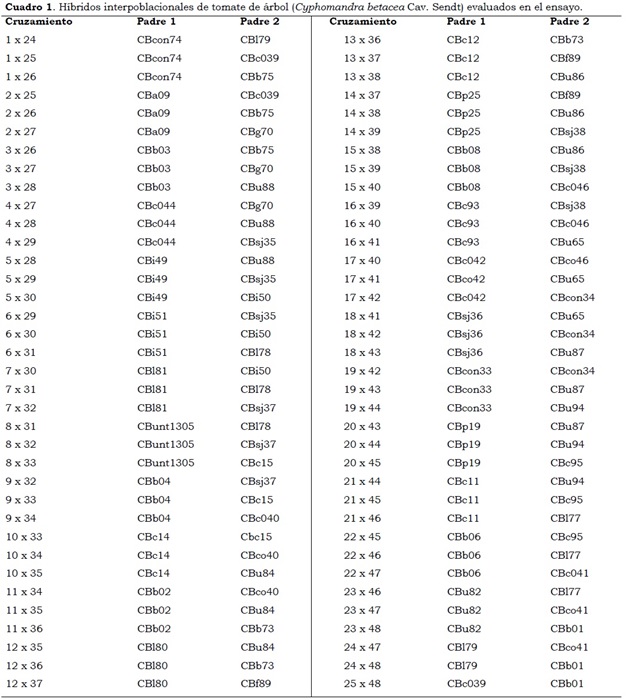
Variables evaluadas
Los frutos, una vez cosechados, fueron llevados al Laboratorio de Biotecnología de la Universidad de Nariño para los análisis respectivos, como se detalla a continuación.
Peso del fruto (PF). Se midió en seis frutos por parcela experimental.
Peso de pulpa más semilla (P + S). Para esta medición se tomaron los pesos (g) de la pulpa incluyendo las semillas de cada uno de los frutos por parcela experimental.
Contenido de jugo (CJ). Utilizando una licuadora se extrajo el jugo de la pulpa de cada uno de los frutos y se midió su volumen en ml.
pH. Se obtuvo empleando un potenciómetro Inolab-WTW series pH 720".
Sólidos solubles totales (°brix). Se determinaron con refractómetro Atago de bolsillo PAL-1. La lectura se corrigió utilizando el porcentaje de ácido cítrico (A.C.), mediante la ecuación: bx = 0.19 x A.C. + S.S.T., donde S.S.T. son los sólidos solubles totales.
Acidez titulable (AT). Se determinó por el método de titulación potenciométrica y se expresó como porcentaje de A.C; para el cálculo se utilizó la ecuación: %A.T. = (V1 x N) / V2) x K x 100, donde: V1 = volumen de NaOH utilizado (ml); V2 = volumen de la muestra (5 ml); K = peso equivalente del ácido cítrico (64,04 g) y N = normalidad del NaOH (0.1 meq/ml).
Índice de madurez (IM). Se calculó mediante la relación entre el contenido de sólidos solubles y la acidez total (Galvis, 1992), mediante la ecuación: I.M. = (S.S.T.) / (A.T.).
Peso de semillas en fruto (PSF). Se obtuvo por extracción de las semillas después de pasar el jugo a través de un colador y posterior lavado, limpieza y secado al ambiente durante 7 días.
Número total de semillas por fruto (NTS). Es la relación entre PSF y el peso de 100 semillas correspondientes a cada genotipo.
Análisis estadístico
Las variables antes mencionadas fueron sometidas a análisis de correlación fenotípica, genotípica y ambiental, de acuerdo con lo propuesto por Ceballos (1997). Para el PF se realizaron dos análisis de sendero (Singh y Chaudhary, 1985), uno con base en las correlaciones fenotípicas y otro teniendo en cuenta las correlaciones genotípicas. También se hicieron grupos de las variables altamente correlacionadas (r > 0.60), que se utilizaron para seleccionar aquellas de mayor importancia.
Los coeficientes de correlación fenotípicos, genéticos, ambientales y de sendero se estimaron usando el programa GENES desarrollado por Cruz (2006). El programa aplica las fórmulas clásicas de correlación:
Correlación genética (rG(XY)): rG(XY) = COVG(XY)/SG(X).SG(Y)
Correlación ambiental (rE(XY)): rE(XY) = COVE(XY)/SE(X).SE(Y)
donde, r(XY) y COV(XY) = son correlaciones y covarianzas fenotípicas (rF), genéticas (rG) y ambientales (rE) entre los caracteres X e Y, respectivamente; S(x) y S(y) = son desviaciones estándar fenotípicas, genéticas y ambientales de X e Y, en su orden.
Una vez estimados los coeficientes de correlación se confirmó la significancia estadística para cada uno de ellos (r), planteando la hipótesis nula: Ho: r = 0 vs. la hipótesis alterna Ha: r ¹ 0, mediante una prueba de ‘t’, dada por tc = rx(n – 2)1/2/(1 – r2) 1/2. La ‘t’ calculada (tc) se comparó con una ‘t’ de la Tabla (tt), al nivel de significancia de 0.05 y con n = 2 grados de libertad. La regla de decisión fue: sí tc > tt, entonces el valor de r es estadísticamente diferente de cero (Espitia et al., 2008).
El análisis de sendero consiste en desdoblar el coeficiente de correlación (fenotípico, genético o ambiental) en los efectos directos e indirectos de varios caracteres (causas) sobre una variable básica compleja (efecto). Con base en las matrices de correlaciones fenotípicas y genéticas se desdoblaron los rF y los rG para determinar los efectos que influyen sobre el PF. En este caso, el sistema PF (variable efecto) estuvo en función del PUS, CJ, pH, BX, AC, IM, PSF y NTS (variables causas).
Los coeficientes de correlación fenotípica (rF), genética (rG) y ambiental (rE) encontrados aparecen en el Cuadro 2. Sin excepciones, las rG fueron de mayor magnitud que las rF. Los resultados obtenidos indican una rG positiva y significativa (P < 0.05) entre las variables PF y P + S (0.92), PF y CJ (0.81) y entre PF y PSF (0.97) y evidencian una acción genética común entre estas variables, lo cual facilita la selección ya que el proceso se haría utilizando cualquiera de ellas. Las rF y rG del PF con P + S (0.71 y 0.97, respectivamente) y con CJ (0.51 y 0.81, en su orden) fueron significativas (P < 0.05). Las magnitudes de la correlación fenotípica genética positivas indican que la selección por PF produce un incremento en el carácter respectivo antes citado (Cuadro 2).
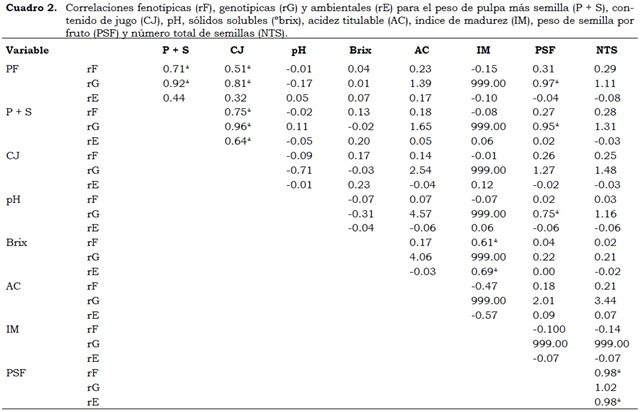
Las rF, rG y rE, entre las variables P + S y CJ muestran correlación fenotípica menor que la correlación genética (rF = 0.75 < rG = 0.96), lo que implica una correlación ambiental importante (rE = 0.64) y por tanto existe un efecto de los factores ambientales y/o de los factores no aditivos que afectan negativamente el nivel de asociación real entre ambos caracteres de estudio (Espitia et al., 2008). Lo anterior también se presenta entre las variables PSF y NTS (rF = 0.98 < rG = 1.02, rE = 0.98).
En la característica SST (°brix) hay una correlación genética y ambiental positiva y significativa con el IM, pero no existe una relación genética entre ellos, por tanto, el fenotipo es debido al efecto del ambiente y no al genotipo.
Las correlaciones genéticas positivas y significativas entre PSF con P + S (0.95) y pH con PSF (0.75) pueden también ser consideradas de importancia, ya que sugieren que una selección por PSF incide de manera directa en el aumento o reducción del P + S y el pH. Se puede asumir que las correlaciones genéticas > 1 (Cuadro 2) son perfectas, o que se recomendaría revisar las varianzas de cada una de las variables involucradas en el análisis de correlación, si una de ellas presenta diferencias o una varianza significativa y la otra no, caso en el cual esta correlación debería ser desechada (Checa, 2012) ya que la correlación mide el grado de covarianza entre dos variables (Mayo, 1980).
Los análisis de sendero, que muestran la descomposición de las correlaciones fenotípicas (rF) y genéticas (rG) para el PF aparecen en los Cuadro 3 y Cuadro 4. En el análisis de sendero para las rF (Cuadro 3) se observa que los efectos directos de P + S (0.743) y PSF (1.166) sobre el coeficiente de correlación (PF: 0.707 y 0.313, respectivamente) son positivos y mayores que los efectos indirectos de las demás variables incluidas en el análisis. Estos resultados, tanto de los efectos directos como el coeficiente de correlación explican la verdadera relación existente entre ambos caracteres, por tanto, una selección directa a través de esta característica resultaría efectiva (Singh y Chaudhary, 1985).
El coeficiente R2 del análisis de sendero con base en las correlaciones fenotípicas fue de 56% (Cuadro 3), considerado bajo por lo que se infiere que las variables presentes en el modelo no explican fenotípicamente el PF. En el análisis las variables con mayor efecto directo sobre el PF fueron P + S (0.743) y PSF (1.166), mientras que en el genético (Cuadro 4) fueron P + S (1.35) y PSF (0.874).
El coeficiente de determinación (R2) en el análisis de sendero para rG indica que 100% de la variabilidad del PF fue explicada por las variables P + S, CJ, PH, BX, AC, IM, PSF y NTS, lo que es un buen ajuste del modelo y muestra la importancia de las variables explicativas en la definición del PF (Espitia et al., 2008).

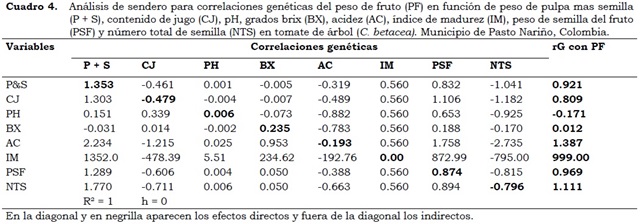
La descomposición de la correlación (rF = 0.512) entre CJ y PF (Cuadro 2) se explica en mayor proporción por los efectos indirectos de P + S (0.554) que por el efecto directo de la variable CJ (-0.077); esto indica que la correlación significativa y directa existente entre CJ y PF es debida, en mayor proporción, a la influencia indirecta a través de P + S. Con base en el primer análisis de sendero (Cuadro 3) se puede inferir que la selección por mayor P + S y CJ permite la obtención de frutos más pesados.
En el caso del análisis de sendero para las rG (Cuadro 4) se observa que el efecto directo de P + S (1.353) sobre el coeficiente de correlación PF = 0.921 es mayor que los efectos indirectos de las otras variables que se incluyeron en el análisis. Al ser positivos, tanto el efecto directo como el coeficiente de correlación, este último explica la verdadera relación existente entre ambos caracteres, e indica que la selección directa a través de esta característica es efectiva (Singh y Chaudhary, 1985). En el mismo Cuadro 4 se observa que en el análisis de sendero para correlaciones genéticas, el efecto directo de la variable PSF sobre el PF (0.874) es menor que el efecto indirecto de P + S sobre PF (1.289); en este caso, el valor de la correlación (0.969) se le atribuye al efecto indirecto de la variable P + S. En esta situación el factor causal indirecto es considerado para los procesos de selección (Singh y Chaudhary, 1985).
- En este estudio, los altos valores de correlación genética mostraron que las variables que más influyen en el peso del fruto de tomate de árbol y que pueden ser utilizadas como criterios de selección son peso de pulpa más semilla por fruto y peso de semilla por fruto.
- En el análisis de sendero para correlaciones fenotípicas y genéticas, las variables que mayor efecto directo tuvieron en el peso del fruto fueron el peso de semilla por fruto y el peso de pulpa más semilla por fruto.
Agronet. 2011. Producción de tomate de árbol en el departamento de Nariño. Ministerio de Agricultura y Desarrollo Rural (MADR). 3 p.
Ceballos, H. 1997. Genética cuantitativa y fitomejoramiento. Palmira, Universidad Nacional de Colombia sede Palmira. 330 p.
Cruz, C.; y Regazzi, C. 1997. Modelos biométricos aplicados aomelhoramento genético. 2a ed. Ediciones Universidade Federal de Vicosa. Vicosa, MG, Brasil. 390 p.
Cruz, C. 2001. Programa GENES. Versao Windows. Aplicativo computacional em genética e estatística. Ediciones Universidade Federal de Vicosa. Vicosa, MG , Brasil. 648 p.
Cruz, C. 2006. Programa GENES. Versao Windows. Aplicativo Computacional em Genética e Estatística. Editora UFV. Universidade Federal de Viçosa. Disponible desde Internet en: www.ufv.br/dbg/genes/genes.htm. Revisado: Julio de 2012.
Checa, C. 2012. Comunicación personal. Pasto, Universidad de Nariño, Facultad de Ciencias Agrícolas.
Espitia, M.; Aramendiz, H.; y Cadena, J. 2008. Correlaciones y análisis de sendero en algodón Gossypium hirsutum L. en el Caribe colombiano. Rev. Fac. Nac. Agron. 61(1):4325 - 4335.
Falconer, D. y Mackay, T. 1996. Introduction to quantitative genetics.4th edition.Prentice Hall, New Jersey, EE. UU. 464 p.
Galvis, A. 1992. Tecnología de manejo de post-cosecha de frutas y hortalizas: Sección de Vegetales. Instituto de Ciencia y Tecnología de Alimentos (ICTA), Universidad Nacional de Colombia. Bogota.
Hallauer, A. y Miranda, J. 1981. Quantitative genetics in maize breeding. lowa State University Press, Ames, IA. 468 p.
Lagos, T. C. 2008. Proyecto Obtención y Evaluación Preliminar de Híbridos de Tomate de Árbol Cyphomandra betacea Cav. en la zona Andina de Nariño. Ministerio de Agricultura y Desarrollo Rural (MADR).
Lobo, M. 2000. Papel de la variabilidad genética en el desarrollo de los frutales andinos como alternativa productiva. En: Memorias 3° Seminario de Frutales de Clima Frío Moderado. Centro de Desarrollo Tecnológico de Frutales, Manizales, Colombia, 15-17 de noviembre de 2000. p. 27 - 36.
Mariotti, J. 1986. Fundamentos de genética biométrica. Aplicaciones al mejoramiento genético vegetal. Secretaría General de la Organización de los Estados Americanos (OEA), Washington, D. C. 152 p.
Mayo, O. 1980. The theory of plant breeding.Oxford University. Clarendon Press. 293 p.
Singh, R. y Chaudhary, D. 1985. Biometrical methods in quantitative genetic analysis. Path analysis. Nueva Delhi, Ludhiana. p. 78.
Tafur, R. 2006. Propuesta frutícola para Colombia y su impacto en la actividad económica, nacional, regional y departamental. En: Sociedad Colombiana de Ciencias Hortícolas. Memorias Primer Congreso Colombiano de Horticultura. 240 p.
Vencovsky, R. y Barriga, P. 1992. Genética biométrica no fitomelhoramento. Sociedad Brasileira de Genética, Brasil. 496 p.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2013 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.






















