


Publicado
MACROALGALES ASOCIADAS AL LITORAL ROCOSO DEL DEPARTAMENTO DE CÓRDOBA, COLOMBIA
Structure of the macroalgal communities associated with the rocky coastline of the department of Córdoba, Colombia
JORGE A. QUIRÓS-RODRÍGUEZ
JORGE E. ARIAS-RÍOS
ROSALBA RUIZ VEGA
Programa de Biología, Universidad de Córdoba, Montería,
Colombia. scaride@yahoo.com, ariosjor@hotmail.com,
rruiz@sinu.unicordoba.edu.co
RESUMEN
El comportamiento ecológico de las comunidades macroalgales en el Caribe cordobés es desconocido, por lo tanto para determinar la diversidad, la abundancia y la distribución de las especies se evaluó la estructura de las comunidades macrolagales asociadas a litoral rocoso del departamento de Córdoba durante los meses de mayo de 2004 hasta octubre de 2005, estableciendo siete puntos de muestreo; cuatro en la ecorregión Morrosquillo y tres en la ecorregión Darién. Para establecer la cobertura de las macroalgas, se utilizó un cuadrante de 625 cm2 con cinco repeticiones dispuestas al azar en cada punto de muestreo. Se reportaron 50 especies de algas bentónicas, de las cuales 27 son rodofitas, 17 clorofitas y 12 feofitas; de las 20 familias encontradas, Dictyotaceae presentó el mayor número de especies (10), seguida por Gracilariaceae (7) y Rhodomelaceae (7) y Halymeniaceae (4). Los géneros con mayor número de especies fueron Gracilaria (6), Padina (5) y Caulerpa (4). La composición de las comunidades macroalgales asociadas al sector comprendido entre Punta Bolívar y Punta Mestizos estuvo integrada por tres divisiones, siendo las rodofitas las que presentaron mayor porcentaje de cobertura (78%), seguida por las clorofitas (14%) y finalmente las feofitas (9%). En el sector que comprende el litoral que va desde Punta Broqueles hasta Puerto Escondido, las rodofitas mostraron el mayor porcentaje de cobertura (62%), seguida por las feofitas (23%) y finalmente las clorofitas (15%). El análisis de clasificación (UPGMA), registró un coeficiente de afinidad del 57%, indicando que la estructura y la composición de las comunidades algales asociadas se comportaron de manera similar en las dos ecorregiones; este comportamiento mostró la conformación de unidades estables en cada punto de muestreo, manifestando cierta homogeneidad espacial, lo cual puede estar relacionado con el tipo de sustrato y las perturbaciones ambientales propias de cada estación.
Palabras clave. Comunidades macroalgales, diversidad, litoral rocoso, riqueza.
ABSTRACT
The ecological behavior of macroalgal communities from the department of Córdoba, in the Colombian Caribbean is unknown. To determine diversity, abundance and distribution of species, the structure of the macroalgal communities distributed throughout the coastline of the department of Cordoba was evaluated in seven stations during May 2004 to October 2005; four located in the Morrosquillo ecoregion and three in the Darién ecoregion. To establish macroalgal cover, in each station a grid of 625 cm2 was used to collect the species during five random repetitions. A total of 50 species of associated algae were reported, 27% of which are red algae, 17% green algae and 12% brown algae; of the 20 families were found, Dictyotaceae presented the highest number of species (10), followed by Gracilariaceae (7) Rhodomelaceae (7), and Halymeniaceae (4). The taxa with the high number of species were Gracilaria (6) Padina (5), and Caulerpa (4). The composition of the macroalgal communities associated to the sector between Punta Bolivar and Punta Mestizos was integrated by three divisions with the red algae exhibiting the highest percentage of cover (78%), followed by the green algae (14%) and finally the brown algae (9%). In the sector between Punta Broqueles and Puerto Escondido, the red algae showed the highest percentage of cover (62%), followed by the brown algae (23%) and finally the green algae (15%). The classification analyses (UPGMA) showed an affinity coefficient of 57%, indicating that the structure and composition of associated macroalgal communities a similar between the two ecoregions. This behavior showed the conformation of stable units in each station suggesting a certain spatial homogeneity which can be related with the kind of substrate and environmental changes of each station.
Key words. Macroalgal communities, diversity, rocky coastline, richness.
Recibido: 31/03/2009
Aceptado:10/09/2010
INTRODUCCIÓN
Las macroalgas bénticas son un componente fundamental de los ecosistemas marinos, debido a que constituyen el principal alimento de peces, moluscos, crustáceos y equinodermos pastoreadores, que al parecer son uno de los factores determinantes en la distribución de las comunidades macroalgales (Reinthal y Macintyre, 1982; Hay, 1984; Lewis, 1985; Littler et al., 1987). El hecho de presentar ciclos de vida cortos y rápidas respuestas a perturbaciones resalta la importancia de estas comunidades permitiendo detectar alteraciones ambientales (Frithsen y Holland, 1992), además son una fuente de material orgánico tanto empleado por las bacterias heterotróficas, como particulado, el cual proviene de la abrasión de talo por efecto del oleaje (Klumpp y Mckinnon, 1992).
La estructura de las comunidades algales en el Caribe ha sido estudiada por autores como Van den Hoeck et al. (1975 y 1978), Wander (1976) y Bak (1976), quienes realizaron investigaciones en Curazao; Littler et al. (1987), realizó trabajos en Belize; asimismo, Adey y Steneck (1995) y Morrison (1988), investigaron sobre las algas en St. Croix en Jamaica. Para el Caribe colombiano los estudios sobre la estructura y ecología de las macroalgas se han concentrado en cinco sectores principales: el área de Santa Marta, desde Ciénaga Grande de Santa Marta hasta el límite oriental del Parque Nacional Natural Tayrona (Bula-Meyer, 1987; Garzón-Ferreira y Cano, 1991; Díaz-Pulido y Garzón-Ferreira, 2002; García y Díaz-Pulido, 2006); Cartagena, incluyendo la costa de la ciudad, Islas del Rosario, Barú y el Bajo de Salmedina (Ángel y Quirós, 1971; Bula-Meyer et al., 1986); el noreste chocoano, entre Acandí y Cabo Tiburón (Bula-Meyer y Schnetter, 1988); el noreste de La Guajira desde Riohacha hasta Castilletes (Bula-Meyer, 1987) y el archipiélago de San Andrés y Providencia (Schnetter, 1976 y 1978); sin embargo, para el litoral Caribe del departamento de Córdoba demarcado entre los 9º 25´N y 75º 42´W en el corregimiento de El Porvenir y los 8º 54´N y 76º 26´W en Punta Arboletes, no se conoce información para este sitio, de tal forma que las especies registradas hacen parte de inventarios parciales referidos al Caribe colombiano en general.
Dada la ubicación del litoral cordobés, éste constituye un sistema formado por ciénagas, estuarios, quebradas, islas e islotes que definen una estructura macroalgal característica para el área, por lo que se hace importante la realización de trabajos que permitan evaluar el comportamiento de las comunidades macroalgales asociadas al litoral del departamento de Córdoba. Para conocer la dinámica de las comunidades de macroalgas en este punto geográfico se propone responder las siguientes preguntas:
1. ¿Cuál es la diversidad de la comunidad macroalgal presente
en el Caribe cordobés?
2. ¿Cuál es la abundancia de las especies macroalgales presentes
en la zona intermareal rocosa entre 0 y 1 m de profundidad?
3. ¿Cómo varía la abundancia de las especies macroalgales
en las dos ecorregiones de estudio y en los meses de muestreo?
METODOLOGÍA
Área de estudio
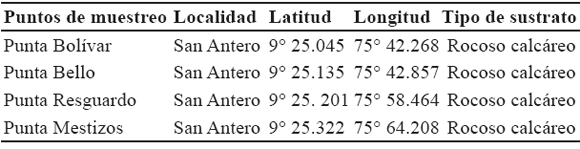
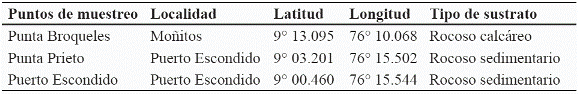
El estudio se ubicó en el litoral Caribe del departamento de Córdoba entre los 9º 25´N y 75º 42´ W en el corregimiento de El Porvenir y los 8º 54´N y 76º 26´W en Punta Arboletes (Figura 1). El litoral costero cordobés en general corresponde a una zona de bosque seco tropical, según el patrón de zonación de Holdrige (1978); además presenta dos épocas climáticas al año, una seca que abarca los meses de diciembre hasta inicios de abril, y otra lluviosa, la cual comprende finales de abril hasta noviembre. En julio se presenta una disminución de las lluvias, la cual es llamada "Veranillo de San Juan". La extensión de la franja costera de Córdoba es de 140 km y recibe la influencia de dos ecorregiones con características propias en cuanto a su geología, hidrología, flora y fauna (Invemar-Ministerio del Medio Ambiente, 2000), situación que permitió dividir el litoral en dos área geográficas (Tabla 1). El sector correspondiente al municipio de San Bernardo del Viento, no presenta una zona intermareal rocosa consolidada, por lo que en su área no se establecieron puntos de muestreo.
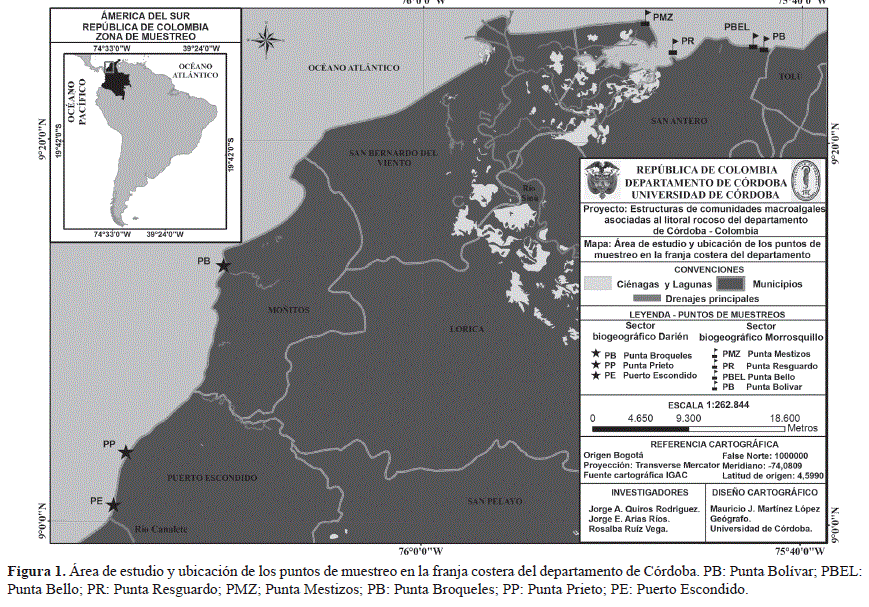
Figura 1. Área de estudio y ubicación de los puntos de muestreo en la franja costera del departamento de Córdoba. PB: Punta Bolívar; PBEL: Punta Bello; PR: Punta Resguardo; PMZ; Punta Mestizos; PB: Punta Broqueles; PP: Punta Prieto; PE: Puerto Escondido.

Ecorregión Morrosquillo. En Córdoba la ecorregión
se encuentra definida por el extremo sur del gofo de Morrosquillo y está
formada por depósitos fluviales de la formación magdalena y por
sedimentación aluvial reciente. El área geográfica corresponde
al litoral costero cordobés que se extiende desde El Porvenir (La Española)
hasta Punta Mestizos y está caracterizado por la formación de
terrazas coralinas y elevaciones del terreno por acción volcánica
(Patiño y Flórez, 1993); esta condición en el sector, brinda
disponibilidad de sustrato, importante para el establecimiento de gran variedad
de algas bentónicas e invertebrados marinos (Quirós, 2009). Asimismo,
el sector está influenciado por el aporte de aguas continentales, siendo
la más importante la del río Sinú hacia el sur de Playa
Blanca (Vernette, 1985).
Ecorregión Darién. Esta ecorregión se proyecta en Córdoba desde el municipio de Los Córdobas hasta San Bernardo del Viento. El área geográfica a nivel del litoral, presenta una costa rocosa interrumpida regularmente por áreas de playa y recibe la influencia de corrientes de agua de origen continental que drenan la costa, tales como el río Canalete, río Córdoba y quebrada Broqueles, aumentando la carga de sedimentos lo cual hace mayor la turbidez (CVS, 2004); en los aspectos geológicos, el sector está formado por acantilados de origen sedimentario que afloran al norte del departamento de Córdoba y son importantes como sustrato para el desarrollo algal. También se observa hacia algunos sectores material arrecifal fósil (Rangel-Buitrago y Posada-Posada, 2005).
FASE DE CAMPO
Ubicación de los Puntos de muestreo
Se realizaron muestreos mensualmente entre junio de 2004 y mayo de 2005, con el fin de abarcar los dos periodos climáticos del año que se dan en los sectores de estudio. En cada punto de muestreo seleccionado se establecieron las coordenadas geográficas mediante un geoposicionador (GPS), Garmin Etrex Legend (Tabla 2, tabla 3). Para la selección de los puntos de muestreo en cada área geográfica se tuvieron en cuenta varios criterios como el tipo de sustrato dominado por algas, grado de intervención antrópica, aporte de aguas continentales y vías de acceso.
Diseño de muestreo
En cada punto de muestreo se ubicó un cinturón de cuadrantes,
para la cual se colocó cinco veces un marco de 625 cm2 de acuerdo a lo
establecido por Connor y Adey (1977), distribuidos en siete estaciones entre
0 y 1 m de profundidad; cuatro estaciones se ubicaron en la ecorregión
Morrosquillo y tres en Darién, cubriendo sustrato rocoso calcáreo
y sedimentario. En todos lo cuadrantes se evaluó la cobertura de las
algas a nivel específico. Debido a la distribución de la cobertura
algal sobre el sustrato rocoso, la ubicación del cinturón de cuadrantes
se realizó de manera preferencial aleatoria, es decir, sobre sectores
homogéneos de vegetación. El valor de cobertura obtenido corresponde
a un valor relativo de la cobertura algal sobre el sustrato rocoso, puesto que
se ubicaron los cuadrantes preferentemente sobre el sustrato dominado por algas.
Fase de laboratorio
Para el inventario de las especies se realizaron colectas de especímenes en frascos plásticos de 250 ml con una solución de formol al 4% en agua de mar y bórax. Los especímenes fueron procesados y montados en cartulina para su posterior identificación, la cual se realizó con base en los trabajos de Joly (1967), Bula-Meyer (1982a y 1982b), Ortega et al. (1993) y Littler et al. (2001).
Análisis estadístico
La estructura numérica de las comunidades de macroalgales en cada muestreo por área geográfica, se describió con base en las variaciones de cobertura, riqueza (Margalef), uniformidad (Pielou), predominio (Simpson), diversidad (Shannon-Wiener) y la similitud entre puntos de muestreo. La técnica considerada para el análisis de clasificación, se basó en el índice cuantitativo de Bray-Curtis. El dendrograma de similitud/distancia en una matriz es el producto final de un proceso aglomerativo jerárquico y politético que obedece a la ecuación lineal combinatoria de la técnica de ligamiento promedio UPGMA, el cual se realizó empleando el software PRIMER 5 versión 5.2.9.
Con el fin de establecer si la abundancia de las macroalgas difiere entre sectores de estudio y periodos climáticos, se realizó una prueba ANOSIM a una vía, a partir de la matriz de similitud con los datos de cobertura transformados con la raíz cuarta. Esta rutina se realiza con grupos determinados a priori y permite establecer si existe evidencia estadística para considerarlos entidades diferentes; se calcula a partir de una matriz de datos de similitud para obtener un estadístico llamado "r", el cual permite establecer si los sectores o periodos climáticos influyen en la estructura de la comunidad macroalgal.
RESULTADOS Y DISCUSIÓN
Se registraron 50 especies asociadas al litoral rocoso del departamento de Córdoba, distribuidas en las dos ecorregiones de estudio: 17 son clorofitas, 12 feofitas y 27 rodofitas (Tabla 4). De las 20 familias encontradas, Dictyotaceae presentó el mayor número de especies (10), seguida por Gracilariaceae (7), Rhodomelaceae (7) y Halymeniaceae (4). Los géneros con mayor número de especies fueron Gracilaria (6), Padina (5) y Caulerpa (4). El total de las especies encontradas, corresponden a comunidades epilíticas presentes en sustrato coralino muerto, rocas y acantilados rocosos.
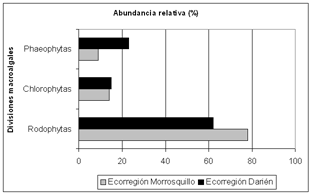
La composición de las comunidades macroalgales asociadas a la ecorregión Morrosquillo (Punta Bolívar y Punta Mestizos) estuvo integrada por tres divisiones, de las cuales las rodofitas presentaron los mayores porcentajes de cobertura (78%), seguida por las clorofitas (14%) y finalmente las feofitas, taxón con menor cobertura asociada a las rocas (9%); tendencia similar se evidenció en la ecorregión Darién, que comprende el litoral desde Punta Broqueles hasta Puerto Escondido, en donde las rodofitas registraron el mayor porcentaje de cobertura (62%), seguidas por las feofitas (23%) y finalmente las clorofitas (15%). En las dos ecorregiones, las rodofitas presentaron un mayor porcentaje de cobertura y una tendencia similar en los valores de las clorofitas (Figura 2).
El número de especies en las dos ecorregiones es relativamente bajo (50 especies), comparado con los resultados de Díaz-Pulido (1995), que reportó 151 especies para los atolones de Alburquerque y Bolívar, y los de Tsuda y Dawes (1974), quienes reportaron 100 especies para el atolón Glover en Honduras. Por otro lado, el presente estudio mostró una composición similar con los trabajos realizados por Márquez (1992) para la Isla de Providencia quien registró 50 especies, y Taylor (1975) para Isla Swan en Honduras señalando 51 especies.
Comunidades algales entre Punta Bolívar y Punta Mestizos
Este sector a nivel de intermareal se caracteriza por presentar sustrato rocoso-coralino muerto, el cual se encuentra casi completamente colonizado por céspedes o asociaciones multiespecíficas de algas (Klumpp y Mckinnon, 1992; Steneck y Dethler, 1994) de características filamentosas, coriáceas, corticadas y folioso-corticadas. Entre los céspedes multiespecíficos se destacan especies filamentosas como Bryopsis pennata, B. plumosa, Ceramium cimbricum, Centroceras clavulatum, Cladophora prolifera, C. vagabunda, Chaetomorpha linum, C. antennina, Enteromorpha flexuosa y E. intestinalis. Entre las macroalgas folioso-corticadas se destacan Dictyota spp., Padina spp. y Stypopodium zonale; las corticadas están representadas por Caulerpa spp., Gelidium spp., Hypnea spp., Acanthophora spp. y Laurencia spp. En este sector las especies coriáceas de mayor importancia en términos de cobertura son Gracilaria spp.
Esta ecorregión registró 43 especies, de las cuales Gracilaria damaecornis presentó los mayores valores de abundancia relativa (12.7%), seguida por G. cervicornis (9.03%), G. mammillaris (7.66) e Hypnea musciformis (6.15%) (Figura 3). Especies calcáreas como Halimeda opuntia y Acetabularia pussilla, así como verdes filamentosas, presentaron valores de abundancia inferior al 1%.

Comunidades algales entre Punta Broqueles y Puerto Escondido
Esta ecorregión está formada por acantilado rocoso en ciertas zonas a lo largo de su litoral, se caracteriza por la influencia de altos gradientes de corrientes y oleaje principalmente durante el periodo seco. Las comunidades macroalgales asociadas a este acantilado que se proyecta hasta una profundad de 1 m, la dominan densas praderas coriáceas de Sargassum polyceratium, mezcladas con abundantes algas rojas corticadas como Acanthophora muscoides, A. spicifera, Bryothamnion triquetrum, B. seaforthii e Hypnea musciformis y verdes filamentosas como Cladophora prolifera, Chaetomorpha linum y Enteromorpha intestinalis. Praderas de algas pardas frondosas como Padina boergesenii, Dictyota ciliolata y Stypopodium zonale se encuentran parcialmente distribuidas a manera de parches entre las densas praderas de S. polyceratium; para el sector se registraron 34 especies, de las cuales S. polyceratium (15.5%), H. musciformis (13.2%) y B. triquetrum (7.2%) presentaron los mayores porcentajes de abundancia (Figura 4), mientras que el alga folioso-membranosa Anadyomene stellata presentó la menor abundancia (menos del 1% de cobertura), presentándose como una especie rara.

Estructura algal para el área de estudio
El Atlántico Occidental presenta alrededor de 1.240 especies de algas marinas (Wynne, 1998), representando cerca de un 45% de la flora del Atlántico Occidental Tropical. Esta importante riqueza algal presente para el Caribe colombiano parece estar relacionada con la gran diversidad de hábitat marinos, presencia de surgencias, corrientes oceánicas e historias geológicas variadas. La presencia de una comunidad algal diferencial desde noreste hasta el sureste de la franja costera rocosa, indica que la flora cordobesa pertenece a ecorregiones geográficas diferentes con características propias y exclusivas (Arias et al., 2006).
Se registraron 50 especies que no eran conocidas anteriormente en el departamento
de Córdoba. Este número resulta relativamente bajo comparado con
inventarios conocidos en otras áreas del Caribe colombiano, como Santa
Marta (309), Archipiélago de San Andrés (203) y gofo de Urabá
(175) (Díaz-Pulido, 1995; Díaz-Pulido y Bula-Meyer, 1997; Díaz-Pulido,
2000; Díaz-Pulido y Díaz-Ruiz, 2003). Sin embargo, es aún
prematuro determinar que tan diversa es la flora algal costera de Córdoba
con respecto a otras localidades debido escasos estudios en el área.
En términos generales, las especies registradas en el presente trabajo
se encuentran ampliamente distribuidas en el Caribe en general (Schnetter, 1976;
1978; Bula Meyer, 1987; Bula Meyer y Schnetter, 1988; Díaz-Pulido, 2000)
y son consideradas nuevos registros para el Caribe cordobés.
Análisis de clasificación
El análisis de clasificación agrupó los puntos de muestreo de acuerdo con las ecorregiones de estudio y no mostró segregación de las estaciones hacia uno u otro sector (Figura 5); los agrupamientos de los puntos de muestreo PE, PP y PBR señalan un 74% de afinidad y corresponden a las estaciones ubicadas en la ecorregión Darién, la cual se caracteriza por ser una zona de acantilados rocosos. Los puntos de muestreo PMZ, PR, PB y PBEL propios de la ecorregión Morrosquillo, se agrupan con un 60% de afinidad y corresponden a las zonas de sustrato rocoso-coralino de fuerte influencia antrópica.
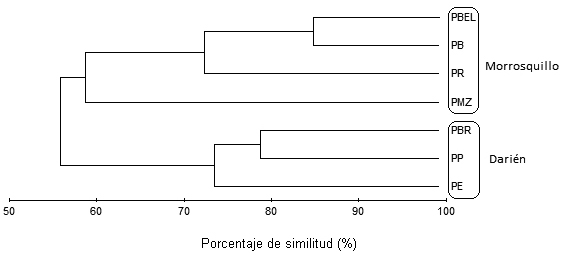
La estructura y composición de las comunidades algales en las dos ecorregiones se comportaron de manera similar durante el periodo de estudio, con un nivel de afinidad del 56%, indicando que las diferencias en las comunidades responden a las características espaciales, y no dependen significativamente de los períodos climáticos del año.
La prueba ANOSIM a una vía, que relaciona los períodos climáticos y las estaciones de muestreo con los porcentajes de cobertura algal en el sector entre Punta Bolívar y Punta Mestizos, señala que no existen diferencias significativas entre la abundancia y composición de especies con relación a los períodos climáticos del año; por el contrario, se presentaron diferencias significativas entre la abundancia y composición de especies con relación a los puntos de muestreo en dicho sector (P<0.05; R=0.78). Un comportamiento similar se presentó en el sector entre Punta Broqueles y Puerto Escondido, en donde el patrón de distribución y abundancia de las especies se muestra más influenciado por los puntos de muestreo que por los periodos climáticos del año (Tabla 5).

El Mar Caribe cordobés no presenta núcleos de surgencias que estimulen un cambio sucesional en los ensamblajes algales, como los registrados por Bula Meyer (1985) en Santa Marta. Esta condición permite el establecimiento de una comunidad algal homogénea durante todo el año, que ajusta su cobertura a las diferentes condiciones ambientales. Sin embargo algunas especies como Padina boergesenii y P. haitiensis aparecieron durante las temporadas lluviosas, estableciéndose como especies macroalgales exclusivas para este periodo.
Índices ecológicos
En la Figura 6 (A y B), se observan las gráficas de diversidad y riqueza durante los 12 meses de estudio. El litoral comprendido entre Punta Bolívar y Punta Mestizos registró el valor más bajo de diversidad en septiembre con 2.56 y el de riqueza en agosto con 4.8, mientras que los valores más altos de diversidad y riqueza (3.0 y 6.3 respectivamente) se presentaron en diciembre. El litoral comprendido entre Punta Broqueles y Puerto Escondido mostró un comportamiento similar con relación a la ecorregión Morrosquillo, presentándose en septiembre el menor valor de diversidad (2.1) y en agosto el menor valor de riqueza (3.9), mientras que en diciembre se presentaron los valores más altos de diversidad y riqueza (2.94 y 5.8 respectivamente); el índice de predominio de Simpson (0.08 para el sector 1 y 0.11 para el sector 2), mostró que los dos sectores presentan una estructura macroalgal homogénea en cuanto no existe una especie que predomine en términos de cobertura, lo cual se ajusta a lo arrojado por el índice de Pielou, cuyos valores oscilaron entre 0.77 para el sector 1 y 0.8 para el sector 2, ratificando la idea de estructuras estables respecto al número de especies.
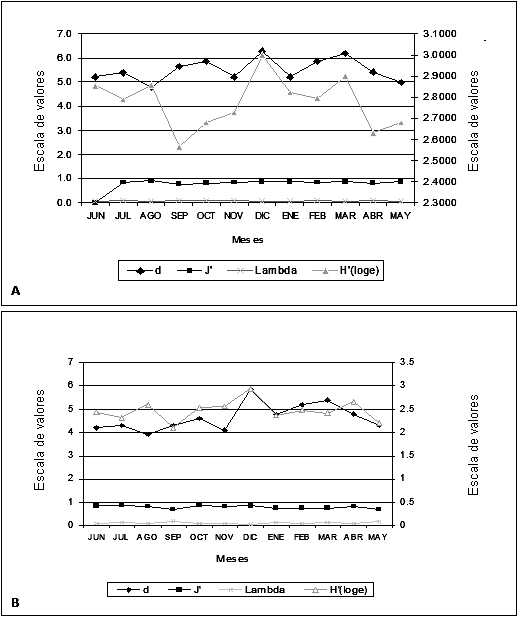
Los valores de diversidad fueron bajos comparados con otros estudio realizados en el Caribe colombiano (Díaz-Pulido, 2000; Díaz-Pulido y Garzón Ferreira, 2002). La causa de este bajo nivel de diversidad puede deberse a la escasa disponibilidad de sustrato rocoso que restringen el establecimiento de una gran variedad de especies y proveen las condiciones para un punto de baja diversidad de macroalgas en Córdoba. Las regiones de mayor diversidad son el Parque Nacional Natural Tayrona, seguido por el noreste chocoano en la parte más sur del Caribe colombiano y el Archipiélago de San Andrés y Providencia, probablemente este alto patrón de diversidad responde al intenso esfuerzo de muestreo en estas áreas, así como a la variedad de ecosistemas y microhábitats (García y Díaz-Pulido, 2006). En este sentido, y como consecuencia de los escasos estudios en algunas áreas de la costa central del Caribe colombiano como Atlántico, Bolívar, Sucre y Córdoba; se recomienda más investigaciones sobre la diversidad, abundancia y distribución de las comunidades macroalgales en estos sectores.
AGRADECIMIENTOS
Los autores agradecen Al profesor Guillermo Díaz Pulido de la Universidad del Magdalena, por su colaboración en la identificación del material biológico y capacitación de los tesistas a través de pasantías realizadas en dicha institución. Al profesor Adolfo Sanjuán Muñoz de la Universidad Jorge Tadeo Lozano, por la asesoría estadística y ecológica en el tratamiento de las muestras, además de las sugerencias al documento. Al Centro de Investigación Piscícola de la Universidad de Córdoba (CINPIC), por el apoyo con la infraestructura y equipos que permitieron la toma de fotografías a los especímenes. Y finalmente a la Oficina de Administración de Investigación y Extensión de la Universidad de Córdoba, por haber aportado los Recursos Económicos necesarios para la realización del presente trabajo.
LITERATURA CITADA
1. ADEY, W.H & R.S. STENECK. 1985. Highly productive eastern Caribbean
reefs: synergistic effects of biological, chemical, physical, and geological
factors. In: REAKA, M. L. (Ed.). The ecology of coral reefs. Symposium Series
of Undersea Research. NOAA Undersea Research Program, Rockville, Maryland. v.
3, p. 163-187.
2. ANGEL & H. QUIRÓS. 1971. Contribución al conocimiento ecológico
y sistemático de las algas marinas de Cartagena y sus alrededores. 98
pp.[Tesis de grado]. Universidad Jorge Tadeo Lozano, Cartagena.
3. ARIAS, J., P. DUEÑAS & J. QUIRÓS. 2006. Distribución
espacio temporal de las comunidades macroalgales asociadas al litoral rocoso
del departamento de Córdoba, Caribe colombiano. Facultad de Ciencias
Básicas e Ingenierías. Universidad de Córdoba. Montería.
109 pp.
4. BAK, R.P.M. 1976. The growth of coral colonies and the importance of crustose
coralline algae and burrowing sponges in relation with carbonate accumulation.
Netherlands Journal of Sea Research 10(3): 285-337.
5. BULA-MEYER, G. 1982a. Una clave para las especies del género Halimeda
(Udotaceae, Chlorophyta) del Atlántico occidental. An. Ins. Inv. Mar.
Punta Betín 12:41-44.
6. BULA-MEYER, G. 1982b. Adiciones a las Clorophytas marinas del Caribe colombiano.
An. Ins. Inv. Mar. Punta Betín 12:117-136.
7. BULA-MEYER, G. 1985. Un núcleo nuevo de surgencia en el Caribe colombiano
detectado en correlación con las macroalgas. Boletín Ecotrópica
12: 3-25.
8. BULA-MEYER, G. 1986. Las macroalgas de los arrecifes coralinos de las Islas
del Rosario, costa Caribe de Colombia. Boletín Ecotrópica 14:3-20.
9. BULA-MEYER, G. 1987. Taxonomic and ecologic studies of a subtidal sand plain
macroalgal community in the Colombian Caribbean. 189 pp[Tesis Ph. D.] Univ.
Of. Delawer, .
10. BULA-MEYER, G. & R. SCHNETTER. 1988. Las macroalgas recolectadas durante
la expedición Urabá II, costa Caribe del noreste chocoano, Colombia.
Bot. Ecotrópica 18:19-32.
11. CONNOR, J.L. & W.H. ADEY. 1977. The benthic composition, standing crop,
and Productivity of a Caribbean algal ridge. Atoll Res. Bull 211:1-15
12. CVS. 2004. Diagnostico Ambiental de las cuencas de la Cuenca del Río
Canalete. Documento en elaboración Convenio CVS - FONADE No. 193040.
13. DAWES, C. 1986. Botánica Marina. Limusa, S.A. de C.V., México,
673 pp.
14. DÍAZ-PULIDO. G. 1995. Comunidades macroalgales de los atolones Alburquerque
y Courtown (Bolívar), Caribe suroccidental, Colombia, y su relación
con la estructura arrecifal. [Tesis de grado]. Universidad Jorge Tadeo Lozano,
Bogotá, D.C.
15. DÍAZ-PULIDO, G. 2000. Vegetación marina de un sector de la
plataforma continental de las Guajira (Caribe colombiano). Bol. Invest. Mar.
Cost. 29: 27-34.
16. DÍAZ-PULIDO, G. & G. BULA-MEYER. 1997. Litorales marinos y costeros.
Programa de Biodiversidad y Ecosistemas Marinos. 115-128. En: INVEMAR: Informe
Nacional sobre el estado de la biodiversidad en Colombia; año 1997. INVEMAR,
Santa Marta. 143 p.
17. DÍAZ-PULIDO, G. & J. GARZÓN-FERREIRA. 2002. J. Seasonality
in algal assemblages on upwelling-influenced coral reefs in the Colombian Caribbean.
Botánica Marina 45(3):284-292.
18. DIAZ-PULIDO, G. & M. DIAZ-RUIZ. 2003. Diversity of benthic marine algae
of the Colombian Atlantic. Biota Colombiana 4(2):203-246.
19. GARCÍA, C.B. & G. DIAZ-PULIDO. 2006. Dynamics of a macroalgal
rocky intertidal community in the Colombian Caribbean. Boletín de Investigaciones
Marinas y Costeras 35:7-18.
20. GARZÓN-FERREIRA, J. & J. CANO. 1991. Tipos, distribución,
extensión y estado de conservación de los ecosistemas marinos
costeros del Parque Nacional Natural Tayrona. Manuscrito. Versión presentada
al Séptimo Concurso Nacional de Ecología "Enrique Pérez
Arbeláez". Fondo FEN-COLOMBIA, Santa Marta, 82 pp.
21. FRITHSEN, J. & A. HOLLAND. 1992. Benthics communities as indicator of
ecosystem conditions. En: Ecological indicator. Chapman and hall. Vol. 1, pp.
459-460.
22. HOLDRIDGE, L. 1978. Ecología basada en las zonas de vida. Instituto
Interamericano de Ciencias Agrícolas, San José.
23. INVEMAR-MINISTERIO DEL MEDIO AMBIENTE. 2000. Programa nacional de investigación
en biodiversidad marina y costera PNIBM, plan de acción 2001-2010. Ministerio
del Medio Ambiente. An. Inst. Invest. Mar. Punta Betín. Santa Marta,
83 pp.
24. JOLY, A. 1967. Géneros de algas marinas da Costa Atlántica
Latino-Americana. Editora da Universidade de São Paulo. 462 p.
25. KLUMPP, D.W. & A.D. MCKINNON. 1992. Community structure, biomass and
productivity of epilithic algal communities on the Great Barrier Reef: dynamics
at different spatial scales. Marine Ecology Progress Series 86:77-89.
26. LEWIS, R. 1985. Water movements and their role in the rocky shore ecology.
Sarsia 34: 13-36.
27. LITTLER, M.M., D.S. LITTLER & P.R. TAYLOR. 1983a. Evolutionary strategies
in a tropical barrier reef system: functional-form groups of marine macroalgae.
J. Phycol. 19:229-237.
28. LITTLE, D.S., M.M. LITTLE, K.E. BUCHER & J.N. NORRIS. 2001. Marine Plants
of the Caribbean. Smithsonian Institution. Press. Washington, 263 pp.
29. LITTLER, M.M., P.R. TAYLOR, D.S. LITTLER, R.H. SIMS & J.N. NORRIS. 1987.
Dominant macrophyte standing stocks, productivity and community structure on
a Belizean barrier reef. Atoll Res. Bull. 302: 1-18.
30. MÁRQUEZ, G.E. 1992. Estudios ecológicos en el complejo arrecifal
de Providencia isla, Caribe occidental colombiano: Ecología arrecifal
y vegetación marina. Memorias VIII Sem. Nal. de Cien. y Tec. del mar,
Santa Marta, 397-422p.
31. MORRISON, D. 1988. Comparing fish and urching in shallow and deeper coral
algal communities. Ecology 69(5): 1367-1382.
32. PATIÑO, F. & F. FLÓREZ. 1993. Ecología Marina del
Golfo de Morrosquillo. Universidad Nacional de Colombia. Fondo (FEN), Colombia.
Bogotá, D.C.
33. QUIRÓS, J. 2009. Dinámica espacial de la composición
y estructura de las poblaciones de crustáceos decápodos asociados
a céspedes algales del litoral rocoso del departamento de córdoba,
Caribe colombiano.
138 pp.[Tesis de Maestría]. Universidad de Córdoba, Facultad de Ciencias
Básicas e Ingenierías, Montería,
34. RANGEL-BUITRAGO, G & B.O. POSADA-POSADA. 2005. Geomorfología
y procesos erosivos en la costa Norte del departamento de Córdoba. Bol.
Invest. Mar. Cost. 34: 101-119.
35. SCHNETTER, R. 1976. Algas Marinas de la Costa Atlántica colombiana.
Biblioteca Phycologica I Phaeophyceae J. Cramer, Vaduz.
36. SCHNETTER, R. 1978. Algas Marinas de La costa Caribe colombiana, Biblioteca
Phycologica II Chlorophyceae J. Cramer, Vaduz.
37. TAYLOR, W.R. 1975. Marine algae of Great Swan Island. Atoll Res. Bull. 185:6-10p.
38. TSUDA R.T. & C.J. DAWES. 1974. Preliminary checklist of the marine benthic
plants from Glover's reef, British Honduras. Atoll Res. Bull. 173:1-13
39. VAN DEN HOECK, C., A.M. BREEMAN & J.B.W. WANDER. 1975. Algal zonation
in the fringing coral reef of Curacao, Netherlands Antilles, in relation to
zonation of coral reef and Gorgonians. Aquat. Bot. 1:269-308.
40. VAN DEN HOECK, C., A.M. BREMAN, R.P.H. BAK & VAN G. BUURT. 1978. The
distribution of algae, corals and gorgonians in relation to depth, light attenuation,
water movement and grazing pressure in the fringing coral reef of Curacao, Netherlands
Antilles. Aquat. Bot. 5:1-46.
41. VERNETTE, G. 1985. La plateforme Continentale Caribe de Colombie (du debouche
du Magdalena au Golfe de Morrosquillo). Tesis Doctoral a L' Universite
de Bordeaux, Bordeaux.
42. WANDERS, J.B.W. 1976. The role of benthic algae in the shallow reef of Curacao
(Netherlands Antilles). Aquat. Bot. 2:235-390.
43. WEDLER, E. 1998. Introducción en la Acuicultura, con Énfasis
en los Neotrópicos. Primera impresión, Santa Marta, 367 pp.
44. WYNNE, M.J. 1998. A checklist of benthic marine algae of the tropical and
subtropical western Atlantic: first revision. Nova Hegwigia 116: 1-155.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2010 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















