



Publicado
Caracterización de cultivares de duraznero [Prunus persica (L.) Batsch.] por resistencia a heladas
Characterization of peach [Prunus persica (L.) Batsch.] cultivars by frost resistance
DOI:
https://doi.org/10.15446/acag.v64n3.47565Palabras clave:
Aclimatación, congelación, densidad de floración, fenología, TL50, Prunus persica (es)Acclimation, freezing, bloom density, phenology, LT50, Prunus persica (en)
Descargas
Caracterización de cultivares de duraznero [Prunus persica (L.) Batsch.] por resistencia a heladas
Characterization of peach [Prunus persica (L.) Batsch.] cultivars by frost resistance
Javier Emilio Chaar
Instituto Nacional de Tecnología Agropecuaria, Estación Experimental Agropecuaria Junín, Isidoro Busquets, Junín, Mendoza, Argentina. Autor para correspondencia: chaar.javier@inta.gob.ar
Rec.: 28.11.2014 Acep.:06.01.2015
Resumen
Las heladas primaverales son una de las principales limitantes de la producción de frutales de clima templado. Dentro de una misma especie existe variabilidad en resistencia frente al daño en órganos florales ocasionado por temperaturas bajo cero durante la salida del reposo invernal. En cinco cultivares de duraznero [Prunus persica (L.) Batsch.] y uno de nectarino se evaluó el daño ocasionado por heladas y se determinaron la fecha de plena floración y la densidad de floración. Adicionalmente se determinó la temperatura letal media (TL50) de las yemas florales en el estado de flor abierta, mediante descensos térmicos controlados en laboratorio. Los cultivares (cv) de duraznero Maria Bianca y Summer Pearl presentaron las mayores densidades de flores sanas por cm de ramo, luego de la ocurrencia de temperaturas bajo cero en campo. La resistencia a heladas en campo se relacionó principalmente con la elevada densidad de floración, en combinación, en algunos casos, con floración tardía. La floración tardía por sí sola no resultó una característica de resistencia; por tanto, para la elección de cultivares de duraznero con menor riesgo de daño por temperaturas bajo cero es importante tener en cuenta más de una variable relacionada con los órganos reproductivos.
Palabras claves: Aclimatación, congelación, densidad de floración, fenología, TL50, Prunus persica.
Abstract
Spring frosts are one of main crop constraints of temperate fruit trees. Within a species exists variability in damage caused by subzero temperatures on flower organs during breaking winter rest. In five peach [Prunus persica (L.) Batsch.] and one nectarine cultivars, field frost damage was assessed, also recording full bloom date and bloom density. Additionally, mean lethal temperature (LT50) of flower buds in the open flower state was determined through laboratory controlled thermal declines. Peach cultivars Maria Bianca and Summer Pearl had higher density of healthy flowers per cm of shoot, after field subzero temperatures. Field frost resistance was related mainly with high bloom density, combined, in some cases, with late bloom. Late bloom did not result a resistance feature itself. Therefore, to select peach cultivars with less damage risk by subzero temperatures, it is important to consider more than one variable related with reproductive organs.
Key words: Acclimation, freezing, bloom density, phenology, LT50, Prunus persica.
Introducción
La distribución y la productividad de las especies vegetales están limitadas, entre otros factores, por los extremos de temperatura (Fiorino y Mancuso, 2000). El comportamiento exitoso de una especie leñosa en un sitio implica la sincronización del desarrollo anual de la resistencia a bajas temperaturas con los cambios estacionales de éstas (Lindén, 2002). Las heladas primaverales son una de las principales limitantes de la producción de frutales de clima templado, no obstante algunas especies han evolucionado y desarrollado mecanismos de resistencia, siendo ésta una de las características más importantes para la selección de cultivos, que puede determinar la factibilidad de obtener cosechas aceptables, minimizando el aporte de energía calórica (Chaar, 2013). Quamme et al. (1982) en especies cultivadas y nativas del género Prunus encontraron que la temperatura a la cual ocurre daño en los tejidos más susceptibles está estrechamente relacionada con el promedio de la temperatura mínima anual en el límite de distribución geográfica de los materiales estudiados. Kadir y Proebsting (1994) encontraron que las yemas florales de 20 especies de Prunus mostraban diferentes estrategias para hacer frente a bajas temperaturas. Aún dentro de una misma especie existe variabilidad frente al daño ocasionado por temperaturas bajo cero en los órganos florales durante la salida del reposo invernal (Kodad y Socias i Company, 2005). La temperatura letal media (TL50) es aquella temperatura bajo cero a la cual muere el 50% de las yemas florales (Mathers, 2004). La resistencia al frío de las estructuras reproductivas depende, entre otros factores, de la fecha de plena floración, la TL50 y la densidad floral (Chaar, 2013). Es posible evaluar la resistencia relativa al frío de distintos materiales vegetales ante la ocurrencia de heladas en campo (Granger y Rousselle, 1984; Lisek, 2007); por lo tanto el objetivo de este trabajo fue evaluar la magnitud del daño en yemas florales y cuantificar las variables de resistencia de distintos cultivares de duraznero y nectarino después de la ocurrencia de heladas en campo durante la salida del reposo invernal de 2013 en Mendoza, Argentina.
Materiales y métodos
Localización y material experimental
Los ensayos se realizaron en el Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria Junín, provincia de Mendoza (Argentina) a 33° 6’ 57.5” sur, 68° 29’ 4” oeste, a 653 m.s.n.m., en una colección de cultivares (cv) de duraznero para consumo en fresco, incluyendo los cvs Flavorcrest, Milenio INTA, Maria Bianca, Summer Pearl y Compact Red Haven, y el nectarino cv Zee Gold. De cada cultivar fueron evaluadas tres plantas adultas, cultivadas en semipalmeta con orientaciones norte–sur y regadas por gravedad.
Determinación de la TL50
A la salida del reposo otoño–invernal de 2012, en laboratorio se determinó la TL50 mediante descensos térmicos en un ‘freezer’, simulando la ocurrencia de heladas en campo. Debido a que la tolerancia a heladas depende del estado de desarrollo de las yemas, para comparar los resultados los cultivares fueron evaluados en un mismo estado fenológico (Dale, 1987; Kodad et al., 2010). De cada cultivar evaluado se utilizaron cuatro ramos mixtos por nivel de frío, cada uno con 10 o más yemas en estado ‘F’ (flor abierta, con pétalos completamente extendidos) (Baggiolini, 1952; Reig et al., 2013). Los ensayos se realizaron entre el 13 y el 18 de setiembre de 2012, en función de la disponibilidad de flores en estado ‘F’ de cada cultivar evaluado. En forma simultánea y en cada ensayo se sometieron a heladas artificiales tres cultivares. Los ramos fueron recolectados en el mismo día de la aplicación de los tratamientos de frío en laboratorio, y conservados en bolsas plásticas. La simulación de la helada se realizó en un ‘freezer’ comercial marca Neba, con un controlador de temperatura marca Ako. Debido a la importancia de simular las heladas naturales en los ensayos de laboratorio (Gusta et al., 2003) la temperatura fue reducida a una tasa de 2 °C/h, retirando los distintos tratamientos de frío luego de 1 h a temperatura constante de –2, –3 y –4 °C. Para evitar la estratificación del aire y lograr así la homogeneización de la temperatura en su volumen, se incorporó un ventilador marca Morris (220 V, 3000 r.p.m. y 18 cm de diámetro). Como testigo para cada cultivar se colocó una muestra a temperatura ambiente. La temperatura del aire en el freezer se registró durante los experimentos con un sensor marca Hobo modelo U12, con un intervalo de 2 segundos entre mediciones.
Después de los tratamientos de frío, los ramos permanecieron durante 16 h a temperatura ambiente de laboratorio, con agua en la base de los frascos para evitar su deshidratación. Para la determinación del daño por frío se observaron los gineceos mediante lupa estereoscópica marca Zeiss modelo Stemi DV4 (32 X), considerando como dañados los tejidos de coloración marrón (Szalay et al., 2010). La TL50 se determinó gráficamente a partir de proyección vertical de la intersección entre la curva de daño según temperatura y la línea horizontal correspondiente a la proporción 0.5 de gineceos dañados.
Fenología
La fenología fue determinada en un ramo por cultivar durante la salida del reposo invernal de 2013; éste estaba localizado en la altura media del lado oeste de las plantas, con una exposición a la radiación solar similar. Para el registro fenológico se contó periódicamente el número de yemas florales correspondiente a cada estado de la escala de Baggiolini (1952) (Reig et al., 2013), expresando los resultados de cada fecha como porcentajes. La fecha de plena floración de cada cultivar correspondió a 50% de yemas florales en estado ‘F’.
Densidad de floración
El número de yemas florales se determinó al finalizar el invierno de 2013, tomando seis ramos —tres de cada lado, oeste y este— por cultivar (dos en cada una de tres plantas). Para el cálculo de la densidad de floración se dividió el número de yemas florales por la longitud de los ramos.
Registro de heladas en campo y evaluación del daño ocasionado
Durante la finalización del reposo invernal de 2013, la temperatura del aire fue registrada cada 30 minutos con un sensor digital marca I–Button, colocado en un abrigo meteorológico dentro del cuadro de cultivo a 1.5 m de altura. En tres plantas de cada cultivar en estudio, el 25 de setiembre de 2013 de las caras este y oeste de las copas se recolectaron seis ramos a 1.5 m de altura. La calificación de los daños en cada cultivar se hizo en 20 yemas florales elegidas al azar en distintos estados fenológicos. Para la determinación del daño se siguió el mismo criterio utilizado en la simulación de heladas en laboratorio (Szalay et al., 2010). Adicionalmente se registró el tipo de flor de cada cultivar, sobresaliendo la flor rosácea por sus pétalos grandes y vistosa, y la flor campanulácea por pétalos pequeños y estambres sobresalientes (UPOV, 2014). Para la discusión, los resultados se expresaron como supervivencia (número de flores sanas/cm de ramo), calculada a partir del porcentaje de flores sanas y la densidad de floración.
Diseño experimental y análisis estadístico
Para la determinación de la TL50 en laboratorio se utilizó un diseño experimental completamente aleatorizado, con cuatro repeticiones (ramos) por tratamiento; mientras que para la determinación de la densidad de floración se utilizó este mismo diseño con seis repeticiones. El análisis estadístico de los datos se realizó con el programa InfoStat/Profesional (Universidad Nacional de Córdoba, 2006). Para la densidad de floración de los cultivares se hizo un análisis de varianza, y para calcular la significancia de las diferencias entre medias, la prueba de Duncan (P < 0.05). Los daños por heladas en campo se analizaron mediante tablas de contingencia, evaluando 20 yemas florales por cultivar según la presencia o ausencia de daño.
Resultados
Temperatura letal media en laboratorio
La determinación del daño floral se hizo por el método propuesto por Szalay et al. (2010) con la utilización de lupa estereoscópica y permitió distinguir claramente entre gineceos sanos y dañados por temperaturas bajo cero (Figura 1).

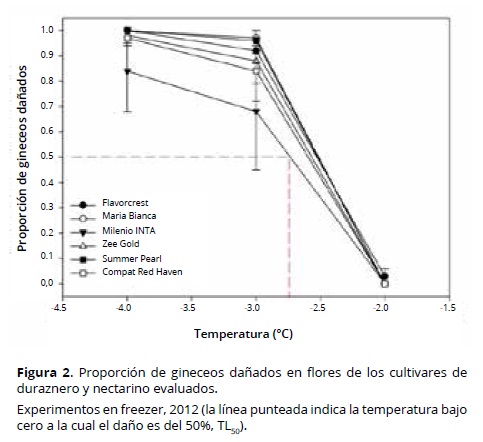
Los pistilos dañados presentaron una coloración marrón, mientras que los sanos permanecieron verdes 16 h después de la aplicación de los tratamientos de frío. Para cada nivel de temperatura, la proporción de gineceos dañados varió según el cultivar. Los valores observados de TL50 en el estado ‘F’ permitieron distinguir cultivares por la resistencia relativa de sus yemas florales a temperaturas bajo cero (Figura 2).
Fenología y densidad de floración
La fecha de plena floración varió ampliamente entre los cultivares de duraznero y nectarino evaluados, con una ocurrencia de 50% flores abiertas entre el 9 y el 29 de setiembre (Tabla 1).
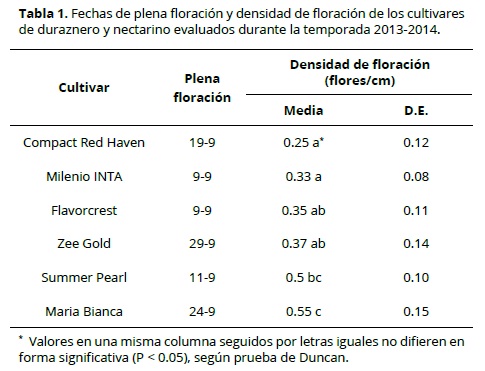
La densidad de floración presentó un amplio rango entre los cultivares evaluados (0.25 a 0.55 flores/cm). Los cvs Compact Red Haven y Milenio INTA presentaron la menor densidad de floración (0.25 y 0.33 flores/cm, respectivamente), mientras que los cvs Maria Bianca y Summer Pearl presentaron los mayores valores (0.55 y 0.5 flores/cm, respectivamente). Se debe mencionar que la mayor densidad de floración favorece una mayor producción de frutos frente a porcentajes similares de daño por heladas.
Registro de heladas en campo y evaluación del daño
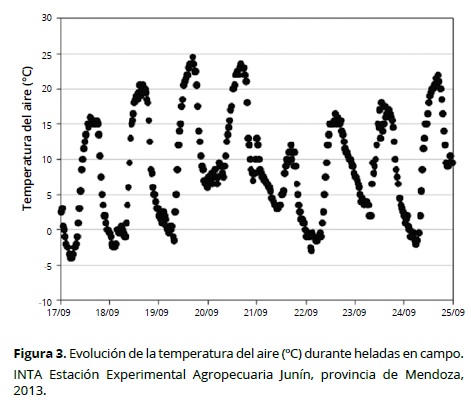
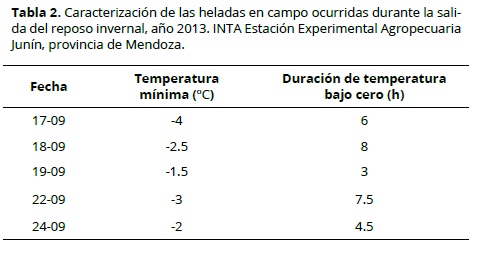
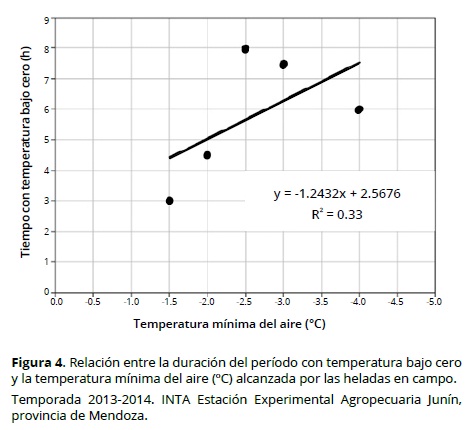
En la Figura 3 se observa la variación temporal de la temperatura del aire en la parcela experimental, durante el final del reposo invernal de los distintos cultivares de duraznero evaluados. Los eventos ocurridos de temperaturas bajo cero aparecen en la Tabla 2. Se observó que las heladas más intensas fueron también, en general, más extensas (Figura 4), no obstante el ajuste por regresión lineal fue bajo (R2 = 0.33).
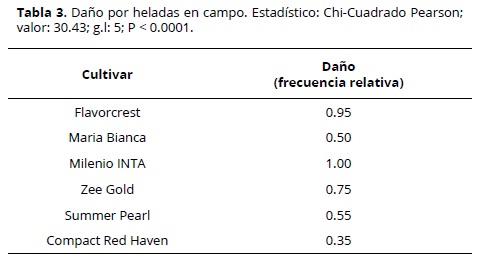
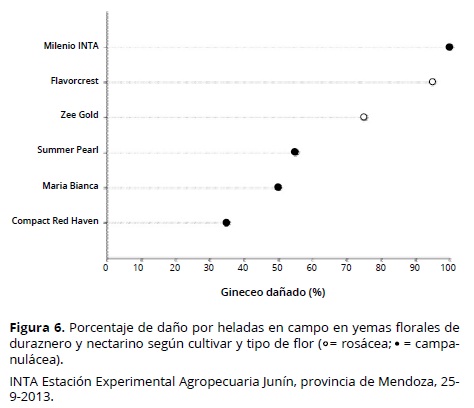
Se encontró una relación entre el cultivar y el daño observado en campo (P < 0.0001), variando entre 35 y 100% de las yemas florales observadas (Tabla 3). Las flores de durazneros y nectarinos pueden ser de tipo rosácea o campanulácea, siendo las primeras más grandes y vistosas (Figura 5). Los materiales que presentaron menor porcentaje de daño tienen, en su mayoría, tipo de flor campanulácea. La excepción fue el cv. Milenio INTA, con flor campanulácea y 100 % de flores dañadas (Figura 6).

Discusión
Fenología y densidad de floración
En distintos cultivares de duraznero conservero, Chaar et al. (2011) también observaron una amplia variabilidad en la fecha media de plena floración, la cual fluctuó entre el 28 de agosto y el 11 de septiembre. Okie y Werner (1996) observaron que el efecto varietal en la densidad de yemas florales en duraznero y nectarino fue marcadamente mayor al ambiental y a su interacción. Por el contrario, Kodad y Socias i Company (2008) observaron durante 3 años consecutivos en almendro (Prunus dulcis [Mill.] D. A. Webb) que la varianza debida al año fue la que más contribuyó a la varianza total de la densidad de floración.
Temperatura letal media en laboratorio
Buyukyilmaz y Kester (1976) observaron en cortes longitudinales de yemas y flores de almendro, que el pistilo fue el elemento más susceptible y utilizaron la coloración marrón como indicador de muerte del órgano reproductivo. De manera similar, Flinn y Ashworth (1994) determinaron el daño por congelación en yemas de arándano (Vaccinium corymbosum L.) a partir de la observación de coloración negra en los ovarios.
La ubicación relativa de los distintos materiales con respecto a la TL50 no puede generalizarse al resto de los estados fenológicos. Existe evidencia de cambios temporales en la tolerancia relativa a temperaturas bajo cero, en guindo (Prunus cerasus L.) (Mathers, 2004), en vid (Vitis vinifera L.) (Ferguson et al., 2011) y en almendro (Viti et al., 1994). Sin embargo, en climas templados como el de los oasis productivos de Mendoza, las temperaturas críticas de daño por congelación ocurren a la salida del reposo invernal, cuando las plantas no se encuentran aclimatadas (Szalay et al., 2010), por tanto el parámetro de TL50 durante el estado F es un buen indicador de la resistencia relativa de las yemas florales de los distintos cultivares.
Caracterización de la resistencia a temperaturas bajo cero
Los cvs Maria Bianca y Summer Pearl presentaron los valores más altos de flores sanas/cm de ramo (supervivencia de flores), después de la ocurrencia de temperaturas bajo cero en campo; mientras el cv María Bianca se caracterizó por una elevada densidad de floración (0.55 flores/cm) y floración tardía, como lo demuestra su plena floración el 24 de setiembre de 2013. El cv Summer Pearl sólo mostró una elevada densidad de floración (0.5 flores/cm) (Figura 7). El cv Compact Red Haven presentó un comportamiento aceptable frente a las temperaturas bajo cero (Figura 7) y se relacionó principalmente con una TL50 media (–2.6 °C) (Figura 2) y floración tardía, por su fecha de plena floración el 19 de septiembre de 2013 (Tabla 1). El bajo porcentaje de daño en campo (35%, Tabla 3) fue compensado por su baja densidad de floración (0.25 flores/cm, Tabla 1).
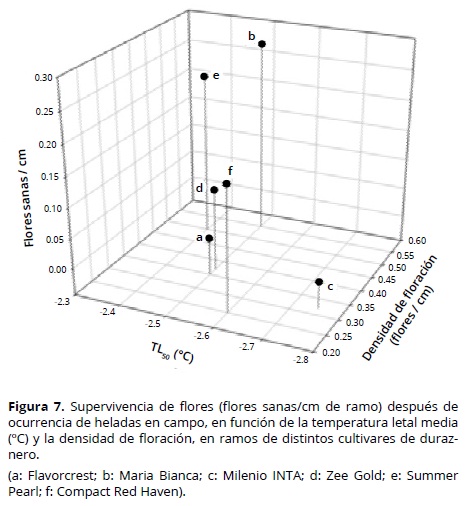
El cv Milenio INTA fue el menos resistente, con 100% de flores dañadas. Aunque presentó la TL50 más baja (alta tolerancia: –2.75 °C) (Figura 7), su floración fue la más temprana (Tabla 1), junto con el cv Flavorcrest, también con alto porcentaje de daño (Figura 6). Debido a su floración temprana, las yemas florales fueron más susceptibles a la congelación. Chaar et al. (2011) consideran que la utilización de materiales de duraznero conservero de floración tardía puede representar una ventaja en zonas con elevado riesgo de heladas primaverales. En este sentido, Rowland et al. (2005) observaron en yemas de arándano una reducción en la resistencia al frío al avanzar las fases de apertura de yemas. Sin embargo, la floración tardía no es por sí sola una característica de resistencia; por ejemplo, el cv Zee Gold presentó su plena floración más tarde que el cv Maria Bianca (Tabla 1), pero la TL50 media (–2.5 °C) y la media densidad de floración (0.37 flores/cm) del primero se relacionan con un elevado daño en campo (75%) y una moderada cantidad inicial de flores, lo que resultó en una supervivencia tan baja como 0.0925 flores/cm de ramo (Figura 7). Mather et al. (1980) observaron en grosellero negro (Ribes nigrum L.) que en años con heladas tardías, los cultivares que poseían floración tardía como única característica de resistencia fueron gravemente dañados. Lamb (1982) observó en peral (Pyrus communis L.) una tendencia de mayor daño por heladas en campo en cultivares de floración temprana, pero dentro de una misma época de floración se presentaron cultivares con distinto grado de supervivencia de flores. Según Imani y Khani (2011) el método indirecto más efectivo para evitar el daño por heladas en almendro consiste en la selección de los cultivares menos susceptibles en los mismos estados fenológicos. Kodad y Socias i Company (2005) mencionan que en almendro el elevado potencial para producir flores puede compensar los daños por heladas y junto con una elevada resistencia intrínseca al frío, puede mantener la frutificación en niveles comercialmente aceptables.
Los mecanismos responsables de resistencia a heladas pueden dividirse en evitación y tolerancia. La evitación o escape es la capacidad de la planta para evitar la formación de hielo en los tejidos, por ejemplo mediante sobre–enfriamiento. La tolerancia es la capacidad de la planta de sobrevivir a la formación de hielo extracelular y la consecuente deshidratación celular sin sufrir daño irreversible (Cary, 1985; Jacobsen et al., 2007). Es importante señalar que los resultados en este trabajo muestran en duraznero una combinación de tolerancia y evitación, ya que en los materiales resistentes ninguno tuvo como única característica sobresaliente la fecha de floración tardía. En forma similar Kodad y Socias i Company (2005) citan que en almendro la floración tardía no siempre es suficiente para superar los daños de las heladas, por lo que la resistencia intrínseca puede ser considerada también un objetivo de selección. Tanto la evitación como la tolerancia se encuentran bajo control genético y han evolucionado en respuesta a presiones de selección (Palonen y Buszard, 1997; Gusta y Wisniewski, 2013). No obstante los mecanismos de resistencia de un determinado órgano no son constantes a lo largo del ciclo anual, sino que pueden variar de forma temporal; al respecto, Pramsohler y Neuner (2013) observaron que yemas de manzano (Malus domestica Borkh.) presentaron congelación extra–órgano durante el reposo invernal, mientras que durante la ruptura de yemas ocurrió sobre–enfriamiento, el cual no se detectó posteriormente durante la plena floración. Ashworth et al. (1989) propusieron que la separación espacial del hielo dentro de las yemas de duraznero en reposo es consecuencia de la morfología de las yemas y del desarrollo vascular. Durante el reposo invernal los tejidos vasculares en órganos florales no están totalmente diferenciados y la continuidad xilemática entre los órganos florales y los tejidos subyacentes no está establecida.
Según Arora y Rowland (2011) el cambio climático global pronosticado puede tener graves efectos en el grado y ubicación de daños por frío, debido al probable aumento a futuro en la frecuencia de fenómenos meteorológicos. Las plantas capaces de resistir una prematura desaclimatación y reaclimatarse rápidamente cuando vuelve el frío, podrían tener mayores posibilidades de sobrevivir inviernos intensos. Al respecto, Chaar y Astorga (2012) mencionan que los cultivares más plásticos, por la posibilidad de lograr cosechas aceptables en zonas geográficas diversas, son aquellos que poseen bajo requerimiento de frío y elevado requerimiento de calor. Mientras que el bajo requerimiento de frío lo pueden satisfacer aún en zonas con inviernos poco fríos, el elevado requerimiento de calor se manifiesta en la floración tardía, lo cual posibilita evadir heladas tardías.
Conclusiones
- En los materiales estudiados de duraznero para consumo en fresco, la resistencia a heladas en campo se relacionó principalmente con la elevada densidad de floración, en combinación, en algunos casos, con floración tardía.
- La floración tardía por sí sola no resultó una característica de resistencia, por tanto, para la elección de cultivares de duraznero con menor riesgo de daño por temperaturas bajo cero, es importante tener en cuenta más de una variable relacionada con los órganos reproductivos, como la densidad y la fecha de floración y la temperatura letal media.
Agradecimientos
El presente estudio fue financiado por el Proyecto CIAC–940142, en el marco del Convenio INTA–AUDEAS–CONADEV.
Referencias
Arora, R.y Rowland, L. 2011. Physiological research on winter–hardiness: deacclimation resistance, reacclimation ability, photoprotection strategies, and a cold acclimation protocol design. Hort. Sci. 46(8):1070–1078.
Ashworth, E.; Davis, G.; y Wisniewski, M. 1989. The formation and distribution of ice within dormant and deacclimated peach flower buds. Plant Cell and Environ. 12:521–528.
Baggiolini, M. 1952. Stade repères du pecher. Revue Romande d’Agriculture, Viticulture et Arboriculture 4:29.
Buyukyilmaz, M.y Kester, D. 1976. Comparative hardiness of flower buds and blossoms of some almond genotypes in relation to time of bloom and leafing. J. Amer. Soc. Hort. Sci. 101(4):344–347.
Cary, J. 1985. Freeze survival in peach and prune flowers. Plant Sci. Letters 37:265–271.
Chaar, J.; Astorga, D.; y Reta, A. 2011. Determinación de la fecha media de plena floración de los principales cultivares de duraznero para industria [Prunus persica (L.) Batsch.] en el oasis Este de Mendoza. XXXIV Congreso Argentino de Horticultura. Buenos Aires. ASAHO Editores. 195 p.
Chaar, J. y Astorga, D. 2012. Determinación del requerimiento de frío y de calor en duraznero [Prunus persica (L.) Batsch.] mediante un modelo de correlación. RIA 38(3):289–298.
Chaar, J. 2013. Resistencia a heladas en plantas frutales. AIA 17(3):109–121.
Dale, A. 1987. Some studies in spring frost tolerance in black currants (Ribes nigrum L.). Euphytica 36:775–781.
Ferguson, J.; Tarara, J.; Mills, L.; Grove, G.; y Keller M. 2011. Dynamic thermal time model of cold hardiness for dormant grapevine buds. Ann. Bot. 107:389–396.
Fiorino, P.y Mancuso, S. 2000. Differential thermal analysis, supercooling and cell viability in organs of Olea europaea at subzero temperatures. Adv. Hort. Sci. 14:23–27.
Flinn, C.y Ashworth, E. 1994. Blueberry flower–bud hardiness is not estimated by differential thermal analysis. J. Amer. Soc. Hort. Sci. 119(2):295–298.
Granger, R.y Rousselle, G. 1984. Cold hardiness rating of pear cultivars and selections. Acta Hort. 161:69–72.
Gusta, L.; Wisniewski, M.; Nesbitt, N.; y Tanino, K. 2003. Factors to consider in artificial freeze tests. Acta Hort. 618:493–507.
Gusta, L.y Wisniewski, M. 2013. Understanding plant cold hardiness: an opinion. Physiol. Plant. 147(1):4–14.
Imani, A.y Khani, Y. 2011. Characteristics of almond selections in relation to late frost spring. Intern. J. Nuts Related Sci. 2(2):77–80.
InfoStat. 2006. InfoStat versión 2006. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
Jacobsen, S.; Monteros, C.; Corcuera, L.; Bravo, L.; Christiansen, J.; y Mujica, A. 2007. Frost resistance mechanisms in quinoa (Chenopodium quinoa Willd.). Eur. J. Agron. 26:471–475.
Kadir, S.y Proebsting, E. 1994. Various freezing strategies of flower–bud hardiness in Prunus. J. Amer. Soc. Hort. Sci. 119(3):584–588.
Kodad, O.; Morales, F.; y Socias, R. 2010. Evaluación de la tolerancia de las flores de almendro a las heladas por la fluorescencia de clorofila. ITEA 106(2):142–150.
Kodad, O. y Socias i Company, R. 2005. Daños diferenciales por heladas en flores y frutos y criterios de selección para la tolerancia a heladas en almendro. ITEA 101(4):349–363.
Kodad, O. y Socias i Company, R. 2008. Significance of flower bud density for cultivar evaluation in almond. Hort. Sci. 43(6):1753–1758.
Lamb, R. 1982. Flower bud survival of pear cultivars following a spring frost. Acta Hort. 124:27–31.
Lindén, L. 2002. Measuring cold hardiness in woody plants. Academic Dissertation, University of Helsinki. Department of Applied Biology. Publication No. 10.
Lisek, J. 2007. Frost damage of grapevines in Poland following the winter of 2005/2006. Folia Hortic. 19(2):69–78.
Mathers, H. 2004. Supercooling and cold hardiness in sour cherry germplasm: flower buds. J. Amer. Soc. Hort. Sci. 129(5):675–681.
Mather, P.; ModlibowskaI.; y Keep, E. 1980. Spring frost resistance in black currants (Ribes nigrum L.). Euphytica 29:793–800.
Okie, W.y Werner, D. 1996. Genetic influence on flower bud density in peach and nectarine exceeds that of environment. Hort. Sci. 31(6):1010–1012.
Palonen, P.y Buszard, D. 1997. Current state of cold hardiness research on fruit crops. Can. J. Plant Sci. 77:399–420.
Pramsohler, M.y Neuner, G. 2013. Dehydration and osmotic adjustment in apple stem tissue during winter as it relates to the frost resistance of buds. Tree Physiol. 33(8):807–816.
Quamme, H.; Layne, R.; y Ronald, W. 1982. Relationship of supercooling to cold hardiness and the northern distribution of several cultivated and native Prunus species and hybrids. Can. J. Plant Sci. 62:137–148.
Reig, G.; Alegre, S.; Gatius, F.; e Iglesias, I. 2013. Agronomical performance under Mediterranean climatic conditions among peach [Prunus persica L. (Batsch)] cultivars originated from different breeding programmes. Sci. Hort. 150:267–277.
Rowland, L.; Ogden, E.; Ehlenfeldt, M.; y Vinyard, B. 2005. Cold hardiness, deacclimation kinetics, and bud development among 12 diverse blueberry genotypes under field conditions. J. Amer. Soc. Hort. Sci. 130(4):508–514.
Szalay, L.; Timon, B.; Németh, S.; Papp, J.; y Tóth, M. 2010. Hardening and dehardening of peach flower buds. Hort. Sci. 45(5):761–765.
UPOV (Unión Internacional para la Protección de las Obtenciones Vegetales). 2014. Durazno/melocotonero Prunus persica (L.) Batsch). Directrices para la ejecución del examen de la distinción, la homogeneidad y la estabilidad. TG/53/7 Rev.
Viti, R.; Bartolini, S.; y Giorgelli, F. 1994. Effect of low temperatures on flower buds of several almond cultivars. Acta Hort. 373:193–199.
Referencias
Ashworth E., Davis G., Wisniewski M. 1989. The formation and distribution of ice within dormant and deacclimated peach flower buds. Plant, Cell and Environment 12:521-528.
Baggiolini M. 1952. Stade repères du pecher. Revue Romande d’Agriculture, Viticulture et Arboriculture 4:29.
Buyukyilmaz M., Kester D. 1976. Comparative hardiness of flower buds and blossoms of some almond genotypes in relation to time of bloom and leafing. J. Amer. Soc. Hort. Sci. 101(4):344-347.
Chaar J. 2013. Resistencia a heladas en plantas frutales. AIA 17(3):109-121.
Chaar J., Astorga D. 2012. Determinación del requerimiento de frío y de calor en duraznero [Prunus persica (L.) Batsch.] mediante un modelo de correlación. RIA 38(3):289-298.
Chaar J., Astorga D., Reta A. 2011. Determinación de la fecha media de plena floración de los principales cultivares de duraznero para industria [Prunus persica (L.) Batsch.] en el oasis Este de Mendoza. XXXIV Congreso Argentino de Horticultura. Buenos Aires. ASAHO Editores. 195 pp.
Dale A. 1987. Some studies in spring frost tolerance in black currants (Ribes nigrum L.). Euphytica 36:775-781.
Ferguson J., Tarara J., Mills L., Grove G., Keller M. 2011. Dynamic thermal time model of cold hardiness for dormant grapevine buds. Annals of Botany 107:389-396.
Fiorino P., Mancuso S. 2000. Differential thermal analysis, supercooling and cell viability in organs of Olea europaea at subzero temperatures. Adv. Hort. Sci. 14:23-27.
Granger R., Rousselle G. 1984. Cold hardiness rating of pear cultivars and selections. Acta Hort. 161:69-72.
Gusta L., Wisniewski M. 2013. Understanding plant cold hardiness: an opinion. Physiol. Plant. 147(1):4-14.
Gusta L., Wisniewski M., Nesbitt N., Tanino K. 2003. Factors to consider in artificial freeze tests. Acta Hort. 618:493-507.
Imani A., Khani Y. 2011. Characteristics of almond selections in relation to late frost spring. International Journal of Nuts and Related Sciences 2(2):77-80.
Kadir S., Proebsting E. 1994. Various freezing strategies of flower-bud hardiness in Prunus. J. Amer. Soc. Hort. Sci. 119(3):584-588.
Kodad O., Morales F., Socias R. 2010. Evaluación de la tolerancia de las flores de almendro a las heladas por la fluorescencia de clorofila. ITEA 106(2):142-150.
Kodad O., Socias i Company R. 2005. Daños diferenciales por heladas en flores y frutos y criterios de selección para la tolerancia a heladas en almendro. ITEA 101(4):349-363.
Lamb R. 1982. Flower bud survival of pear cultivars following a spring frost. Acta Hort. 124:27-31.
Lindén L. 2002. Measuring cold hardiness in woody plants. Academic Dissertation, University of Helsinki. Department of Applied Biology. Publication Nº 10.
Lisek J. 2007. Frost damage of grapevines in Poland following the winter of 2005/2006. Folia Horticulturae 19(2):69-78.
Mather P., Modlibowska I., Keep E. 1980. Spring frost resistance in black currants (Ribes nigrum L.). Euphytica 29:793-800.
Mathers H. 2004. Supercooling and cold hardiness in sour cherry germplasm: flower buds. J. Amer. Soc. Hort. Sci. 129(5):675-681.
Okie W., Werner D. 1996. Genetic influence on flower bud density in peach and nectarine exceeds that of environment. HortSci. 31(6):1010-1012.
Palonen P., Buszard D. 1997. Current state of cold hardiness research on fruit crops. Can. J. Plant Sci. 77:399-420.
Pramsohler M., Neuner G. 2013. Dehydration and osmotic adjustment in apple stem tissue during winter as it relates to the frost resistance of buds. Tree Physiology 33(8):807-816.
Quamme H., Layne R., Ronald W. 1982. Relationship of supercooling to cold hardiness and the northern distribution of several cultivated and native Prunus species and hybrids. Can. J. Plant Sci. 62:137-148.
Rowland L., Ogden E., Ehlenfeldt M., Vinyard B. 2005. Cold hardiness, deacclimation kinetics, and bud development among 12 diverse blueberry genotypes under field conditions. J. Amer. Soc. Hort. Sci. 130(4):508-514.
Szalay L., Timon B., Németh S., Papp J., Tóth M. 2010. Hardening and dehardening of peach flower buds. HortSci. 45(5):761-765.
Viti R., Bartolini S., Giorgelli F. 1994. Effect of low temperatures on flower buds of several almond cultivars. Acta Hort. 373:193-199.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Danielle A. Lake Diver, Jessica A. Savage. (2024). Weighing the risks and benefits of flowering early in the spring for the woody perennial Prunus pumila. American Journal of Botany, 111(11) https://doi.org/10.1002/ajb2.16417.
2. Michele Antonio Savoia, Loredana Del Faro, Andrea Turco, Valentina Fanelli, Pasquale Venerito, Cinzia Montemurro, Wilma Sabetta. (2023). Biodiversity Evaluation and Preservation of Italian Stone Fruit Germplasm (Peach and Apricot) in Southern Italy. Plants, 12(6), p.1279. https://doi.org/10.3390/plants12061279.
3. Ekaterina Vodiasova, Elina Chelebieva, Ekaterina Kladchenko, Oksana Grebennikova, Victoria Uppe, Valentina Tsiupka, Sergey Dolgov, Anatoly Smykov. (2025). Changes of Catalase and Peroxidase Activity and Expression Under Cold Stress in Prunus persica Cultivars with Different Cold Tolerances. Agronomy, 15(3), p.556. https://doi.org/10.3390/agronomy15030556.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2015 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.






















