


Publicado
ARAÑAS ORBITELARES DE UN BOSQUE HÚMEDO SUBTROPICAL DE LA SIERRA NEVADA DE SANTA MARTA (MAGDALENA, COLOMBIA)
Orbweaving spiders of subtropical humid forest of the Sierra Nevada of
Santa Marta (Magdalena, Colombia)
LUIS FERREIRA-OJEDA
EDUARDO FLÓREZ-D.
ALEXANDER SABOGAL-GONZÁLEZ
Grupo de Investigación en Insectos Neotropicales, INTROPIC, Universidad
del Magdalena, Carrera 32 Nº 22-08, Santa Marta, Colombia. ferreluis31@gmail.com
Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Apartado 7495, Bogotá, D.C. Colombia. aeflorezd@unal.edu.co
Grupo de Biotecnología, Centro Internacional de Física, Universidad
Nacional de Colombia. Apartado 7495, Bogotá, D.C. Colombia. asabogalg@yahoo.com
RESUMEN
Se estudió la estructura y la composición de arañas orbitelares (Araneae: Orbiculariae) en un bosque húmedo subtropical de la Sierra Nevada de Santa Marta. Los muestreos se realizaron en cuatro jornadas diurnas y nocturnas, que abarcaron las dos estaciones secas y las dos lluviosas, entre marzo y octubre de 2006. Se emplearon métodos de recolección manual, barrido con red entomológica y agitación de follaje. Se recolectaron 496 arañas orbitelares pertenecientes a seis familias y 35 morfoespecies. Araneidae fue la familia más rica en especies y la más abundante, Nephilidae y Anapidae fueron las menos ricas en especies y Nephilidae la menos abundante. La araneofauna orbitelar mostró preferencia por los microhábitats de vegetación arbustiva y herbácea respectivamente. La araneofauna presentó mayor abundancia diurna con relación a la abundancia nocturna. La composición y estructura de la comunidad de arañas orbitelares parecen estar relacionadas con el régimen climático. La distribución de abundancia de especies se ajustó al modelo de distribución log-normal lo que sugiere que la comunidad de arañas orbitelares es estable y se encuentran en relativo equilibrio.
Palabras clave. Arañas orbitelares, estructura y composición, estacionalidad, preferencia de microhábitats, hábitos de actividad.
ABSTRACT
We studied the structure and composition of orb-weaving spiders (Araneae: Orbiculariae) in a subtropical humid forest of the Sierra Nevada of Santa Marta. The samples were taken during four sampling dates, including night and day, and covering two dry and two rainy seasons, between March and October of 2006. We used manual collection, sweeping net and foliage beating. We collected 496 orb-weaving spiders belonging to six families and 35 morphospecies. Araneidae was the most species-rich and abundant family, Nephilidae and Anapidae showed the least richness and being Nephildae the least abundant. Orb-weaving spiders showed preference for the microhabitats of arbustive vegetation, and herbaceous respectively. The araneofauna of diurnal habits was more abundant than that of nocturnal habits. The composition and structure of the community of the orb-weaving spider fauna seem to be related to the climatic regime. The rank-abundance curve fitted a log-normal pattern, which suggests that the community of orb-weaving spiders is stable and are in relative balance.
Key words. Orb-weaving spiders, structure and composition, seasonality, microhabitats preference, activity habits.
Recibido: 11/03/2008
Aceptado: 09/09/2009
INTRODUCCIÓN
La Sierra Nevada de Santa Marta es la formación montañosa litoral más alta del mundo. Su aguda variación altitudinal y su posición tropical permiten que se encuentre en ella una amplia variedad de climas y ecosistemas que representan casi por completo los encontrados en Colombia y América tropical (Mayr et al. 1997). Esta variedad de climas y ecosistemas genera una amplia diversidad de nichos que favorecen una gran biodiversidad. Sin embargo, los diferentes procesos económicos y sociales que se han dado en la Sierra Nevada, han generado graves efectos ambientales; la tala y quema indiscriminada, la formación de potreros y pastizales para la ganadería, y la expansión de cultivos, han destruido la mayoría de los bosques así como la riqueza y la variedad de flora y fauna nativas presentes en la Sierra Nevada (Mayr et al. 1997).
Las arañas comprenden un grupo faunístico diverso y ampliamente distribuido en todos los ecosistemas terrestres del mundo, ocupando incluso ambientes dulceacuícolas (Turnbull 1973). Son consideradas como uno de los grupos más diversos de la Tierra, ubicándose en el séptimo lugar de órdenes de animales (Coddington & Levi 1991), con 40 998 especies agrupadas en 3 747 géneros y 109 familias (Platnick 2009). Son particularmente diversas en los bosques tropicales, los cuales albergan alrededor del 80 % de la araneofauna desconocida, correspondiendo a la mayor proporción de la araneofauna del planeta, la cual se estima podría llegar a 170 000 especies (Coddington & Levi 1991). A pesar de su alta diversidad, las arañas son unos de los componentes faunísticos menos estudiados en el neotropico y el conocimiento que de ellas se tiene es aún incipiente (Flórez 1996, 1997, 1998, 1999a).
En Colombia han sido registradas 680 especies de arañas agrupadas en 249 géneros y 49 familias (Flórez & Sánchez 1995), de este total de especies cerca del 20 % son endémicas, lo que refleja la alta riqueza de la fauna de arañas que habita en los bosques Colombianos (Flórez 1992). La región Caribe colombiana es considerada una de las más ricas del país en número de especies de arañas (Rangel et al. 1995), sin embargo, es poco el conocimiento de la araneofauna de esta región, los trabajos realizados hasta el momento son: la influencia de las actividades agropecuarias en la diversidad y abundancia de arañas tejedoras en un bosque seco tropical de la Serranía de Pajuancho, Atlántico (Ow 2001) y finalmente la descripción de las familias de arañas presentes en un paleocauce del río Palomino, en La Guajira (Morón 2004).
Teniendo en cuenta lo antes expuesto, este estudio se realizó con el fin de contribuir al conocimiento de la estructura y la composición de arañas orbitelares en un bosque húmedo de la Sierra Nevada de Santa Marta.
MATERIALES Y MÉTODOS
Área de estudio: La Sierra Nevada de Santa Marta, es un macizo montañoso situado en el extremo noroccidental de Suramérica, al norte de la república de Colombia, entre los 10º 01' 05'' y 11º 01' 11'' Norte y entre los 72º 36' 16'' y 74º 12' 49'' Oeste. El régimen de lluvias esta definido en gran parte, por el movimiento de la zona de convergencia intertropical (ZCIT), distinguiéndose en general dos estaciones lluviosas, de abril a junio y de agosto a principios de diciembre, alternadas por dos estaciones secas, de diciembre a marzo y de junio a agosto (veranillo de San Juan) (Mayr et al. 1997, 1998).
Este estudio se realizó en un bosque húmedo de la Sierra Nevada de Santa Marta, según el sistema de formaciones vegetales de Holdridge ajustado por Espinal & Montenegro (1963), el cual se encuentra en los predios de la Hacienda La Victoria, su ubicación es 11º 07' 45'' Norte y 74º 05' 42'' Oeste y tiene una altitud de 900 m.s.n.m.
Fase de campo: Las arañas orbitelares se recolectaron utilizando el método de recolección manual de revisión superior e inferior (looking up and looking down) propuesto por Coddington et al. (1991), examinando exhaustivamente la vegetación y los diferentes microhábitats encontrados (al azar). Este método fue empleado por una sola persona en muestreos diurnos y nocturnos, por espacio de una hora y media cada uno. Complementariamente fueron utilizados otros dos métodos: 1) barrido con red entomológica, el cual consistió en realizar cien pases dobles sobre vegetación herbácea y/o arbustiva, posteriormente se extrajeron los especímenes de la red con un aspirador bucal; y 2) agitación de follaje arbóreo, que consistió en sacudir (veinte golpes con una vara) una rama frondosa o varias ramas de un árbol, colocando previamente una lona de color blanco (0.8 m2) por debajo, para colectar los especímenes que caen con la agitación; este método se realizó en diez árboles por cada jornada de muestreo. Las arañas orbitelares recolectadas fueron separadas en el laboratorio y preservadas en alcohol etílico al 70 % para su posterior identificación.
Se realizaron cuatro jornadas de muestreos teniendo en cuenta las estaciones de la región, las cuales fueron llevadas acabo entre el 31 marzo - 4 abril (seca), entre el 26 - 30 mayo (lluviosa menor), entre el 18 - 22 Julio (veranillo de San Juan) y entre el 19 - 24 septiembre 2006 (lluviosa mayor) respectivamente, de los cinco días dedicados por cada estación, cuatro fueron empleados en recolección manual y uno en barrido con red entomológica y en agitación de follaje arbóreo.
Fase de laboratorio: Las arañas recolectadas fueron diferenciadas en morfoespecies y la identificación taxonómica se realizó utilizando las claves de Levi (1968), Opell (1979), Coddington (1986), Nentwing (1993), Flórez (1996), Dippenaar-Schoeman & Jocqué (1997), Ramírez (1999), Levi (2002) y Aguilera & Casanueva (2005). Posteriormente se confirmó la identificación en la colección aracnológica de referencia del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia. Los especimenes fueron depositados en la colección aracnológica del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia.
Análisis de datos: Se estimó la riqueza de la araneofauna orbitelar a través de la curva de acumulación de especies propuesta por Jiménez-Valverde & Hortal (2003), utilizando los programas EstimateS versión 7.5 (Colwell 2005) y STATISTICA versión 6 (Statsoft 2001).
La estructura y la composición de arañas orbitelares se determinó a través del índice de Simpson (Dominancia), e índice de Shannon Wiener (Equidad). La similitud entre estaciones se determinó a través del índice de Similitud Bray-Curtís, para obtener dichos índices se utilizó el programa BioDiversity Professional versión 2 (McAleece 1997).
Se describió la estructura de la comunidad de arañas orbitelares en términos de la distribución de abundancia de especies, determinándose a cual de los modelos (serie geométrica, serie logarítmica y distribución log-normal) se ajusta dicha comunidad, con el programa PAST (Paleontological Statistics) versión 1.12 (Hammer et al. 2003)
RESULTADOS
Se recolectaron 496 arañas orbitelares, agrupadas en seis familias, 24 géneros y 35 morfoespecies (Tabla 1). Araneidae fue la familia más rica en especies con 22 morfoespecies y la más abundante con 348 individuos, que corresponden al 70.16 % del total de individuos colectados, seguida por Tetragnathidae con seis morfoespecies y 76 individuos, mientras que Nephilidae y Anapidae fueron la menos ricas en especies con una morfoespecie cada una y Nephilidae con tres individuos fue la menos abundante.
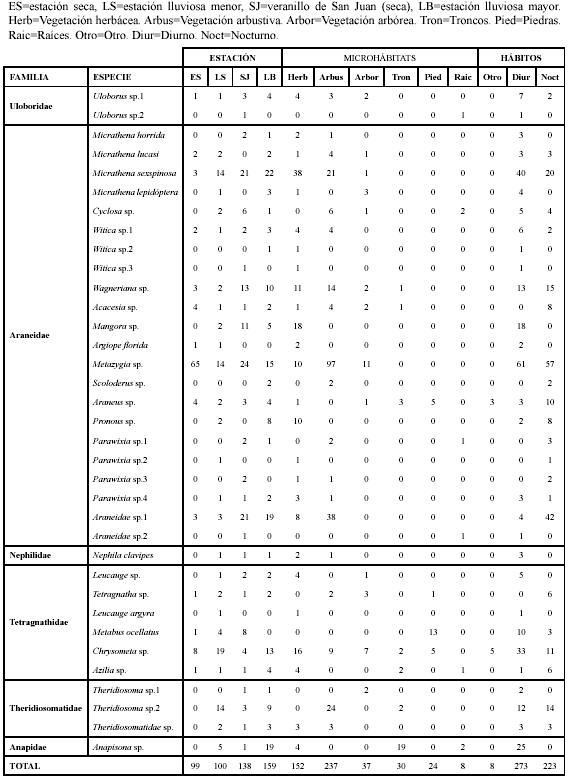
Micrathena y Parawixia fueron los géneros más ricos en especies, cada uno con cuatro morfoespecies, la morfoespecie con mayor abundancia fue Metazygia sp. con 118 individuos (23.79 %) seguida por Micrathena sexspinosa con 60 individuos (12.09 %), mientras que Uloborus sp.2, Witica sp.2, Witica sp.3, Parawixia sp.2, Araneidae sp.2 y Leucauge argyra fueron las morfoespecies que presentaron la menor abundancia cada una con un solo individuo (0.2 %).
Estimación de riqueza
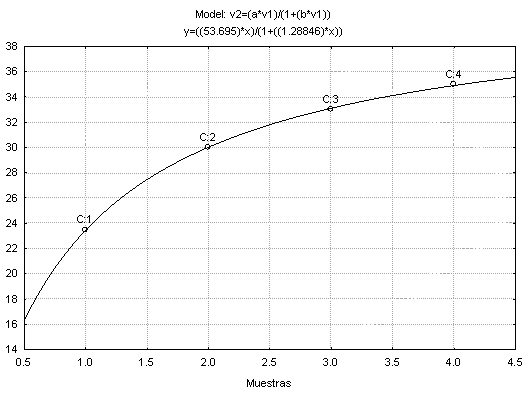
La curva de acumulación de especies (Fig. 1) permite deducir que el inventario es completo, debido a que se recolectó el 83.98 % de la araneofauna que puede albergarse en un bosque húmedo subtropical de la Sierra Nevada. Sin embargo, sí se considera el valor de la pendiente de la curva se puede inferir que el inventario es aún insuficiente y debe ser tomado con prudencia si se intenta obtener conclusiones sobre la riqueza del lugar (Jiménez-Valverde & Hortal 2003).
Araneofauna orbitelar y su relación con las estaciones
En la estación seca se registraron sólo tres familias, 14 morfoespecies y 99 individuos. En las demás estaciones se recolectaron las seis familias; durante la estación lluviosa menor fueron registradas 26 morfoespecies y 100 individuos, en el veranillo de San Juan 27 morfoespecies y 138 individuos, y durante la estación lluviosa mayor 27 morfoespecies y 159 individuos (Tabla 1).
Preferencia de microhábitats de la araneofauna orbitelar
La araneofauna orbitelar presentó mayor abundancia en los microhábitats de vegetación arbustiva, herbácea y arbórea respectivamente (Tabla 1). Dentro de las arañas orbitelares recolectadas Uloborus sp.2, Witica sp.2, Witica sp.3, Mangora sp., Argiope florida (Chamberlin & Ivie 1944), Scoloderus sp., Pronous sp., Parawixia sp.2, Araneidae sp.2, Leucauge argyra (Walckenaer 1842), Metabus ocellatus (Keyserling 1864) y Theridiosoma sp.1 fueron morfoespecies que se registraron en un microhábitat, mientras que Araneus sp. fue registrada en cinco microhábitats y Chrysometa sp. fue registrada en seis microhábitats. Las morfoespecies restantes de arañas orbitelares fueron registradas en dos, tres o cuatro de los microhábitats (Tabla 1).
Hábitos de actividad de la araneofauna orbitelar
Las familias Nephilidae y Anapidae sólo fueron registradas en muestreos diurnos, mientras que las familias Uloboridae, Araneidae, Tetragnathidae y Theridiosomatidae en ambos tipos de muestreo. Se registraron trece morfoespecies con presencia exclusivamente diurna Uloborus sp.2, Micrathena horrida (Taczanowski 1873), M. lepidóptera (Mello-Leitão 1941), Witica sp.2, Witica sp.3, Mangora sp., Argiope florida, Araneidae sp.2, Nephila clavipes (Linnaeus 1767), Leucauge sp., Leucauge argyra, Theridiosoma sp.1 y Anapisona sp., seis morfoespecies con presencia exclusivamente nocturna Acacesia sp., Scoloderus sp., Parawixia sp.1, Parawixia sp.2, Parawixia sp.3 y Tetragnatha sp., y 16 morfoespecies con presencia tanto diurna como nocturna. En términos generales la araneofauna presentó mayor abundancia diurna con relación a la abundancia nocturna (Tabla 1).
Estructura y composición
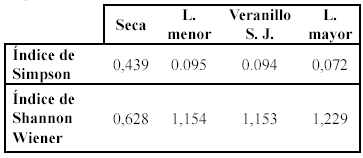
Los valores de dominancia y equidad de la araneofauna orbitelar recolectada en un bosque húmedo subtropical de la Sierra Nevada según las estaciones se presentan en la tabla 2.
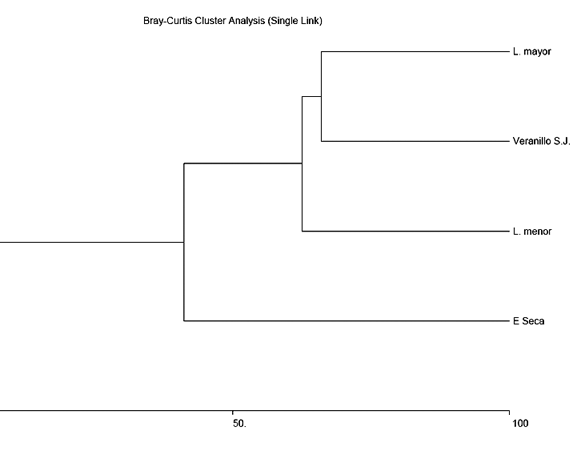
El índice de similitud de Bray-Curtis entre estaciones mostró mayor similitud entre la estación lluviosa menor, el veranillo de San Juan y la estación lluviosa mayor, mientras que la estación seca presentó la menor similitud (Fig. 2). La probabilidad de ajuste a los modelos de distribución de abundancia de especies fueron 1.337e-10 para la serie geométrica, 0.6076 para la serie logarítmica y 0.8878 para la distribución log-normal.
DISCUSIÓN
La familia Araneidae se registró como la más rica en especies dentro de la araneofauna orbitelar recolectada en un bosque húmedo subtropical de la Sierra Nevada, esto concuerda con lo registrado por Silva (1996) y Silva & Coddington (1996) en el neótropico. En Colombia coincide con lo reportado por Barriga (1995), Bello (1995), Flórez & Sánchez (1995), Flórez (1996, 1998, 1999b), Vallejo (1997) y Ow (2001).
Araneofauna orbitelar y su relación con las estaciones
La riqueza y la abundancia de arañas orbitelares aumentó a lo
largo del año (Tabla 1), similar al aumento de
niveles de precipitación, estos resultados coinciden con los obtenidos
por Barriga (1995) y Niño et al. (2002); sin embargo, difieren de los
resultados obtenidos por Bello (1995), Blanco-Vargas et al. (2003) y Ow (2001).
Los resultados obtenidos en el presente estudio podrían atribuirse a
que el rango y la frecuencia en el número de individuos de una comunidad
de arañas puede aumentar o disminuir con los periodos de lluvia (Döbel
et al. 1990 en: Niño et al. 2002), igualmente se conoce que otros factores
como, la temperatura y la humedad, determinan la abundancia de las poblaciones
de arañas (Flórez 1997) y en la Sierra Nevada la humedad relativa
muestra un comportamiento estrechamente relacionado con el régimen de
lluvias (Mayr et al. 1997, 1998). Por otra parte estudios fenológicos
de la artropodofauna tropical demuestran cambios en las densidades poblacionales
regulados por las variaciones interanuales de lluvia y sequía (Willis
1976, Buskirk & Buskirk 1976, Levings & Windsor 1985, Pearson &
Derr 1986).
Preferencia de microhábitats de la araneofauna orbitelar
La araneofauna orbitelar mostró mayor abundancia en los microhábitats de vegetación arbustiva y herbácea (Tabla 1). Resultados similares han sido registrados por Flórez (1996, 1997, 1998, 1999a, 1999b), Bello (1995), Vallejo (1997), Niño et al. (2002), y Blanco-Vargas et al. (2003), sin embargo, difieren de los hallados por Rico et al. (2005) que registran mayor abundancia en el microhábitat de vegetación alta para la araneofauna del Parque Nacional Natural Isla Gorgona. La preferencia de microhábitats registrada probablemente se debió a que algunas especies de arañas presentan preferencia por hábitats determinados en el establecimiento de sus telas (Hatley & Macmahon 1980, Robinson 1981, Provencher & Vickery 1988, Gunnarson 1990 en: Blanco-Vargas et al. 2003). También a que la presencia de ciertas formas de plantas como soportes de las telas es importante en determinar la distribución de algunas especies de arañas (Fraser & Frankie 1986 en: Niño et al. 2002) así como la altura sobre el suelo muestra influencia en el sitio donde se construye la tela. Además es probable que la vegetación arbustiva presente mayor heterogeneidad espacial con respecto a los otros microhábitats, lo cual podría favorecer el establecimiento de mayor número de telas en dicho microhábitat.
Estructura y composición
Se registró la mayor dominancia (índice de Simpson) durante estación
seca y lluviosa menor respectivamente (Tabla 2). Estos
valores fueron debidos en gran parte a la alta abundancia de la morfoespecie
Metazygia sp. (morfoespecie más abundante en el presente estudio) en
dichas estaciones, debido a que este índice esta fuertemente influenciado
por la importancia de las especies más abundantes (Magurran 1988). Al
analizar los valores de equidad (Índice de Shannon Wiener) se observa
una tendencia contraria a la dominancia debido a que la equidad es un parámetro
inverso a la dominancia de la comunidad (Moreno 2001). Se registraron estos
resultados porque el régimen climático parece estar asociado con
variaciones en la riqueza de especies de arañas, así como en la
composición y estructura de las comunidades de arañas orbitelares.
La similitud registrada entre estaciones (Fig. 2) estaría debida a los
niveles de precipitación a lo largo del año, lo cual puede proporcionar
las condiciones donde se ven favorecidas ciertas especies de arañas,
considerando que el gradiente de factores ambientales es un limitante en la
abundancia y riqueza de especies, debido a que influye de manera diferente en
cada especie y las poblaciones reúnen una serie de características
que les confieren la capacidad de adaptación a estos cambios (Matteuci
& Colma 1982 en: Barriga 1995).
La distribución de abundancia de especies se ajustó al modelo de distribución log-normal, similarmente a lo registrado por Barriga (1995), Bello (1995) en uno de los hábitat considerados (pastizal) y Cepeda & Flórez (2006), probablemente porque las comunidades de arañas orbitelares presentes en un bosque húmedo subtropical de la Sierra Nevada son estables y se encuentran en relativo equilibrio (Moreno 2001) y además la mayoría de los estudios ecológicos parecen ajustarse a el modelo de distribución de abundancia de especies log-normal (May 1975, Sugihara 1980, Gaston & Blackburn 2000, Longino et al. 2002 en: Magurran 2004).
AGRADECIMIENTOS
Este trabajo fue posible gracias al programa de Biología de la Universidad del Magdalena (Santa Marta) y al Instituto de Ciencias Naturales de la Universidad Nacional de Colombia (Bogotá). Agradecemos a Micky Weber por permitir llevar acabo esta investigación dentro de los predios de la Hacienda La Victoria y al Grupo de Investigación en Insectos Neotropicales, Universidad del Magdalena.
LITERATURA CITADA
1. AGUILERA, M.A. & M.E. CASANUEVA. 2005. Arañas chilenas: estado
actual del conocimiento y clave para las familias de Araneomorphae. Gayana 69(2):201-224.
2. BARRIGA, J.C. 1995. Cambios en la diversidad de arañas constructoras
de telas orbiculares (Araneae: Orbicularie) a lo largo de un gradiente altitudinal,
en el Parque Nacional de Munchique, Cauca. Trabajo de grado, Pontificia Universidad
Javeriana, Bogotá, D.C.
3. BELLO, J.C. 1995. Efectos de borde sobre la distribución de arañas
orbitelares (Aranae: Orbiculariae), en un bosque de niebla de la Reserva Natural
La Planada, Nariño. Trabajo de grado, Pontificia Universidad Javeriana,
Bogotá, D.C.
4. BLANCO-VARGAS, E., G. AMAT-GARCÍA & E. FLÓREZ-DAZA. 2003.
Araneofauna orbitelar (Araneae: Orbiculariae) de los Andes de Colombia: Comunidades
en hábitats bajo regeneración. Revista Ibérica de Aracnología
7:189-203.
5. BUSKIRK, R.E. & W.H. BUSKIRK. 1976. Changes in arthropod abundance in
highland Costa Rican forest. Amer. Midl. Nat. 95(2):288-298.
6. CEPEDA, J. & E. FLÓREZ. 2006. Arañas tejedoras: uso de
diferentes microhábitats en un bosque andino (3131 msnm) de Colombia.
Revista Ibérica de Aracnología 14:39-48.
7. CODDINGTON, J.A. 1986. The genera of spider family Theridiosomatidae. Smiths.
Contr. Zool. 442:1-96.
8. CODDINGTON, J. & H. LEVI. 1991. Systematic and evolution of spiders (Araneae).
Ann. Rev. Ecol. Syst. 22:565-592.
9. COLWELL, R.K. 2005. EstimateS: Statistical estimation of species richness
and shared species from samples. Version 7.5. User's Guide and application published
at: http://purl.oclc.org/estimates.
10. DIPPENAAR-SCHOEMAN, A.S. & R. JOCQUÉ. 1997. African spiders an
identification manual. Agricultural Research Council. South Africa.
11. DÖBEL, H., R. DENNO & J. CODDINGTON. 1990. Spider (Araenae) Community
structure in a intertidial salt marsh: Efects of vegetation structure and tidal
flooding. Environ. Entomol. 19(2):1356-1370
12. ESPINAL, L.S. & E. MONTENEGRO. 1963. Formaciones vegetales de Colombia.
Instituto Geográfico Agustín Codazzi. Bogotá, D.C.
13. FLÓREZ, E. 1992. Las arañas de Colombia. Aspectos históricos
y estado actual de su conocimiento. Cespedesia 19(62-63):239-241.
14. FLÓREZ, E. 1996. Las arañas del departamento del Valle. Un
manual introductorio a su diversidad y clasificación. Inciva-Colciencias.
Santiago de Cali.
15. FLÓREZ, E. 1997. Estudio de la comunidad de arañas del bosque
seco tropical de la estación biológica "El Vinculo".
Cespedesia 22(69):37-57.
16. FLÓREZ, E. 1998. Estructura de comunidades de arañas (Araneae)
en el departamento del Valle, Suroccidente de Colombia. Caldasia 20(2):173-192.
17. FLÓREZ, E. 1999a. Estructura y composición de una comunidad
de arañas (Araneae) en un bosque muy seco tropical de Colombia. Bol.
Entomol. Venez. 14(1):37-51.
18. FLÓREZ, E. 1999b. Estudio de comunidades de arañas (Araneae)
del Parque Nacional Farallones de Cali, Colombia. Cespedesia 23(73-74):99-113.
19. FLÓREZ, E. & H. SÁNCHEZ. 1995. Diversidad de los arácnidos
en Colombia. Aproximación inicial. Págs 327-372. En: Rangel, J.
O. Colombia diversidad biótica I. Guadalupe LTDA. Bogotá, D.C.
20. FRASER, J.B. & G.W. FRANKIE. 1986. An ecological comparison of spiders
from urban and natural habitats in California. Hilgardia 54:1-25.
21. GASTON, K. J. & T.M. BLACKBURRN. 2000. Macroecology. Oxford, UK: Blackwell
Science.
22. GUNNARSON, B. 1990. Vegetation structure and the abundance and size distribution
of spruce-living spiders. J. Anim. Ecol. 59: 743-752.
23. HAMMER, O., D.A.T. HARPER & P.D. RYAN. 2003. PAST: Paleontological Statistics.
Version 1.12
24. HATLEY, C. & J. MACMAHOM. 1980. Spider community organization: Seasonal
variation and the role of vegetation architecture. Environ. Entomol. 9:632-639.
25. JIMÉNEZ-VALVERDE, A. & J. HORTAL. 2003. Las curvas de acumulación
de especies y la necesidad de evaluar la calidad de los inventarios biológicos.
Revista Ibérica de Aracnología 8:151-161.
26. LEVI, H. 1968. The spider genera Gea and Argiope in America (Aranea: Araneidae).
Bull. Mus. Comp. Zool. 136(9):319-352.
27. LEVI, H. 2002. Keys to the genera of Araneid orbweavers (Araneae, Araneidae)
of the Americas. J. Arachnol. 30:527-562.
28. LEVINGS, S.C. & D.M. WINDSOR. 1985. Litter arthropod populations in
a tropical deciduous forest: relationships between years and groups. J. Anim.
Ecol. 54:6-169.
29. LONGINO, J.T., J. CODDINTON & R.K. Colwell. 2002. The ant fauna of a
tropical rain forest: estimating species richness three different ways. Ecology.
83:689-702.
30. MAGURRAN, A.E. 1988. Ecological diversity and its measurement. Princeton
University Press. Nueva Jersey. 179 p.
31. MAGURRAN, A.E. 2004. Measuring Biological Diversity. Blackwell Science Ltd.
32. MATEUCCI, S.D. & S. COLMA. 1982. Metodología para el estudio
de la vegetación. Secretaría general de la OEA. Programa Regional
de Desarrollo Científico y Tecnológico. Washington D.C.
33. MAY, R.M. 1975. Patterns of species abundance and diversity. Págs
81-120. In: Cody, M. L & J. M. Diamond (ed). Ecology and evolution of communities.
Cambridge M. A. Harvard University Press.
34. MAYR, J., H. SÁNCHEZ, P. APREZA, M. CASTILLA, M.C. SUAZA, G.E. RODRÍGUEZ
& C.A. AYALA. 1997. Plan de desarrollo sostenible de la Sierra nevada de
Santa Marta. Fundación Pro-Sierra Nevada de Santa Marta. Santa Marta.
35. MAYR, J., M. C. DÍAZ-GRANADOS, M.C. SUAZA, G.E. RODRÍGUEZ,
F. SALAZAR, E. CARBONÓ, D. RODRÍGUEZ, O. MENDIOLA & M. CASTILLA.
1998. Evaluación ecológica rápida. Definición de
áreas criticas para la conservación en la Sierra Nevada de Santa
Marta. Fundación Pro-Sierra Nevada de Santa Marta. Santa Marta.
36. MC ALEECE, N. 1997. Biodiversity Professional Beta 2.0. The Natural History
Museum y The Scottis Association for Marine Science.
37. MORENO, C.E. 2001. Métodos para medir la biodiversidad. M y T-Manuales
y Tesis SEA, Vol. 1. Zaragoza. 84 p.
38. MORÓN, E. 2004. Descripción de las familias de arañas
(Arácnida: Aranae) en un paleocauce del río Palomino, Dibulla-Guajira.
Trabajo de grado, Universidad del Magdalena, Santa Marta. D.T.C.H.
39. NENTWING, W. 1993. Spiders of Panama. Flora y fauna Handbook No. 12. Sandihill
Crane Press, Inc.
40. NIÑO, D., A. MARTÍNEZ & G. MORA. 2002. Estudio de las
comunidades de arañas (Arácnida: Aranae) de dos ecosistemas en
la región subxerofítica de la Vereda Mosquera (Cundinamarca).
Trabajo de grado, Universidad Distrital Francisco José de Caldas, Bogotá,
D.C.
41. OPELL, B.D. 1979. Revision of genera and tropical American species of the
spider family Uloboridae. Bull. Mus. Comp. Zool. 148(10):443-549.
42. OW, Y. 2001. Influencia de las actividades agropecuarias en la diversidad
y abundancia de arañas tejedoras en un bosque seco tropical en la Serranía
de Pajuancho, Municipio de Juan de Acosta, Atlántico. Trabajo de grado,
Universidad del Atlántico, Barranquilla.
43. PEARSON, D.L. & J.A. DERR. 1986. Seasonal patterns of lowland forest
floor arthropod abundance in Southeastern Peru. Biotropica 18:244-256.
44. PLATNICK, N.I. 2009. The world spider catalog. Version 10.0. The American
Museum of Natural History. Disponible en Internet: http://research.amnh.org/entomology/spiders/catalog/COUNTS.html
(Citado 14 Octubre 2009).
45. RANGEL-CH., J.O., P. LOWY & H. SÁNCHEZ. 1995. Región Caribe.
Págs 217-232. En: Rangel, J. O. Colombia Diversidad Biótica I.
Instituto de Ciencias Naturales-Universidad Nacional de Colombia. Bogotá,
D.C.
46. RAMÍREZ, M.J. 1999. Araneae. Págs 39-59. En: Crespo, F. A.,
M. S. Iglesias y A. C. Valverde. El ABC en la determinación de artrópodos
I. CCC Educando.
47. RICO, A., J. P. BELTRÁN, J. A. ÁLVAREZ, & E. FLÓREZ.
2005. Diversidad de arañas (Arachnida: Araneae) en el Parque Nacional
Natural Isla Gorgona, Pacífico colombiano. Biota Neotropica 5(1a). Disponible
en Internet: http://www.biotaneotropica.org.br/v5n1a/pt/abstract?inventory+BN007051a2005.
48. ROBINSON, J. 1981. The effect of architectural variation in habitat on spider
community: An experimental field study. Ecology 62:73-80.
49. SILVA, D. 1996. Species composition and community structure of Peruvian
rainforest spiders: A case study from seasonally inundated forest along the
Samira river. Rev. Suis. Zool., vol. hors serie: 597-610.
50. SILVA, D. & J.A. CODDINGTON. 1996. Spiders of Pakitza (Madre de Dios)
Peru: Species richness and notes on community structure. Págs 241-299.
En: Wilson, D. y A. Sandoval (eds). The biodiversity of Pakitza and its environs.
Smithsonian Institution, Washington, D.C.
51. STATSOFT. 2001. STATISTICA (data analysis software system and computer program
manual). Version 6. StatSoft, Inc., Tulsa.
52. SUGIHARA, G. 1980. Minimal Community structure: an explanation of species
abundance patterns. Am. Nat. 116:770-787.
53. TURNBULL, A.L. 1973. Ecology of the true spiders (Araenomorpha). Ann. Rev.
Entomol. 18:305-348.
54. VALLEJO, M. 1997. Estructura de una comunidad de arenas orbitelares (Arácnida:
Aranae en sistemas agroforestales del bajo Anchicayá, Pacifico colombiano.
Trabajo de grado, Universidad del Valle, Cali.
55. WILLIS, E.O. 1976. Seasonal changes in the invertebrate. Rev. Bras. Biol.
36:643-657.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2009 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















