


Publicado
REGENERACIÓN DE PALMAS EN BOSQUES NATIVOS Y PLANTACIONES DEL SANTUARIO DE FAUNA Y FLORA OTÚN – QUIMBAYA (RISARALDA, COLOMBIA)
Regeneration of palms in native forests and plantations at Otún - Quimbaya Fauna and Flora Sanctuary (Risaralda, Colombia)
DIEGO F. CORREA-GÓMEZ
ORLANDO VARGAS-RÍOS
Departamento de Biología, Universidad Nacional de Colombia, Bogotá, Colombia. diegofelipecorrea@gmail.com; jovargasr@unal.edu.co
RESUMEN
Las palmas constituyen un grupo clave para comprender los procesos de regeneración en bosques montanos neotropicales, por su alta representatividad estructural, funcional y ecológica. En el Santuario de Fauna y Flora Otún-Quimbaya (SFFOQ) se evaluó la regeneración de palmas a partir de cinco transectos de 50 x 4 m (0,1 ha) por tipo de bosque: bosque nativo maduro (BNM), bosque nativo secundario (BNS), plantación forestal de pino (Pinus patula) (PP), de roble (Quercus humboldtii) (PR) y de urapán (Fraxinus chinensis) (PU). Se censaron todos los individuos por especie, desde plántulas hasta adultos, tomando como variables la diversidad, la frecuencia de aparición de especies y la estructura poblacional por tipo de bosque. La riqueza encontrada (diez especies) fue alta en comparación con estudios realizados en otros bosques montanos. La especie más abundante fue Geonoma undata, con cerca de la mitad del total de individuos muestreados, seguida de Prestoea acuminata, Chamaedorea pinnatifrons, Ceroxylon alpinum, Wettinia kalbreyeri, Chamaedorea linearis, Geonoma jussieuana, Aiphanes simplex, Geonoma orbignyana y Aiphanes lindeniana. El BNM presentó la mayor diversidad (Shannon ln, H´ = 1,83, recíproco del índice de Simspon, 1/D = 5,43), seguido por el BNS, la PP, PR y PU. También presentó la mayor riqueza (diez especies), seguido por el BNS, la PU, PP y PP. La mayor diversidad y riqueza de palmas en el BNM se relaciona con el mayor tiempo para el reclutamiento y la persistencia de las especies, así como con la mayor heterogeneidad estructural y funcional. La menor diversidad y riqueza de las plantaciones se relaciona con la menor heterogeneidad estructural y funcional, y con las restricciones edáficas en robledales y pineras. La presencia de individuos de G. undata, C. alpinum y Ch. pinnatifrons en todos los tipos de bosque, con las mayores frecuencias de aparición por total de transectos (fr=0,88, 0,72 y 0,72 respectivamente) indica que estas especies tienen un alto potencial para colonizar ambientes distintos. En la PU se encuentran las mejores condiciones para la germinación y el reclutamiento de Ch. linearis, G. undata, Ch. pinnatifrons y C. alpinum, según lo demuestra el mayor porcentaje de plántulas e individuos jóvenes encontrados entre tipos de bosque (70,2%, 69%, 49,8% y 37,1% respectivamente), lo que probablemente se relaciona con la menor varianza y/o mayor disponibilidad lumínica, o con la presencia de abundante hojarasca que favorece la germinación. Al contener la totalidad de especies y el mayor porcentaje de adultos por especie, el BNM es clave para la producción de semillas y regeneración de las palmas del SFFOQ.
Palabras clave. Arecaceae, Andes, Bosques Montanos, Diversidad, Plántulas, Reclutamiento.
ABSTRACT
Because of their structural, functional and ecological importance, palms are a key group in understanding the processes of regeneration in Neotropical montane forests. We studied the regeneration of palms in the Fauna and Flora Otún-Quimbaya Sanctuary along five transects of 50x4 m (0,1 ha) in five forest types: mature native forest (MNF), secondary forest (SF), pine plantation (Pinus patula) (PP), oak plantation (Quercus humboldtii) (OP), and Chinese ash plantation (Fraxinus chinensis) (AP). We recorded all individuals of each species, including seedlings, juveniles and adults, taking as variables the diversity, frequency of occurrence and population structure by forest type. Species richness (ten species) was high compared with other studies conducted in similar mountain forests. The most abundant species was Geonoma undata, with nearly half of all individuals recorded, followed by Prestoea acuminata, Chamaedorea pinnatifrons, Ceroxylon alpinum, Wettinia kalbreyeri, Chamaedorea linearis, Geonoma jussieuana, Aiphanes simplex, Geonoma orbignyana and Aiphanes lindeniana. The MNF had the highest diversity (Shannon index H' = 1,83, Simpson's Reciprocal index 1/D = 5,43), followed by the SF, PP, OP and AP. The MNF also presented the highest richness (ten species), followed by the SF, AP, PP and OP. The high diversity and richness of palms in the MNF is associated with a longer recruitment time and persistence of species, as well as with the higher structural and functional heterogeneity. The low diversity and richness of the plantations are associated with the lower structural and functional heterogeneity, and the edaphic restrictions in the OP and PP. The presence of individuals of G. undata, C. alpinum and Ch. pinnatifrons in all types of forests, with the highest frequency of occurrence by total transects (fr = 0,88, 0,72 and 0,72 respectively), indicates that these species have a high potential to colonize different environments. The AP has the best conditions for the germination and recruitment of Ch. linearis, G. undata, Ch. pinnatifrons and C. alpinum, as evidenced by the higher percentage of seedlings and juveniles found among forest types (70,2%, 69%, 49,8% and 37,1% respectively), which is probably related to a low light variance and/or increased light availability, as well as abundant litter. Because the MNF has all palm species, with the highest percentage of adults per species, this type of forest is critical for seed production and palm regeneration in the Sanctuary.
Key words. Arecaceae, Andes, Mountain Forests, Diversity, Seedlings, Recruitment.
Recibido: 16/10/2008
Aceptado: 19/05/2009
INTRODUCCIÓN
La diversidad de las comunidades vegetales, es el resultado de la interacción entre el conjunto de propágulos que tienen la capacidad para llegar a un sitio determinado y las interacciones ecológicas locales (Lepš 2005), constituyéndose en la principal medida de la regeneración vegetal. El proceso de regeneración vegetal es complejo y dinámico, incluye la dispersión, el establecimiento y la persistencia de un conjunto de poblaciones a través de los ciclos sucesionales (Glenn-Lewin & van der Maarel 1992, van der Valk 1992), y como tal, depende de la historia de vida de las especies (Caswell 1989) y de su interacción con el medio ambiente y biológico circundante (Noble & Slatyer 1980).
De acuerdo con Tomlinson (1990) y Henderson (2002), las palmas constituyen un grupo interesante para estudiar los procesos de regeneración en bosques tropicales, como resultado de su amplia distribución, suficiente definición taxonómica, alta representatividad en todo tipo de hábitats, y alta diversificación estructural, anatómica, fisiológica y ecológica. De 600 especies de palmas conocidas en Sudamérica, 86 especies pueden encontrarse por encima de los 1000 m de altitud (Moraes et al. 1995). Aunque su riqueza es menor en comparación con las tierras bajas (Moraes et al. 1995, Borschsenius et al. 1998), hacen parte de las familias andinas más frecuentes, llegando a constituir una importante fracción del total de especies con diámetro a la altura del pecho (DAP) mayor o igual a 2,5 cm registradas en muestras de bosques andinos premontanos y montanos (Gentry 1995). Varios géneros de palmas se han diversificado ampliamente en las montañas del neotrópico (Henderson et al. 1995, Moraes et al. 1995), incluyendo los géneros Chamaedorea y Geonoma, con alta frecuencia de aparición en transectos de bosques andinos (Gentry 1995), el género exclusivamente andino Ceroxylon, y los géneros Prestoea, Wettinia y Aiphanes (Moraes et al. 1995). Además, varias especies pueden llegar a ser dominantes en bosques húmedos andinos (Borschsenius et al. 1998) y formar una importante fracción del sotobosque (Valencia 1995, Svenning 2001). Son, por lo tanto, elementos estructurales y funcionales indispensables dentro de la dinámica de bosques montanos neotropicales, constituyendo un grupo apropiado para estudiar la estructura de las comunidades andinas (Svenning 2001).
La regeneración de palmas se puede correlacionar con una gran cantidad de variables ecológicas, que son el resultado del éxito en dispersión, establecimiento y persistencia de las especies. La dispersión de palmas a largas distancias, por agentes abióticos, parece ser poco probable (Uhl & Moore 1973), y por lo tanto la zoocoria constituye el principal mecanismo de dispersión (Zona & Henderson 1989). La germinación y el establecimiento están sujetos a procesos de depredación posdispersión de semillas (Brown 1976, Lott et al. 1995), germinación diferencial por sensibilidad a factores edáficos (Svenning 2001), climáticos y lumínicos (Vandermeer et al. 1979, Bullock 1980, Borschsenius et al 1998, Svenning 2001), daño mecánico (Ponce et al. 1999), depredación (Lott et al. 1995) y competencia interespecífica e intraespecífica de plántulas (Vandermeer et al. 1979, Svenning 2001).
Una vez la población se ha establecido, su persistencia depende de la capacidad para reemplazar los individuos que mueren, a través de la producción de semillas y/o propagación vegetativa (van der Valk 1992), proceso que se puede estudiar a través de la estructura poblacional.
Dados los múltiples factores que intervienen en la regeneración, es de esperar diferencias en la estructura de las comunidades sujetas a condiciones ambientales y biológicas distintas (Lugo 1992, Sawyer 1993, Murcia 1997, Cavelier & Santos 1999), y por lo tanto cambios en la diversidad y la estructura poblacional de palmas como respuesta a diferencias en la dispersión, el establecimiento y la persistencia de las especies. En el presente estudio se evaluó la regeneración diferencial de palmas en cinco tipos de bosque: bosque nativo maduro (BNM), bosque nativo secundario (BNS), plantación forestal de pino (Pinus patula) (PP), de roble (Quercus humboldtii) (PR) y de urapán (Fraxinus chinensis) (PU) del Santuario de Fauna y Flora Otún - Quimbaya (SFFOQ), tomando como variables la diversidad, la frecuencia de aparición de especies, y la estructura poblacional.
El estudio de la regeneración de palmas, es indispensable para proponer estrategias de conservación para las poblaciones existentes dentro del SFFOQ. Además, las palmas constituyen un grupo focal para comprender la complejidad propia de los bosques montanos neotropicales, que se encuentran entre los más amenazados y menos conocidos del neotrópico (Gentry 1995), a pesar de su alta riqueza biológica.
MATERIALES Y MÉTODOS
Área de estudio. El estudio se desarrolló en el Santuario de Fauna y Flora Otún-Quimbaya (SFFOQ), localizado en la vereda La Suiza, municipio de Pereira, departamento de Risaralda (Fig. 1). El SFFOQ hace parte del Sistema de Parques Nacionales Naturales de Colombia (UAESPNN), tiene un área de 410,97 ha, y está situado entre 1750 y 2250 m de altitud sobre la vertiente occidental de la Cordillera Central (Guerrero et al. 2004). Corresponde a bosque muy húmedo montano bajo (bmh-MB) de acuerdo con la zonificación de Holdridge (Espinal & Montenegro 1963) y a bosque subandino de acuerdo con la clasificación de Cuatrecasas (Rangel-Ch. et al. 1997). En la estación climatológica El Cedral (4º52´ Norte, 75º32´ Oeste, 2120 m de altitud), la temperatura media anual registrada entre 1989 y 2003 es de 15,3ºC, la humedad relativa equivale al 86,3%, y la precipitación media anual es de 2556 mm. El régimen de lluvias es bimodal, con máximos de precipitación de octubre a diciembre y de marzo a mayo (Anónimo 1989-2003).
Fase de campo. Con base en los mapas de cobertura vegetal disponibles para la zona (Anónimo 2003), y con información de campo (feb 2005) se determinaron los tipos de bosque a estudiar. De acuerdo con Guerrero et al. (2004) el BNS y las plantaciones tienen de 30 a 50 años. Adicionalmente se ubicaron individuos reproductivos por especie (a excepción de G. orbignyana, de la cual no existían registros para el SFFOQ), lo que permitió identificar las plántulas. Se trazaron cinco transectos (15 sep - 15 oct 2005) de 50x4 m (0,1 ha por tipo de bosque), censando todos los individuos por especie de palma, desde plántulas hasta adultos. En las especies clonales se tomaron los datos de abundancia de los individuos genéticamente distintos (genets), que reflejan el éxito en dispersión. Los transectos se trazaron al azar, en dirección perpendicular a los cursos de agua y/o caminos, paralelos entre sí y distanciados a más de 50 m.
Para el análisis poblacional en los individuos sin tallo se contó el número de venas de la mitad lateral de la hoja más joven. En los individuos con tallo se contó el número de cicatrices, se midió el diámetro en la base, y se determinó la presencia de estructuras reproductivas o cicatrices de estas estructuras. El número de venas está correlacionado con la edad de la palma (Vergara 2002, Rodríguez-Buriticá et al. 2005), así como el diámetro de la base del tallo durante la fase de ensanchamiento máximo (Henderson 2002) y el número de cicatrices (Tomlinson 1990), siendo variables útiles para establecer las clases de desarrollo (Plántulas, Individuos Jóvenes, Adultos).
Los ejemplares de palmas se recolectaron siguiendo la metodología propuesta por Dransfield (1986) y se depositaron en el Herbario Nacional Colombiano (COL) bajo los números de colección del primer autor.
Análisis de diversidad. Para el análisis de la diversidad de palmas por tipo de bosque se siguió la metodología propuesta por Magurran (1989): se determinó la representatividad de las muestras mediante una curva de acumulación de especies, y se realizó un análisis conjunto entre el ajuste a los modelos teóricos de abundancia de especies y varios índices de diversidad. El ajuste a los modelos (serie geométrica, serie logarítmica, serie normal logarítmica, y modelo "vara quebrada") se realizó mediante una prueba ?2 con nivel de significancia igual a 0,05, reduciendo los grados de libertad cuando por clase de abundancia se encontraron valores observados iguales a cero. Los índices de diversidad calculados mediante el programa BioDiversity Pro 5,1, correspondieron al índice de Shannon en base logaritmo natural (H'), al recíproco del índice de dominancia de Simpson (1/D), y al índice de equidad de Pielou (J´) (Moreno 2001).
Mediante el programa de estadística MINITAB 14 se realizó un
análisis de varianza con un nivel de significancia igual a 0,05 sobre
los índices calculados por transecto, para determinar diferencias significativas
entre los índices por tipo de bosque. Se probó la normalidad de
la población de índices mediante la prueba de Anderson Darling
(Montgomery 2000), y se probaron los supuestos de homocedasticidad y normalidad
de los residuos. Para que el índice de Shannon se ajustara a una distribución
normal se eliminaron los datos extremos, correspondientes a cero. Se realizaron
regresiones lineales entre todos los pares de índices calculados, incluyendo
el número de especies (S) y el número de individuos (N).
La diversidad Beta se determinó mediante el análisis de similitud
de Jaccard con el programa BioDiversity Pro 5,1. Se realizó una tabla
con la frecuencia de aparición de cada especie por el total de transectos
en cada tipo de bosque.
Análisis poblacional. Para cada especie, se determinaron tres clases de desarrollo (Plántulas, Individuos Jóvenes y Adultos), teniendo en cuenta las fases de desarrollo propuestas por Tomlinson (1990) y Henderson (2002) (fases de plántula, establecimiento, adulto vegetativo, y adulto reproductivo), y siguiendo criterios morfológicos. Mediante la construcción de un gráfico de secuencia de número de venas por lado para el total de individuos, se determinó el límite de la clase de desarrollo Plántulas, que de acuerdo con Bernal (1998) se caracteriza por la mayor mortalidad de individuos en la población (en el presente estudio 3-9 venas por lado para G. undata, 3-13 para C. alpinum, 3-5 para P. acuminata, 4-8 para Ch. pinnatifrons, 4-16 para Ch. linearis, 7-13 para W. kalbreyeri, 4-7 para G. jussieuana), valores que coinciden aproximadamente con el estudio realizado por Vergara (2002) para C. alpinum (3-12 venas por lado) y Gamba-Trimiño (2004) para P. acuminata (hojas bífidas, que en el presente estudio corresponden a 3-8 venas por lado). La clase de desarrollo Jóvenes incluyó los individuos sin estructuras reproductivas o sin rastros de estas, que continúan con la expansión gradual del axis hasta alcanzar el máximo diámetro posible del tallo, es decir la fase de establecimiento, y la posterior elongación del tallo y aumento del número de cicatrices, es decir la fase de adulto vegetativo. La clase de desarrollo Adultos incluyó los individuos reproductivos con tallo, es decir la fase de desarrollo adultos reproductivos. Se debe tener en cuenta que la ontogenia de las palmas es gradual, por lo que es arbitrario considerar límites marcados entre clases de desarrollo (Tomlinson 1990).
RESULTADOS
La inclusión de la totalidad de las especies de palmas registradas para el SFFOQ, y de un nuevo registro para la zona (Geonoma orbignyana), demuestra la representatividad de las 0,5 hectáreas estudiadas.
La estabilización de la curva de acumulación de especies muestra que 0,1 ha son suficientes para incluir las especies de palmas del BNS y PU (Fig. 2). En el BNM se incluyeron todas las especies, por lo que aunque la curva no se estabilice, el muestreo es completo. Aunque es necesario aumentar la superficie de muestreo en PR y PP, se pueden hacer comparaciones en diversidad con los otros tipos de bosque al contar con la misma área de muestro.

Riqueza y abundancia relativa. Se registraron diez especies de palmas. La mayor cantidad de especies se encontró en el BNM (S=10), seguido por el BNS (S=7), la PU (S=6), la PP (S=5) y la PR (S=4). El promedio de especies de palmas teniendo en cuenta todos los tipos de bosque correspondió a 6,4.
La especie más abundante fue Geonoma undata (Tabla 1), con cerca de la mitad del total de individuos, seguida por Prestoea acuminata, Chamaedorea pinnatifrons, Ceroxylon alpinum, Wettinia kalbreyeri, Chamaedorea. linearis, Geonoma jussieuana, Aiphanes simplex, Geonoma orbignyana y Aiphanes lindeniana.

El mayor porcentaje de individuos se encontró en la PU (50,3%), seguido por el BNS (25,1%), el BNM (14,4%), la PP (5,6%) y la PR (4,6%).
Diversidad y modelos de abundancia. El BNS presentó el mejor ajuste al modelo de serie logarítmica (?²=3,18, 5gl, p>0,70) (Fig. 3, Tabla 2), como resultado de la distribución de las especies Ch. pinnatifrons, C. alpinum, Ch. linearis, W. kalbreyeri y A. simplex en clases de abundancia intermedias a bajas, con valores del 17,9%, 5,6%, 2,1%, 1,1% y 0,2% del total de individuos para el tipo de bosque respectivamente.

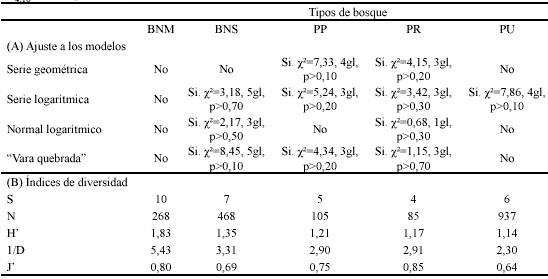
Así mismo, el ajuste de la PU al modelo de serie logarítmica (?²=7,86, 4gl, p>0,10), se relaciona con la abundancia intermedia a baja de las especies Ch. pinnatifrons, P. acuminata, C. alpinum, Ch. linearis y W. kalbreyeri, con el 15,0%, 11,2%, 7,3%, 3,5% y 0,2% del total de individuos para el tipo de bosque respectivamente. La PP y PR presentaron un mejor ajuste al modelo de "vara quebrada" (?²=4,34, 3gl, p>0,20 y ?²=1,15, 3gl, p>0,70 respectivamente), lo que muestra la distribución uniforme en clases de abundancia. El BNM estuvo cerca al ajuste del modelo "vara quebrada" (?²=11,24, 4gl, p<0,05).
El BNM presentó la mayor diversidad (H'=1,83, 1/D=5,43) (Tabla 2), seguido por el BNS (H'=1,35, 1/D=3,31). Aunque se encontró que estos índices están mutuamente explicados (r2=84,9%, F1,19=107,22, p<0,001) (Tabla 3), y son explicados por el número de especies (r2 = 67,7%, F1,19=39,79, p<0,001 para H'; r2 = 36,6%, F1,19=10,95, p=0,004 para 1/D), sólo el recíproco del índice de Simpson (1/D) estuvo explicado por el índice de equidad de Pielou J' (r2=41,4%, F1,19=13,41, p=0,002). Por lo tanto, la mayor diversidad de Shannon encontrada fue el resultado de la inclusión de la mayor cantidad de especies entre tipos de bosque (S=10 para el BNM, S=7 para BNS), y la mayor diversidad de Simpson (1/D) fue el resultado de dicho número de especies, además de la alta equidad (J'=0,80) para el BNM.

A pesar de contener la menor cantidad de especies (S=4), la mayor diversidad de Simpson de la PR (1/D=2,91), en relación con la PP (1/D=2,90), se relaciona con la alta equidad (J´=0,85), y con el mejor ajuste al modelo de "vara quebrada" entre tipos de bosque. El índice de Shannon, que es explicado por el número de especies, es por lo tanto menor en la PR (H'=1,21, S=5 para la PP; H'=1,17, S=4 para la PR).
Las PU, que fue la menos equitativa (J´=0,64) como resultado de la mayor dominancia de una especie entre tipos de bosque (G. undata, con el 62,8% del total de individuos), presentaron los menores valores de diversidad (1/D=2,30, H'=1,14). Por lo tanto la equidad está también explicando al índice de Shannon, aunque la regresión no sea significativa (r2=16,6%, F1,19=3,70, p=0,069).
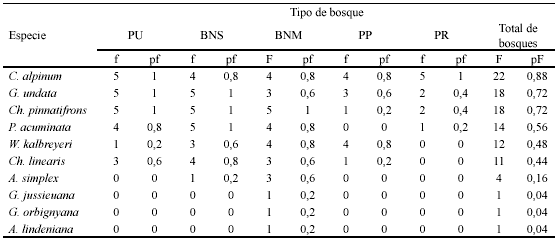
El BNS y la PU compartieron la mayor cantidad de especies (C. alpinum, G. undata, Ch. pinnatifrons, P. acuminata, W. kalbreyeri, Ch. linearis) (Fig. 4). P. acuminata se encontró en todos los tipos de bosque excepto la PP, y W. kalbreyeri y Ch. linearis se encontraron en todos los tipos de bosque excepto la PR. Sólo C. alpinum, G. undata y Ch. pinnatifrons estuvieron compartidas entre todos los tipos de bosque. Las especies propias a bosques nativos correspondieron a G. jussieuana, G. orbignyana y A. lindeniana (BNM) y A. simplex (compartida entre BNM y BNS).

Frecuencia y análisis poblacional. C. alpinum fue la especie con mayor frecuencia de aparición en todos los transectos muestreados, seguida de G. undata, Ch. pinnatifrons, P. acuminata, W. kalbreyeri y Ch. linearis (Tabla 4).
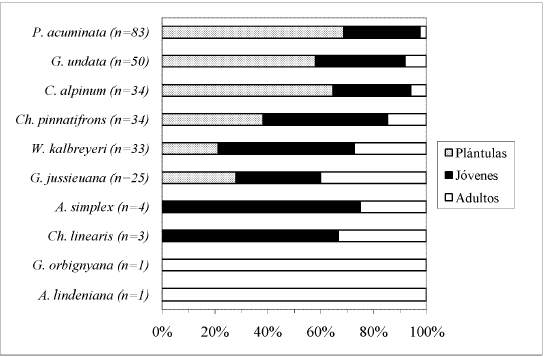
Al sumar los individuos de cada especie entre tipos de bosque, se encontró un bajo porcentaje de adultos (<2%) e individuos jóvenes (=40%) para G. undata, C. alpinum y P. acuminata. Para Ch. pinnatifrons el porcentaje adultos fue menor al 7%, y el de individuos jóvenes menor al 50%. Para Ch. linearis y W. kalbreyeri el porcentaje de adultos fue menor al 11%, y el de jóvenes mayor al 60%. Para G. jussieuana, A. simplex, G. orbignyana y A. lindeniana el porcentaje de adultos encontrado superó el 20%.
El mayor porcentaje de individuos por especie, dentro cada tipo de bosque, correspondió a la clase de desarrollo Plántulas (Fig. 5, Fig. 6, Fig. 7, Fig. 8 y Fig. 9), a excepción de A. lindeniana y G. orbignyana en el BNM (0%), Ch. linearis en el BNM, PP y PU (0%, 0% y 30,3% respectivamente), A. simplex en el BNM y BNS (0%), G. jussieuana en el BNM (28%), W. kalbreyeri en el BNM, BNS y PP (21,2%, 40% y 17,3% respectivamente), Ch. pinnatifrons en el BNM, PP y PU (38,2%, 25% y 42,5% respectivamente), y C. alpinum, G. undata y P. acuminata en la PP (28,9%, 14,3% y 22,2% respectivamente). Los mayores porcentajes de individuos adultos por especie se encontraron en los bosques nativos maduros.
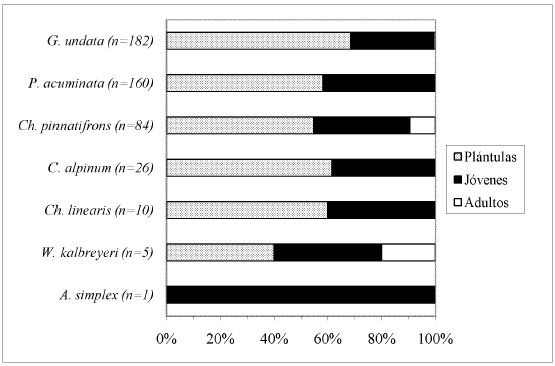

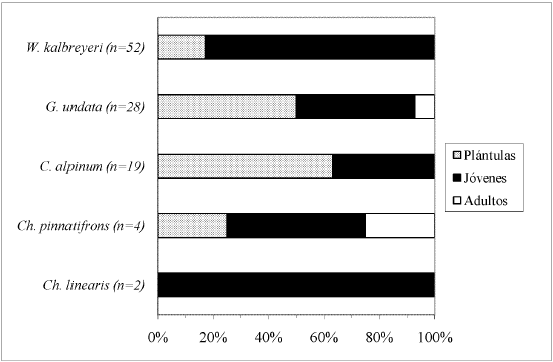
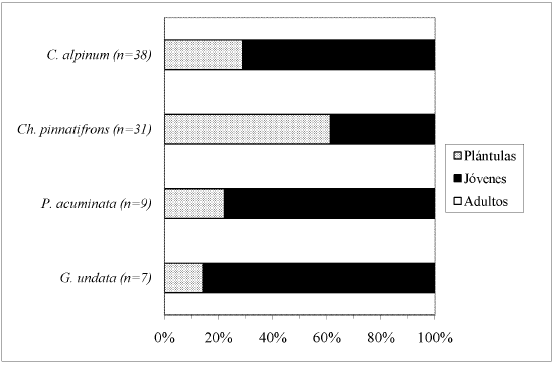
DISCUSIÓN
Riqueza y abundancia relativa. La riqueza de palmas del BNM (S=10) es superior a la encontrada por Gentry (1995) en las muestras de 0,1 ha de Murrí-Antioquia (960 m de altitud, S=9), Alto de Mira-Magdalena (1200 m de altitud, S=7), Alto de Cuevas-Antioquia (1710 m de altitud, S=6) y La Planada-Nariño (1810 m de altitud, S=6). Así mismo, es superior a la riqueza encontrada por Valencia (1995) sobre 1 ha de muestreo en Baeza-Ecuador (2000 m de altitud, S=3), y por Svenning (2001) en 167 parcelas de Maquipicuna-Ecuador (1200 m de altitud, S=7). Además, el promedio de especies (6,4) es superior al promedio de 2,7 especies encontrado por Gentry (1995) en 28 muestras de 0,1 ha en bosques premontanos y montanos sudamericanos (800 a 2900 m de altitud). La mayor riqueza encontrada puede ser el resultado de aspectos biogeográficos que determinan la presencia de mayor cantidad de especies en la zona, así como de sesgos por ausencia de muestreo de individuos con DAP < 2,5 cm en el estudio de Gentry (1995) y con DAP < 5 cm en el estudio de Valencia (1995), lo que en el presente estudio excluiría a adultos de Ch. pinnatifrons y A. simplex (DAP < 2,5 cm), G. jussieuana y G. orbignyana (DAP < 5 cm), además de individuos jóvenes y plántulas de todas las especies.
G. undata es un componente importante en bosques andinos. En el presente estudio
constituye el 45,9% del total de individuos de los tipos de bosques estudiados,
y en Baeza-Ecuador (Valencia 1995) conforma el 85,2% del total de palmas. P.
acuminata y Ch. pinnatifrons también son abundantes en el estudio de
Svenning (2001). Por su parte la alta abundancia de C. alpinum parece ser local.
Diversidad y modelos de abundancia. La mayor riqueza (S=10) y diversidad de
palmas en el BNM (H'=1,83, 1/D=5,43) en relación con el BNS (H´=1,35,
1/D=3,31) sigue la tendencia encontrada en sucesiones de bosques tropicales
(Lepš 2005), lo que se relaciona con el mayor tiempo del BNM para la regeneración
de las palmas. Además, la alta heterogeneidad estructural, resultado
de la formación de claros por caída de árboles (Hartshorn
1980, Sloan 1980, Nicotra et al. 1999), aumenta la probabilidad de reclutamiento
de palmas con historias de vida dependientes de disturbios cíclicos,
y la alta heterogeneidad funcional aumenta la complejidad de las interacciones
ecológicas (e.g. relaciones de dispersión), permitiendo la regeneración
de la mayor cantidad de especies de palmas entre tipos de bosque.
La cercanía al ajuste del modelo "vara quebrada" para el
BNM (?²=11,24, 4gl, p<0,05), indica la división de un recurso
de manera aleatoria entre pocas especies (Magurran 1989, Ricklefs & Millar
1999). La luz es un recurso limitante en el BNM. Por lo tanto, la alta varianza
en disponibilidad de luz por caída de árboles (Hartshorn 1980,
Sloan 1980, Nicotra et al. 1999) podría determinar la regeneración
de palmas en micrositios adecuados distribuidos al azar, y explicaría
la ausencia de alguna especie con marcada dominancia (alto índice de
equidad, J'=0,80). Si los claros están alejados, habría
menos micrositios disponibles, y por lo tanto menor probabilidad de reclutamiento,
lo que se refleja en la menor abundancia total en relación con la PU
y el BNS (14,4% del total de individuos muestreados), y en la menor abundancia
de plántulas e individuos jóvenes para P. acuminata, Ch. pinnatifrons,
G. undata y Ch. linearis en relación con la PU y el BNS (22,8%, 10,5%,
5,5%, y 4,2% del total de plántulas e individuos jóvenes entre
bosques). La germinación y el reclutamiento sólo es alto para
las especies propias G. orbignyana y G. jussieuana, para A. simplex (75% del
total de plántulas e individuos jóvenes entre tipos de bosque),
y para W. kalbreyeri (29,3% del total de plántulas e individuos jóvenes
entre tipos de bosque).
La reducción de micrositios con distintas intensidades lumínicas
en el BNS (Nicotra et al. 1999) podría explicar el ajuste a la serie
logarítmica (?²=3,18, 5gl, p>0,70). La colonización secuencial
podría determinar la dominancia de especies con alto potencial de germinación
y reclutamiento en condiciones de luz menos heterogéneas que en el BNS
(G. undata y P. acuminata, con el 38,9% y 34,2% del total de individuos para
el tipo de bosque respectivamente, índice de equidad J´=0,69).
La menor varianza en disponibilidad lumínica podría explicar la
alta abundancia de individuos (25,1% del total de individuos muestreados), el
mayor porcentaje de plántulas e individuos jóvenes de P. acuminata
entre tipos de bosque (45,1%), y el alto porcentaje de plántulas e individuos
jóvenes entre tipos de bosque para Ch. pinnatifrons (27,4%), G. undata
(21,6%), y Ch. linearis (21,3%). El mayor porcentaje de adultos de Ch. pinnatifrons
entre tipos de bosque (47%) está de acuerdo con la tendencia reportada
por Svenning (1998) a distribuirse en sitios altamente intervenidos.
El ajuste de la PU a la serie logarítmica (?²=7,86, 4gl, p>0,10),
al igual que en el BNS, podría relacionarse con la alta homogeneidad
estructural encontrada en plantaciones (Murcia 1997), lo que aumenta la disponibilidad
de luz y disminuye la varianza lumínica (Lentijo & Kattan 2005),
permitiendo la dominancia de las especies con mayor capacidad de colonización
(G. undata, con el 62,8% del total de individuos para el tipo de bosque, J´=0,64).
La mayor disponibilidad y/o menor varianza lumínica en relación
con los demás tipos de bosque, podría dar como resultado la mayor
cantidad de individuos encontrada (50,3% del total de individuos muestreados),
así como el mayor porcentaje de plántulas e individuos jóvenes
entre tipos de bosque para Ch. linearis (70,2%), G. undata (69%), Ch. pinnatifrons
(49,8%) y C. alpinum (37,1%), y el alto porcentaje de plántulas e individuos
jóvenes para P. acuminata (29,6%). Además, la acumulación
de hojarasca que se descompone rápidamente podría favorecer la
germinación.
Así mismo, el dendrograma de similitud de Jaccard muestra que el BNS y la PU comparten la mayor cantidad de especies (C. alpinum, G. undata, Ch. pinnatifrons, P. acuminata, W. kalbreyeri y Ch. linearis), lo que indica que los factores que afectan la regeneración de las palmas son similares para los dos tipos de bosque.
Las restricciones edáficas impuestas por la PP (Cavelier & Santos 1999) y roble (Lozano & Torres 1974), podrían disminuir la complejidad de la comunidad de palmas. Además de contener menos especies (S=5 en la PP, S=4 en la PR), se ajustan al modelo de "vara quebrada" (?²=4,34, 3gl, p>0,20 para la PP; ?²=1,15, 3gl, p>0,70 para la PR), indicando que la comunidad es simple y tiene alta equidad (J´=0,85 en robledales, J=0,75 en pineras). Además, contienen la menor cantidad de individuos entre tipos de bosque (5,6% del total de individuos muestreados para la PP y 4,6% para la PR), y la menor cantidad de plántulas e individuos jóvenes entre tipos de bosque para C. alpinum (10,4% para la PP), Ch. linearis (4,2 % para la PP, ausente para la PR), G. undata (3,1% para la PP, 0,8% para la PR), P. acuminata (2,5% para la PR, ausente en la PP), Ch. pinnatifrons (1,1% para la PP) y W. kalbreyeri (ausente en la PR). En la PR la germinación y el reclutamiento solo es alto para C. alpinum y Ch. pinnatifrons (20,8% y 11,2% del total de plántulas e individuos jóvenes entre tipos de bosque) y en la PP para W. kalbreyeri (63,4% del total de plántulas e individuos jóvenes entre tipos de bosque).
Frecuencia y análisis poblacional. La presencia de individuos de G. undata, C. alpinum y Ch. pinnatifrons en todos los tipos de bosque, con las mayores frecuencias de aparición por total de transectos (fr=0,88, 0,72 y 0,72 respectivamente) indica que estas especies tienen un alto potencial para colonizar ambientes distintos, posible resultado de alta producción de semillas, dispersores eficientes, germinación, establecimiento y persistencia en distintas condiciones microambientales. Los tamaños del fruto, de 15 a 23 mm de diámetro para C. alpinum, de 7 a 12 mm de largo por 6 a 12 mm de ancho para Ch. pinnatifrons, y de 7,5 mm de largo por 6,3 mm de ancho para G. undata, posibilita el consumo por parte de aves, y para Ch. pinnatifrons y C. alpinum adicionalmente por mamíferos (Ríos et al. 2004). En bosques montanos del Quindío, Mejía (1999) registró un total de doce dispersores potenciales para C. alpinum, incluyendo pavas (Chamaepetes goudotti), tucanes (Aulacorhynchus haematopygus), murciélagos (Artibeus jamaicensis) y roedores (Dasyprocta fuliginosa). Podría haber un número similar de dispersores para Ch. pinnatrifrons y G. undata, o aún mayor por el tamaño pequeño del fruto. Además, las especies podrían germinar y establecerse en distintos microambientes, aunque de acuerdo con Schwarzkopf (1985, en Ataroff & Schwarzkopf 1992) y Svenning (2001) Ch. pinnatifrons muestra una marcada preferencia por sustratos secos y profundos.
La alteración de las propiedades fisicoquímicas en suelos de
la PP (Cavelier & Santos 1999) y roble (Lozano & Torres 1974) podría
limitar el establecimiento de P. acuminata (fr=0 en la PP, fr=0,2 en la PR).
En cambio está presente en los demás tipos de bosque (Fr=0,56),
lo cual podría considerarse como el resultado del éxito en producción
de semillas, dispersión (Zona & Henderson 1989, como P. montana),
germinación, establecimiento y persistencia en condiciones distintas.
La baja frecuencia de aparición de las dos especies exclusivas a bosques
nativos maduros (G. jussieuana y G. orbignyana, fr=0,04 para las dos especies)
se podría relacionar con especificidades ecológicas para su germinación
y establecimiento, baja producción de semillas y/o dispersión
limitada. A pesar del pequeño tamaño del fruto, de 5 a 8 mm de
largo por 4 a 6 mm de ancho para G. jussieuana (Ríos et al. 2004) y de
9 mm de largo por 8 mm de ancho para G. orbignyana (observación personal),
no se ha reportado la presencia de dispersores para el SFFOQ. Además,
la baja tasa de germinación y crecimiento, y el establecimiento y persistencia
en condiciones de poca luminosidad en sotobosque (Rodríguez-Buriticá
et al. 2005) restringe la habilidad de G. orbignyana para colonizar áreas
de disturbio, lo que probablemente también es cierto para G. jussieuana.
La presencia, y por lo tanto la conservación de estas dos especies, está
ligada a la dinámica en el BNM.
La baja frecuencia de aparición de A. simplex y A. lindeniana (fr=0,16 y 0,04 respectivamente) podría estar relacionada con la baja producción de semillas, poca efectividad dispersora de los animales consumidores, que incluyen mamíferos y aves (Ríos et al. 2004) y/o con especificidades ecológicas que determinan su bajo potencial de reclutamiento. Probablemente, el crecimiento clonal en estas dos especies es una respuesta evolutiva efectiva ante las limitaciones en reclutamiento, lo que también propone Svenning (2001) para la especie clonal A. erinacea.
La ausencia de Ch. linearis y W. kalbreyeri en la PR, podría ser el resultado de la susceptibilidad de las semillas dispersadas a las alteraciones fisicoquímicas del suelo (Lozano & Torres 1974), y/o a la limitación de los rangos de acción de los dispersores potenciales. La presencia de las especies en los demás tipos de bosque (Fr=0,48 para Ch. linearis y 0,44 para W. kalbreyeri) indica que son tolerantes a condiciones microambientales distintas en suelos no alterados.
El mayor porcentaje de plántulas (>58%), seguido por individuos jóvenes (=40%) y adultos (<2%) para G. undata, C. alpinum y P. acuminata se relaciona con una alta mortalidad en estadios de desarrollo tempranos, y por lo tanto con una curva de supervivencia tipo III (Pearl 1927, en Rickleffs & Miller 1999). Esta curva se encontró en rametos de P. acuminata (Gamba-Trimiño 2004), e individuos de C. alpinum (Vergara 2002). Ch. pinnatifrons, Ch. linearis y W. kalbreyeri podrían ajustarse a una curva de supervivencia tipo III, con un porcentaje de adultos menor al 11%. Por su parte G. jussieuana, G. orbignyana, A. simplex y A. lindeniana podrían ajustarse a una curva tipo I o tipo II, con baja mortalidad en estadios de desarrollo tempranos. Una probabilidad de mortalidad distribuida a lo largo de todas las clases de desarrollo, correspondiente a una curva de supervivencia tipo II, fue encontrada por Rodríguez-Buriticá et al. (2005) en el estudio demográfico realizado sobre G. orbignyana. Es necesario aumentar el tamaño muestra para hacer inferencias sobre el comportamiento demográfico de las especies.
AGRADECIMIENTOS
Al personal de la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales, UAESPNN, por la oportunidad de realizar esta investigación en el área del SFFOQ. Al profesor Rodrigo Bernal del Instituto de Ciencias Naturales de la Universidad Nacional, por la valiosa información y material suministrados, y por la ayuda en la determinación taxonómica de G. orbignyana. A los integrantes de la Fundación Ecoandina, a los funcionarios del SFFOQ, a la cooperativa La Suiza, y a la bióloga Lina María González, por su colaboración durante el trabajo de campo.
LITERATURA CITADA
1. ANÓNIMO. 1989-2003. Anuario meteorológico cafetero. Centro
Nacional de Investigaciones de café CENICAFÉ, Chinchiná,
Colombia.
2. ANÓNIMO. 2003. Mapas 1:17000, Propuesta de ordenamiento ambiental
para el Santuario de Fauna y Flora Otún Quimbaya. Unidad Administrativa
Especial del Sistema de Parques Nacionales Naturales.
3. ATTAROF, M. & T. SCHWARZKOPF. 1992. Leaf production, reproductive patterns,
field germination and seedling survival en Chamaedorea bartlingiana, a dioecious
understory palm. Oecologica 92: 250-256.
4. BERNAL, R. 1998. Demography of the vegetable ivory palm Phytelephas seemannii
in Colombia, and the impact of seed harvesting. Journal of Applied Ecology 35:
64-74.
5. BORSCHSENIUS, F., H.B. PEDERSEN & H. BALSLEV. 1998. Manual to the Palms
of Ecuador. Aarhus University Press, Aarhus.
6. BROWN, K.E. 1976. Ecological studies of the Cabbage Palm, Sabal palmetto.
II. Dispersal, predation, and escape of seeds. Principes 20: 49-56.
7. BULLOCK, S.H. 1980. Demography of an undergrowth palm in littoral Cameroon.
Biotropica 12: 247-255.
8. CASWELL, H. 1989. Life-history strategies. Pags. 285-307 en J.M. Cherret
(ed.), Ecological Concepts. Blackwell Scientific Publication, Oxford.
9. CAVELIER, J. & C. SANTOS. 1999. Efectos de plantaciones abandonadas de
especies exóticas y nativas sobre la regeneración natural de un
bosque montano en Colombia. Revista de Biología Tropical 47: 775-784.
10. DRANSFIELD, J. 1986. A guide to collecting palms. Annals of the Missouri
Botanical Garden 73: 166-176.
11. ESPINAL, L.S. & E. MONTENEGRO. 1963. Formaciones vegetales de Colombia.
Memoria explicativa sobre el mapa ecológico. Instituto Geográ?co
Agustín Codazzi IGAC, Bogotá.
12. GAMBA-TRIMIÑO, C. 2004. Demografía de Prestoea acuminata (Palmae)
en el suroccidente colombiano: implicaciones evolutivas y para su manejo. Tesis
de pregrado. Facultad de Estudios Ambientales y Rurales, Pontificia Universidad
Javeriana, Bogotá.
13. GENTRY, A.H. 1995. Patterns of diversity and floristic composition in Neotropical
Montane Forests. Pags. 103-126 en: S. Churchill, H. Balslev, E. Forero &
J.L Luteyn (eds.), Biodiversity and Conservation of Neotropical Montane Forests.
The New York Botanical Garden, Bronx, Nueva York.
14. GLENN-LEWIN, D.C. & E. VAN DER MAAREL. 1992. Patterns and processes
of vegetation dynamics. Pags. 11-59 en: D.C. Glenn, R.K. Peet & T.T. Veblen
(eds.), Plant succession, Theory and prediction. Chapman & Hall, Londres.
15. GUERRERO, L.A, M.A. MORENO, E. GALLEGO, G. MARÍN, R. WALKER, F.E.
OROZCO, O. GARCÍA, G.R. LÓPEZ & M. ZAMORA. 2004. Plan básico
de manejo 2005-2009. Santuario de Fauna y Flora Otún Quimbaya. Parques
Nacionales Naturales de Colombia, Dirección Territorial Noroccidente,
Medellín.
16. HARTSHORN, G.S. 1980. Neotropical forest dynamics. Pags. 23-30 en: J. Ewel
(ed.), Tropical Succession. Suplemento Biotropica 12.
17. HENDERSON, A. 2002. Evolution and ecology of palms. The New York Botanical
Garden Press, Bronx, Nueva York.
18. HENDERSON, A., G. GALEANO & R. BERNAL. 1995. Field guide to the palms
of the Americas. Princeton University Press, Princeton, Nueva Jersey.
19. LENTIJO, G.M, & G.H. KATTAN. 2005. Estratificación vertical de
las aves en una plantación monoespecífica y en bosque nativo en
la Cordillera Central de Colombia. Ornitología Colombiana 3: 51-61.
20. LEPŠ, J. 2005. Diversity and ecosystem function. Pags. 200-237 en:
E. van der Maarel (ed.), Vegetation Ecology. Blackwell publishing, Oxford.
21. LOTT, R.H., G.N. HARRINGTON, A.K. IRVINE & S. MCINTYRE. 1995. Density
- dependent seed predation and plant dispersion of the tropical palm Normanbya
normanbyi. Biotropica 27: 87-95.
22. LOZANO, G., & J.H. TORRES. 1974. Aspectos generales sobre la distribución,
sistemática fitosociológica y clasificación ecológica
de los bosques de roble (Quercus humboldtii) en Colombia. Ecología Tropical
1: 45-78.
23. LUGO, A.E. 1992. Comparison of tropical tree plantations with secondary
forests of similar age. Ecological Monographs 62: 1-41.
24. MAGURRAN, A. 1989. Diversidad Ecológica y su Medición. Ediciones
Vedrá, Barcelona.
25. MEJÍA, G.D. 1999. Dispersión de semillas de la palma de cera
Ceroxylon. alpinum y estado actual de la población de aves en un bosque
montano del departamento del Quindío-Colombia. Trabajo de Grado. Departamento
de Ciencias Biológicas, Facultad de Ciencias, Universidad de Los Andes,
Bogotá.
26. MONTGOMERY, D. C. 2000. Design and Analysis of Experiments. 5a edición.
John Wiley & Sons, Nueva York.
27. MORAES, M., G. GALEANO, R. BERNAL, H. BALSLEV & A. HENDERSON. 1995.
Tropical Andean palms (Arecaceae). Pags. 473-487 en: S. Churchill, H. Balslev,
E. Forero & J.L Luteyn (eds.), Biodiversity and Conservation of Neotropical
Montane Forests. The New York Botanical Garden, Nueva York.
28. MORENO, C.E. 2001. Métodos para medir la biodiversidad. M & T.
Manuales y Tesis SEA, vol,1, Zaragoza.
29. MURCIA, C. 1997. Evaluation of Andean alder as a catalyst for the recovery
of tropical cloud forests in Colombia. Forest Ecology and Management 99: 163-170.
30. NICOTRA, A., R. CHAZDON & S. IRIARTE. 1999. Spatial heterogeneity of
light and woody seedling regeneration in tropical wet forests. Ecology 80: 1908-1926.
31. NOBLE I.R. & R.O. SLATYER. 1980. The use of vital attributes to predict
successional changes in plant communities subject to recurrent disturbances.
Vegetatio 43: 5-21.
32. PEARL, R. 1927. The growth of populations. The Quarterly Review of Biology
2: 532-548.
33. PONCE, M.E., J. BRANDIN, M.A. PONCE, & V. GONZÁLEZ. 1999. Germinación
y establecimiento de plántulas de Mauritia flexuosa L.f (Arecaceae) en
los llanos sur-orientales del estado Guarico, Venezuela. Acta Botánica
Venezuela 22: 167-183.
34. RANGEL-CH., J.O., P.D. LOWY-C & M. AGUILAR-P. 1997. Distribución
de los tipos de vegetación en las regiones naturales de Colombia. Aproximación
inicial. Pags. 383-436 en: J.O. Rangel-Ch., P.D. Lowy-C & M. Aguilar-P (eds.),
Colombia Diversidad Biótica II: Tipos de Vegetación en Colombia.
Universidad Nacional de Coombia, Instituto de Ciencias Naturales, Bogotá.
35. RICKLEFFS, R.E. & G.L. MILLER. 1999. Population Structure. Cap. 14.
Pags. 271-297 en: R.E. Ricklefs & G.L. Miller (eds.). Ecology, Fourth edition.
W.H. Freeman and Company, Nueva York.
36. RÍOS, M., P. GIRALDO & D. CORREA. 2004. Guía de frutos
y semillas de la cuenca media del río Otún. Fundación Ecoandina,
Wildlife Conservation Society, Santiago de Cali.
37. RODRÍGUEZ-BURITICÁ, S., M.A. ORJUELA & G. GALEANO. 2005.
Demography and life history of Geonoma orbignyana: An understory palm used as
foliage in Colombia. Forest Ecology and Management 211: 329-340.
38. SAWYER, J. 1993. Plantations in the tropics. IUCN, Gland.
39. SCHWARSKOPF, T. 1985. Patrones de distribución de plantas del sotobosque
de una selva nublada. Tesis de pregrado. Facultad de Ciencias, Universidad de
los Andes, Mérida.
40. SLOAN, J. 1980. Gap partitioning among tropical rainforest trees. Pags.
47-55 en: J. Ewel (ed.), Tropical Succession. Suplemento Biotropica 12.
41. SVENNING, J.C. 1998. The effect of land-use on the local distribution of
palm species in an Andean rain forest fragment in northwestern Ecuador. Biodiversity
and Conservation 7: 1529-1537.
42. SVENNING, J.C. 2001. Environmental heterogeneity, recruitment limitation
and the mesoscale distribution of palms in a tropical montane rain forest (Maquipicuna,
Ecuador). Journal of Tropical Ecology 17: 97:113.
43. TOMLINSON, R.B. 1990. The structural biology of palms. Claredon Press, Oxford.
44. UHL, N.W. & H.E. MOORE. 1973. The protection of pollen and ovules in
palms. Principes 17: 111-149.
45. VALENCIA, R. 1995. Composition and structure of an Andean forest fragment
in eastern Ecuador. Pags. 239-249 en: S. Churchill, H. Balslev, E. Forero &
J.L Luteyn (eds.), Biodiversity and Conservation of Neotropical Montane Forests.
The New York Botanical Garden, Nueva York.
46. VANDERMEER J.H., J. STOUT & S. RISCH. 1979. Seed dispersal of a common
Costa Rican rainforest palms (Welfia georgii). Journal of Tropical Ecology 20:
17-26.
47. VAN DER VALK, A.G. 1992. Establishment, colonization and persistence. Pags.
60-102 en: D.C. Glenn, R.K. Peet & T.T. Veblen (eds.), Plant succession,
Theory and Prediction. Chapman & Hall, Londres.
48. VERGARA, L.K. 2002. Demografía de Ceroxylon alpinum en bosques relictuales
del valle de Cocora, Salento (Quindío). Tesis de pregrado. Departamento
de Biología, Facultad de Ciencias, Universidad Nacional de Colombia,
Bogotá, D.C.
49. ZONA, S. & A. HENDERSON. 1989. A review of animal-mediated seed dispersal
of palms. Selbyana 11: 6-21.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2009 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















