



Publicado
Validación y aplicación de un modelo de restauración de manglar basado en individuos para tres especies en la Ciénaga Grande de Santa Marta
DOI:
https://doi.org/10.15446/caldasia.v38n2.55360Palabras clave:
Manglares, modelos basados en individuos, monitoreo, rehabilitación (es)En la década de los años 90 los manglares de la Ciénaga Grande de Santa Marta (CGSM) murieron masivamente por los altos niveles de salinidad en el suelo. Este hecho estimuló la reapertura de cinco canales naturales para reducir la salinidad y mejorar las condiciones del bosque (Proyecto PROCIENAGA). Con base en esta experiencia, en 1999 un modelo basado en individuos (FORMAN) fue desarrollado para simular trayectorias de restauración del bosque; sin embargo, este modelo fue restringido a un único sitio de la CGSM y a la fecha no había sido probado. Para mejorar su sensibilidad y predictibilidad para simular tendencias del Área Basal (AB) de los árboles de Rhizophora mangle, Avicennia germinans y Laguncularia racemosa en este estudio se validó, calibró y ajustó el modelo FORMAN, usando datos de 16 años de monitoreo. Para ello, se modificó la interfaz del modelo y se ajustaron algunas rutinas de cálculo y parámetros. Se ejecutaron dos escenarios de simulación: (1) Asumiendo estabilidad en el intervalo de variación de la salinidad y, (2) aumentado la salinidad de acuerdo con el incremento promedio de los últimos tres años. Los ajustes en las tasas de reclutamiento y salinidad calculada permitieron calibrar el modelo y reproducir el patrón y la magnitud observada en tres estaciones. En dos estaciones solo se reprodujo el patrón de los datos. Las tendencias generales del AB de cada especie y su dinámica, así como las magnitudes actuales para las estaciones que sufrieron mayor impacto inicialmente, fueron reproducidas adecuadamente. Bajo un escenario en el que la salinidad se mantiene en un intervalo estable, el sistema alcanzaría la estabilidad en el largo plazo (>100 años). Un incremento promedio constante en la salinidad intersticial, resultaría en una disminución severa del AB en el corto plazo (menos de 20 años), demostrando la vulnerabilidad de la CGSM a los incrementos en salinidad y la necesidad de implementar medidas de manejo sostenibles en el tiempo. El estudio contribuye al entendimiento de la dinámica de lagunas costeras tropicales en el largo plazo y resalta la importancia de la modelación como herramienta básica para los proyectos de rehabilitación y manejo.
doi: https://dx.doi.org/10.15446/caldasia.v38n2.55360
Ecología
Validación y aplicación de un modelo de restauración de manglar basado en individuos para tres especies en la Ciénaga Grande de Santa Marta
Validation and application of an individual based restoration model for three mangrove species in Ciénaga Grande de Santa Marta
Jenny Alexandra Rodríguez-Rodríguez
Instituto de Investigaciones Marinas y Costeras de Colombia "José Benito Vives de Andreis" INVEMAR. Calle 25 No. 2-55, Playa Salguero, Santa Marta D.T.C.H., Colombia.
José Ernesto Mancera-Pineda
Universidad Nacional de Colombia, Sede Bogotá, jemancerap@unal.edu.co
Juan Manuel Rodríguez-P.
Instituto de Investigaciones Marinas y Costeras de Colombia "José Benito Vives de Andreis" INVEMAR. Calle 25 No. 2-55, Playa Salguero, Santa Marta D.T.C.H., Colombia.
RESUMEN
En la década de los años 90 los manglares de la Ciénaga Grande de Santa Marta (CGSM) murieron masivamente por los altos niveles de salinidad en el suelo. Este hecho estimuló la reapertura de cinco canales naturales para reducir la salinidad y mejorar las condiciones del bosque (Proyecto PROCIENAGA). Con base en esta experiencia, en 1999 un modelo basado en individuos (FORMAN) fue desarrollado para simular trayectorias de restauración del bosque; sin embargo, este modelo fue restringido a un único sitio de la CGSM y a la fecha no había sido probado. Para mejorar su sensibilidad y predictibilidad para simular tendencias del Área Basal (AB) de los árboles de Rhizophora mangle, Avicennia germinans y Laguncularia racemosa en este estudio se validó, calibró y ajustó el modelo FORMAN, usando datos de 16 años de monitoreo. Para ello, se modificó la interfaz del modelo y se ajustaron algunas rutinas de cálculo y parámetros. Se ejecutaron dos escenarios de simulación: (1) Asumiendo estabilidad en el intervalo de variación de la salinidad y, (2) aumentado la salinidad de acuerdo con el incremento promedio de los últimos tres años. Los ajustes en las tasas de reclutamiento y salinidad calculada permitieron calibrar el modelo y reproducir el patrón y la magnitud observada en tres estaciones. En dos estaciones solo se reprodujo el patrón de los datos. Las tendencias generales del AB de cada especie y su dinámica, así como las magnitudes actuales para las estaciones que sufrieron mayor impacto inicialmente, fueron reproducidas adecuadamente. Bajo un escenario en el que la salinidad se mantiene en un intervalo estable, el sistema alcanzaría la estabilidad en el largo plazo (>100 años). Un incremento promedio constante en la salinidad intersticial, resultaría en una disminución severa del AB en el corto plazo (menos de 20 años), demostrando la vulnerabilidad de la CGSM a los incrementos en salinidad y la necesidad de implementar medidas de manejo sostenibles en el tiempo. El estudio contribuye al entendimiento de la dinámica de lagunas costeras tropicales en el largo plazo y resalta la importancia de la modelación como herramienta básica para los proyectos de rehabilitación y manejo.
Palabras clave. Manglares, modelos basados en individuos, monitoreo, rehabilitación.
ABSTRACT
During the 1990s, the mangroves of the Ciénaga Grande de Santa Marta (CGSM) experienced a massive die-off due to high levels of soil salinity. In response, five natural channels were reopened to reduce salinity and improve the forests' conditions (PROCIENAGA Project). Based on this experience, in 1999, an Individual Based Model (FORMAN) was developed to simulate forest restoration trajectories; however, this model was restricted to one site in the CGSM and until now it has not been tested. In order to improve its accuracy and predictability in simulations of basal area (BA) trends of the tree species Rhizophora mangle, Avicennia germinans y Laguncularia racemosa we validated, calibrated, and adjusted the FORMAN model using 16 years of monitoring data. We modified the model interface and adjusted calculation routines and parameters. We performed two simulation scenarios: (1) Assuming a stable range of variation in salinity and (2) Increasing salinity based on the average increase over the last three years. The adjustments to the salinity calculation and recruitment rates allowed us to calibrate the model and reproduce both the pattern and magnitude of the observed data in three stations. In two stations we could reproduce only the pattern of the data. The model accurately reproduced general trends in changes in the BA of each species, as well as the current BA for stations that initially suffered greater impact. Under a scenario of stable soil salinity, the system will reach stability in the long term (> 100 years), while a constant increase in interstitial salinity, would result in a major decrease in BA in the short term (<20 years). These results show the vulnerability of CGSM to increases in salinity and the need to implement sustainable management practices. This study contributes to the understanding of long term dynamics in tropical coastal lagoons and highlights the importance of modeling as a basic tool for rehabilitation and management projects.
Keywords. Mangroves, individual based models, monitoring, rehabilitation.
INTRODUCCIÓN
La dinámica de los humedales costeros está influenciada por distintos factores biológicos, climáticos y geológicos haciendo que su estudio y gestión sean complejos. Para facilitar estos procesos, la modelación ecológica ha surgido como una herramienta que permite simplificar la realidad al analizar algunos aspectos del sistema a emular, funcionando como substitutos del mismo (Blanco 2013). Para apoyar la selección de medidas de éxito en los esfuerzos de restauración ecosistémica y evaluar su progreso, los modelos ecológicos dinámicos han surgido como un instrumento integral capaz de simular metas más realistas, considerando los largos tiempos que requieren este tipo de proyectos (Twilley & Rivera-Monroy 2005). Los modelos, así mismo, pueden ser usados para establecer trayectorias de respuesta de manera temprana a una variedad de condiciones, sintetizar resultados de estudios de campo, y dirigir los esfuerzos de investigación para probar teorías e hipótesis específicas acerca de los procesos ecológicos (Twilley et al. 1998, Berger & Hildenbrandt 2000, Berger et al. 2006).
El proceso de retroalimentación de los programas de monitoreo a los modelos propuestos es estratégico para su validación pues permite probar la plausibilidad de las hipótesis expuestas en las simulaciones, por ejemplo, en un amplio ámbito geográfico sobre aspectos particulares de los manglares y su ecología (Berger et al. 2008). En el campo de la restauración ecológica, solo recientemente algunos trabajos han empleado marcos conceptuales o técnicas específicas donde la modelación ecológica se usa como herramienta para cuantificar efectos y respuestas de los ecosistemas en los procesos de restauración o apoyar la toma de decisiones relacionadas con estos proyectos (manejo adaptativo), entre otros (Twilley et al. 1998, Wang et al. 2010, Fitz et al. 2011, Obeysekera et al. 2011, Rodhouse et al. 2011, Visser et al. 2013, Martínez et al. 2014).
Los modelos basados en individuos, o IBMs, por sus siglas en inglés provenientes de Individual Based Models, han demostrado ser adecuados para entender la "biocomplejidad" de los manglares; es decir, las interrelaciones entre todos los componentes del ecosistema, incluyendo las sociedades humanas, a múltiples escalas espacio temporales (Feller et al. 2010). Así mismo, han servido para explicar patrones complejos a nivel de población y para predecir los impactos inducidos por los disturbios naturales y por el ser humano en la dinámica de los bosques (Berger et al. 2008). En general, los IBM se basan en la representación de la dinámica de la comunidad a través de la descripción explícita de la respuesta de cada individuo a los mecanismos que estocásticamente actúan sobre él, reflejando las interacciones con sus vecinos y su entorno (Campillo & Champagnat 2012).
Los manglares son humedales característicos de las zonas costeras tropicales y subtropicales del mundo. Las variaciones en estructura y función de estas comunidades vegetales se relacionan con la disponibilidad de recursos reguladores y gradientes hidrológicos que interactúan a diferentes escalas (Feller et al. 2010). De tal forma, por ejemplo, en las especies de manglar del neotrópico, los distintos grados de tolerancia a la salinidad (Cintrón et al. 1978, Odum et al. 1982), la distribución de nutrientes en el suelo, principalmente de Nitrógeno y Fosforo (Feller et al. 2003) y la tolerancia a la luz (Ball 1980), han mostrado modelar la composición y estructura de los bosques (Chen & Twilley 1998, Cardona-Olarte et al. 2013).
En Colombia hacia la década de los años 90 se desarrolló el proyecto PROCIENAGA, considerado como la iniciativa de restauración de manglares más ambiciosa en Latinoamérica (Botero & Salzwedel 1999; Rivera-Monroy et al. 2001, Cotes 2004). El proyecto fue formulado ante el deterioro ambiental del complejo estuarino de la Ciénaga Grande de Santa Marta (CGSM), el cual se vio reflejado entre otros, en la pérdida de 28.570ha de manglar entre 1956 y 1995 (INVEMAR 2014), como consecuencia entre otros, de los altos niveles de salinidad en suelo, que en 1993 alcanzaron en promedio 120 unidades. A través de la reapertura de cinco canales naturales, el proyecto pretendía modificar el régimen hidrológico de las áreas afectadas a través de la entrada de agua del río Magdalena reduciendo la salinidad de los suelos. Tras la culminación de las obras y como estrategia para monitorear el proceso de recuperación de la CGSM, se implementó el "programa de monitoreo de las condiciones ambientales y las condiciones estructurales y funcionales de las comunidades vegetales y de los recursos pesqueros durante la rehabilitación de la Ciénaga Grande de Santa Marta". El programa, que permanece activo hasta la fecha, representa un conjunto de datos único e invaluable con más de 18 años de mediciones, luego de un proceso de rehabilitación en los ambientes marinos. Estas mediciones incluyen elementos bióticos y variables fisicoquímicas en aguas y sedimentos (INVEMAR 2014).
Con los resultados parciales de este moni-toreo y estudios preliminares de Giraldo (1995) y Cardona & Botero (1998), Twilley et al. (1998) adaptaron el modelo ecológico FORMAN, que es un IBM, para simular trayectorias de restauración en una estación de monitoreo de la CGSM. Sin embargo, a la fecha el modelo no se había verificado, calibrado, ni complementado. Por lo tanto, los objetivos específicos de este estudio fueron: (1) validar y calibrar el modelo ecológico FORMAN desarrollado para la CGSM (Twilley et al. 1998) para las tres especies de mangle Avicennia germinans (L.) L., Laguncularia racemosa (L.) C.F. Gaertn, Rhizophora mangle L. (2) simular escenarios futuros de potenciales respuestas específicas de cada una de las tres especies de manglar, y finalmente, (3) potenciar una herramienta para el manejo oportuno y sostenible del complejo estuarino.
MATERIALES Y MÉTODOS
Descripción del Modelo
El IBM FORMAN, desarrollado por Chen & Twilley (1998) para manglares neotropicales, tiene como objetivo predecir el crecimiento anual de cada árbol de manglar en un área de estudio determinada. Para ello, establece las condiciones ideales de crecimiento de los árboles de manglar según características propias de su historia de vida y evalúa el efecto del estrés y la disponibilidad de recursos en el crecimiento de cada individuo a través de funciones modificadoras.
A continuación, se muestra la función integrada por FORMAN para predecir el crecimiento anual y las variables involucradas en las funciones modificadoras de crecimiento. Los parámetros requeridos para su cálculo se pueden observar con detalle en Chen & Twilley (1998) y Twilley et al. (1998).
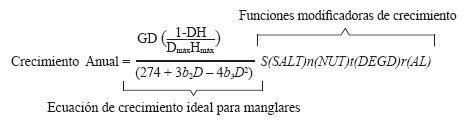
Donde:
D: Diámetro del árbol a la altura del pecho (DAP); H: altura del árbol (cm); Dmáx: Diámetro máximo que puede alcanzar un árbol; Hmax: Altura máxima que puede alcanzar un árbol; G, b2, b3: Constantes de crecimiento especie específicos; S (SALT): Efecto de la salinidad; n (NUT): Efecto de la disponibilidad de nutrientes en suelo; t (DEGD): Efecto de la temperatura medida en Grados Día1; r (AL): Efecto de la intensidad lumínica.
El modelo parte con un número determinado de árboles y anualmente contempla la adición de nuevos individuos. Como recursos de salida, el modelo incluye eventos de mortalidad estocástica basados en las observaciones de Botkin et al. (1972), según las cuales: (1) la probabilidad de morir de un árbol con un crecimiento activo es independiente de su edad, sin embargo, solo tiene una probabilidad del 2% de sobrevivir hasta la edad máxima y (2) la probabilidad de muerte se incrementa al 36,5% si el crecimiento en diámetro de un árbol es menor a 0,01cm por dos años consecutivos.
Implementación y validación
El código original del modelo FORMAN fue desarrollado en lenguaje de programación "C". Para la implementación del modelo en la CGSM, se realizó un desarrollo propio a partir del código original, con el fin de implementar una interface gráfica de fácil manejo que pudiera ser operada en cualquier sistema operativo con soporte JAVA.
El modelo se ejecutó para cinco estaciones de monitoreo con diferente grado de perturbación en los bosques de manglar de la CGSM (Tabla 1). Para inicializar el modelo, se establecieron las condiciones del bosque inicial utilizando los datos derivados del proyecto de monitoreo ejecutado por el instituto de investigaciones marinas y costeras de Colombia INVEMAR (INVEMAR 2014). Para ello, se tomaron los datos de composición de especies y diámetro a la altura del pecho (DAP) del primer año de monitoreo en cada estación; se conservaron los supuestos, parámetros, y constantes descritas por Twilley et al. (1998). Para validar el modelo, las predicciones arrojadas se compararon con las observaciones desde 1995 hasta el 2014.

Calibración
Con el fin de mejorar el ajuste del modelo FORMAN original a las tendencias observadas en el bosque de manglar de la CGSM, se implementaron los siguientes ajustes:
- Modificación en el uso de las variables: Para el ingreso de las variables salinidad, nutrientes y temperatura, se implementó la posibilidad de que el modelador ingrese valores y funciones predefinidas como lineal, exponencial o aleatoria. Para la aplicación del modelo en este caso y asumiendo el supuesto inicial de Twilley et al. (1998), se simuló la disminución en los valores de salinidad hasta el año donde esta alcanzó el valor de referencia definido por el autor que es de 40 unidades; luego de este umbral, se usó una función aleatoria con valores que oscilaron alrededor de la mediana de los valores de salinidad históricos, teniendo en cuenta sus desviaciones.
- Modificación de la rutina de reclutamiento: Las nuevas plántulas que ingresan a la simulación poseen un diámetro de 0,1 cm en lugar de los 1,27 cm propuestos en el modelo inicial de Twilley et al. (1998). Este valor se obtuvo a partir del tamaño mínimo promedio de las plántulas medidas en campo. Además, se activó un valor aleatorio de plántulas en el año inicial, relacionado con el factor de reclutamiento para cada especie de mangle. Para los bosques de la CGSM y otros bosques del caribe colombiano, las plántulas se convierten en reclutas una vez alcanzan un DAP de 2,5 cm, es decir que entran a considerarse para el cálculo de área basal y densidad (INVEMAR 2014, Hoyos et al. 2013).
- Ajuste de las constantes del número de plántula reclutadas: El número de plántulas disponibles para inicializar el modelo en cada estación monitoreada en la CGSM fue ajustado con base en los valores promedio de abundancia de plántulas observados en campo desde el año 2002 hasta el año 2014. Las constantes calculadas y ajustadas mediante ensayos de sensibilidad se muestran en la Tabla 2.

- Modificación del cálculo de luz disponible: La luz disponible es utilizada para el cálculo de uno de los factores de la ecuación de crecimiento ideal y para el reclutamiento, utilizando la ecuación de expresada en Botkin et al. (1972):
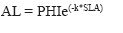
Donde:
AL: Luz disponible;
SLA: Sumatoria de las áreas de sombra generada por todos los árboles mayores al que se le está calculado la luz disponible;
PHI: Insolación anual = 1 (según Botkin et al. 1972).
k = constante de sombreado bajo un dosel denso = 1/6000 para 100 m2 (según Botkin et al. 1972); 1/140000 para un área de 500 m2 (según Twilley et al. 1998).
Teniendo en cuenta las variaciones en área de muestreo observadas en el monitoreo de la vegetación en la CGSM, se construyó un k dependiente del área considerando los valores propuestos por Botkin et al. (1972) y Twilley et al. (1998) y asumiendo un comportamiento lineal, obteniéndose la siguiente ecuación:

Donde A es el área que puede ser ajustada durante la simulación.
Escenarios futuros
Una vez ajustado y calibrado el modelo se simularon escenarios futuros en el comportamiento del AB de las estaciones monitoreadas según los siguientes escenarios:
Escenario 1: Mantenimiento de las condiciones abióticas en intervalos similares a los observados en el presente. Para ello se hallaron los intervalos de variación de la salinidad históricos y se generaron funciones aleatorias oscilantes alrededor de estos valores. Las simulaciones se corrieron con 10 iteraciones y el resultado se proyectó hasta el año 2500, permitiendo ver los cambios y el comportamiento de las estaciones en el corto, mediano y largo plazo.
Escenario 2: Incremento en la salinidad de cada estación de acuerdo con la tendencia registrada en el monitoreo ambiental de la CGSM en los últimos años. Para esto se calculó el incremento promedio anual desde el 2012 hasta el 2014 en cada estación así: Aguas Negras: 8,8; Luna: 17,5; Kilómetro 22: 33,66; Rinconada; 7,1 y Caño Grande: 6,75. Para encontrar la salinidad en el 2015, este incremento anual se sumó al valor promedio de salinidad reportado para cada estación en el año 2014. El modelo se proyectó sumando el incremento promedio anual cada año. Las simulaciones se corrieron con 10 iteraciones y se proyectaron hasta el año 2035, permitiendo ver los cambios y el comportamiento de las estaciones en el corto y mediano plazo.
RESULTADOS
Validación
Los resultados al implementar el modelo formulado por Twilley et al. (1998) mostraron discrepancias en términos de magnitud y tendencia del Área Basal (AB) al ser comparadas con las observaciones en campo de las cinco estaciones monitoreadas actualmente en la CGSM (Fig. 1).

El AB observada en campo para la estación LUN alcanzó para el 2014 un valor total de 25,08 m2/ha, siendo L. racemosa la especie con mayor AB (18,03 m2/ha). Sin embargo, el patrón modelado para la misma estación siguiendo la propuesta de Twilley et al. (1998), para el 2014 preveía una AB total de 7,45m2/ha con una mayor contribución de A. germinans (AB=6,01 m2/ha). Las proyecciones para los años 2014 en las estaciones KM22, ANE y CGE preveían un AB total de 18,34 m2/ha, 4,50 m2/ha y 11,63 m2/ha, respectivamente; mientras que para este año las observaciones en campo indicaron un AB total de 28,07 m2/ ha, 27,34 m2/ha y 19,20 m2/ha demostrando que los valores simulados están en algunos casos por debajo de los observados en un orden de magnitud. Los cambios de las especies a lo largo del tiempo en términos de AB tampoco fueron mostrados por las simulaciones, principalmente en la estación KM22 donde los modelos predecían mayor AB para L. racemosa desde el 2001 y para LUN un importante AB para A. germinans hasta el final de la simulación. Las observaciones hechas en campo demostraron que en KM22 A. germinans incrementó su AB desde 2003 hasta el presente, mientras que L. racemosa disminuyó rápidamente a partir desde ese mismo año.
Calibración
Luego de implementar los cambios descritos en la metodología se observó un buen ajuste general de los modelos generados frente a las observaciones hechas en campo. Los resultados se muestran en la tercera columna de la Figura 1.
Las tendencias generales del AB de cada especie y su dinámica, así como las magnitudes actuales para las estaciones que sufrieron mayor impacto inicialmente fueron reproducidas adecuadamente por el modelo ajustado. Así por ejemplo, para el año 2014 el AB total observado para el KM22 fue de 28,07 m2/ha y el valor modelado de 24,8 m2/ha; para ANE el AB total observado fue de 27,3 m2/ha y el valor modelado de 29,9 m2/ha; finalmente, para LUN el AB observado fue de 22,1 m2/ha, mientras que el valor modelado fue de 25,08 m2/ha (Fig. 1). Las tendencias de las especies a lo largo del tiempo y su dinámica, también fueron bien representadas para las estaciones con menor impacto inicial como CGE y RIN, aunque las magnitudes fueron sobreestimadas (AB Total CGE: Valor observado: 19,21 m2/ha, Valor Modelado: 49,51 m2/ha; AB Total RIN: Valor observado: 35,01 m2/ha, Valor Modelado: 53,23m2/ha).
Simulación de escenarios futuros
Según la simulación ejecutada, conservando la tendencia de la salinidad mostrada por el Monitoreo Ambiental CGSM hasta el año 2014, A. germinans mantendría la mayor AB en KM22 hasta el año 2243 aproximadamente con una máxima AB registrada de 95,52 m2/ ha, cuando ésta empezaría a descender hasta cerca de los 70 m2/ha en el año 2500. El AB de L. racemosa se mantendría en ascenso tal como se observa en el presente y continuaría hasta el año 2108 alcanzando un AB de 22,57 m2/ha, tiempo después del cual disminuiría significativamente hasta 0,26 m2/ha en el año 2500. El AB de R. mangle mantendría niveles bajos, tal como ahora, alcanzando un máximo AB de 1,03 m2/ha en el año 2120, esta área basal empezaría a disminuir hasta llegar a la desaparición de la especie en el año 2347 (Fig. 2a).

En la estación ANE a los 15 años la dominancia de L. racemosa se remplazaría por la de A. germinans y posteriormente nuevamente por L. racemosa hasta el 2145, momento a partir del cual A. germinans se haría dominante y el sistema lograría estabilizarse cerca de 100 años más tarde (Fig. 2b). La simulación para la estación LUN muestra la dominancia de L. racemosa en el mediano plazo, con un pico en el año 2069 cuando el AB de esta especie alcanzaría los 172,84 m2/ha y luego disminuiría hasta el año 2194 llegando a 29,69 m2/ha. Hacia el año 2081 el sistema mostrará aumento de R. mangle y A. germinans con descenso de L. racemosa hasta el final del periodo simulado llegando a 26,4 m2/ha, 13,67 m2/ha y 12,37 m2/ha, respectivamente (Fig. 2c). Para CGE se muestra un aumento del AB de R. mangle cercano a A. germinans en el año 2032; cuando esta última superaría a R. mangle y se mantendría dominante en el bosque por cerca de 200 años más. Hacia el año 2220 R. mangle volvería a aumentar su AB, mientras que A. germinans y L. racemosa tenderían a disminuir hasta el final del periodo de simulación con un AB de 38,56 m2/ha para R. mangle, 13,72 m2/ha para A. germinans y 0,60 m2/ha para L. racemosa (Fig. 2d). Finalmente, manteniendo las condiciones actuales, la estación RIN evidenciaría estabilidad hasta el año 2200 sin cambios significativos en las áreas basales, cuando R. mangle superaría el AB de L. racemosa. La estación mantendría las tendencias hasta el final del periodo de la simulación alcanzando 53,27 m2/ha, 0,038 m2/ha y 7,050 m2/ha, para A. germinans, L. racemosa y R. mangle, respectivamente (Fig. 2e).
Por otro lado, al evaluar las estaciones frente a un aumento anual constante en salinidad intersticial (0,5 m) el modelo predice reducciones severas en el AB de todas las especies llegando a valores muy bajos en el 2035 en todas las estaciones (Fig. 3). Ante este escenario, la estación KM22 disminuiría drásticamente a partir del año 2021 y registraría un AB total de 0,021m2/ha en el año 2035 con presencia de A. germinans y L. racemosa, mientras que; la especie R. mangle perdería toda su masa forestal en el 2025 (AB = 0m2/ ha) (Fig. 3a), Las estaciones CGE, ANE, LUN y RIN conservarían todas las especies luego de 20 años, sin embargo, llegarían a masas forestales muy reducidas (0,413m2/ ha; 0,754m2/ha; 0,045m2 ha y 0,677 m2/ ha, respectivamente). En todos los casos R. mangle sería la especie con menor Área Basal en cada estación, mientras que A. germinans sería la especie con mejor desempeño a excepción de la estación LUN donde L. racemosa continuaría siendo dominante al final del tiempo de simulación (0,0375 m2/ha) (Fig. 3). Las interpretaciones para el caso de las estaciones RIN y CGE y sus comparaciones con otras estaciones evaluadas en este estudio deberían tomarse con cuidado, teniendo en cuenta la sobreestimación del AB en magnitud existente que no se logró corregir durante el proceso de calibración.

DISCUSIÓN
Los trabajos de Twilley et al. (1998) tuvieron como supuesto principal la operación óptima de los caños reabiertos en el marco del proyecto PROCIENAGA; basados en ello y en datos de textura e hidrología preliminares, estimaron que la salinidad reduciría de manera lineal en el Complejo Pajarales en 80 unidades después de cinco años. Sin embargo, las observaciones en campo mostraron que para una diminución de 40 unidades de salinidad en la estación más cercana al complejo (LUN), se requirieron ocho años y que esta variable en ninguno de los casos siguió una tendencia lineal, en lugar de ello, ha disminuido de manera oscilatoria y nunca ha permanecido constante a lo largo del tiempo (Fig. 1). El comportamiento oscilatorio de la salinidad puede estar relacionado por un lado, con la falta de continuidad e ineficiencia en el mantenimiento de los canales del complejo estuarino que fueron reabiertos, y por otro, con los cambios en los fenómenos climáticos como El Niño y La Niña que modulan esta variable en la CGSM (Blanco et al. 2006). Adicionalmente, en varios casos el modelo subestimó los valores de Área Basal, sugiriendo que el modelo tal como fue inicialmente planteado es muy conservador y que se requieren mejoras en la calibración de sus parámetros.
Pocos proyectos de restauración en el mundo han contado con un programa de monitoreo a largo plazo como el de la de rehabilitación de la CGSM. Estos programas resultan vitales a la hora de evaluar el éxito de los proyectos planteados, entender la dinámica de los sistemas intervenidos y realizar su manejo a través de la observación de las respuestas o trayectorias del sistema y la modelación del mismo. En este trabajo, los ajustes implementados para calibrar el modelo fueron obtenidos a partir de los datos del monitoreo de las condiciones ambientales de la CGSM, logrando reproducir las tendencias generales de AB observadas para todas las especies monitoreadas, aunque fallaron en la reproducción de la magnitud para dos de las estaciones que recibieron menor impacto inicial (Fig.1). Esto último, podría sugerir que el modelo tiene mejor poder predictivo en situaciones de disturbio severo por salinidad, debido al mayor impacto que generan las funciones modificadoras del crecimiento bajo estas condiciones. Adicionalmente, este hecho resalta la heterogeneidad de condiciones que poseen los ecosistemas de manglar en la CGSM, por lo que un único modelo configurado con los mismos parámetros resulta ineficiente para todos los bosques de manglar en el complejo lagunar. De este modo, diferentes estrategias de manejo deberían considerar la variabilidad inmersa en estos sistemas boscosos, a fin de obtener respuestas más exitosas.
Dentro de los ajustes que más impactaron el modelo se destacan la modificación en el uso de las variables, cambiando funciones lineales por unas más plásticas definidas por el modelador, situación que resulta más cercana a la realidad, como por ejemplo en el caso de la salinidad.
El reclutamiento demostró ser una variable clave para ajustar las tendencias del modelo. En este estudio las constantes de reclutamiento fueron calculadas partiendo de observaciones realizadas en campo utilizando datos de 13 años. No obstante, se requirió realizar ajustes adicionales a ensayo y error para reproducir las tendencias observadas, lo que sugiere que los datos de campo no son suficientes para explicar el comportamiento del AB. Esto podría explicarse por la limitación del modelo FORMAN al no incluir explícitamente el efecto de la herbívora, la mortalidad, el hidroperíodo y los nutrientes en el desarrollo de nuevos individuos, asumiéndose que estos factores están implícitos en las tasas de reclutamiento calculadas (Chen & Twilley 1998). Otras limitaciones halladas en el modelo se relacionan con la tasa de mortalidad, al estar condicionada a una probabilidad independiente de su entorno, contemplando que solo un porcentaje de las plantas alcanzan la edad máxima de un árbol e ignorando factores como las enfermedades, la herbívora o la interacción de los árboles con otros organismos del ecosistema (Porté & Bartelink 2002).
A lo largo de la simulación, considerando la salinidad como disturbio principal, se observa un patrón unimodal del AB en los bosques de las cinco estaciones de muestreo. Este patrón podría ser explicado con base en la hipótesis de disturbio intermedio, desarrollada para explicar la riqueza de especies (Huston 1979). De tal modo, cuando la salinidad intersticial está en valores bajos, cerca de 25, hay coexistencia entre las tres especies de manglar con dominancia evidente de alguna de ellas. Cuando la salinidad se incrementa, el AB baja y aunque en tres de las cinco estaciones hay coexistencia de especies, A. germinans domina en la CGSM. Estos resultados en parte se explican por la diferencia de los umbrales de tolerancia a la salinidad descritos para cada especie, según los cuales R. mangle puede desarrollarse en un ambiente con máximo 70 unidades de salinidad, L. racemosa con máximo 85 unidades y A. germinans en ambientes de hasta 100 unidades de salinidad intersticial (Cintrón et al. 1978, Odum et al. 1982).
Si bien la salinidad es un regulador fundamental de la estructura de los bosques, otras variables extrínsecas e intrínsecas también son importantes en la estructuración de las comunidades.
La simulación a 500 años muestra el resultado de la sucesión ecológica a la que las especies responden. De acuerdo con Chen & Twilley (1998), L. racemosa domina en suelos fértiles con baja salinidad; sin embargo, su abundancia disminuye con el paso del tiempo, mientras que A. germinans aumenta y domina en zonas con salinidades altas. En la estación con mayor disturbio para 1998 (LUN) y en la estación donde se realizaron acciones de reforestación (CGE) dominaría R. mangle (Fig. 2). Este hecho sugiere que los sistemas de manglar gravemente impactados a pesar de ser recuperados, no recrean exactamente las características del sistema inicial o de referencia (RIN) y que la reforestación tiene un impacto importante en la modificación de las trayectorias naturales de los sistemas, trascendiendo en el tiempo (Rodríguez-R 2015).
Por otro lado, la disminución severa en menos de 20 años de la masa forestal de manglar evidenciada al simular un aumento progresivo de la salinidad intersticial (Fig. 2), demuestra que la salinidad es uno de los reguladores más importantes que influencia la estructura de estos bosques (Cintrón et al. 1978, Ball 1980, Castañeda-Moya et al. 2006) y ponen de manifiesto la vulnerabilidad de la CGSM a los incrementos en esta variable, destacando la importancia de garantizar los flujos hídricos en el sistema a fin de evitar aumentos en su concentración que son letales para el ecosistema.
En estos casos, los modelos proyectados en general demuestran una estancia más prolongada de A. germinans en el sistema (Fig. 3), hecho relacionado con la menor tolerancia a la salinidad y la baja capacidad de balance hídrico exhibida por R. mangle y L. racemosa. En las simulaciones estas dos especies tendieron a un menor desarrollo y a desaparecer más rápidamente que A. germinans, especie que es dominante en áreas donde la evaporación excede la precipitación y la salinidad es mayor a 120 unidades (Cintrón et al. 1978, Castañeda-Moya et al. 2006). Bajo estas condiciones de incremento progresivo de la salinidad, el sistema más vulnerable fue LUN, la estación más impactada inicialmente en la CGSM, mientras que la más resistente fue RIN, la estación que fue seleccionada como referencia de recuperación del sistema, por haber sido la menos impactada y la que exhibe la mayor madurez y estabilidad. Este hecho sugiere que los sistemas en mejor estado de conservación, son más resistentes a los cambios adversos del medio (Fig. 3).
Aunque con varios años de trabajo en la formulación y aplicación de Modelos Basados en Individuos (IBM), como el modelo FORMAN, su uso como herramienta para avanzar en el entendimiento de la dinámica de los bosques y su manejo aún es restringido, pues se requieren avances en la parametrización y contrastes con observaciones reales, entre otros (Berger et al. 2008). Esto destaca que el poder predictivo del modelo presentado con los ajustes realizados, dependerá de mantener en el futuro los parámetros y supuestos considerados y de una constante re-para-metrización cuando se requiera.
En el contexto local, ante la posibilidad de generar nuevas estrategias de rehabilitación para el humedal dadas las recientes afectaciones, resultaría de gran utilidad contar con un programa de modelación que facilite el manejo adaptativo. Particularmente se destaca la necesidad de: (1) seguir afinando el cálculo de las tasas de reclutamiento, (2) entender mejor los procesos de mortalidad del bosque y calcular sus tasas para cada especie, (3) incluir en el monitoreo la medición de nutrientes en suelo como Nitrógeno y Fósforo, que permitan contar con los datos necesarios para calibrar esta variable en el modelo FORMAN y entender su papel en los procesos de desarrollo y reclutamiento de los individuos, (4) comprender otros procesos que puedan afectar la dinámica del bosque como herbivoría, efecto de la distancia entre individuos, y finalmente (5) acoplar al modelo implementado modelos adicionales que permitan entender el efecto de la calidad del suelo y de las aguas, la hidrodinámica y operación de canales así como efectos del cambio climático global, aspectos que no son tenidos en cuenta directamente en FORMAN y que potenciarían su alcance y aplicación.
PARTICIPACIÓN DE LOS AUTORES
JARR concepción, análisis de datos y escritura del documento, JEMP concepción y escritura del documento, JMRP análisis de datos.
AGRADECIMIENTOS
Los autores agradecen a la Corporación Autónoma Regional del Magdalena (COR-PAMAG) por el apoyo en la financiación del proyecto "Monitoreo de las condiciones ambientales y los cambios estructurales y funcionales de las comunidades vegetales y de los recursos pesqueros durante la rehabilitación de la Ciénaga Grande de Santa Marta" y a todos los investigadores del Instituto de investigaciones marinas y costeras de Colombia INVEMAR que han hecho parte del proceso de obtención de los datos por más de 20 años. Así mismo, al Dr. Robert Twilley de la Universidad Estatal de Louisiana, quien suministró el código original del modelo FORMAN. Este proyecto fue financiado por el Banco de Proyectos de Inversión Nacional 2015 ejecutado por el programa Calidad Ambiental Marina del INVEMAR y hace parte de los resultados en el marco de la tesis de maestría titulada "Trayectorias de rehabilitación del bosque de manglar de la Ciénaga Grande de Santa Marta, luego de su reconexión con el río Magdalena" de la Universidad Nacional de Colombia, Sede Caribe.
LITERATURA CITADA
Ball, M. 1980. Patterns of secondary succession in a mangrove forest of southern Florida. Oecologia 44: 226-235.
Berger, U. & H. Hildenbrandt. 2000. A new approach to spatially explicit modeling of forest dynamics: spacing, ageing and neighborhood competition of mangrove trees. Ecological Modelling 132: 297-302.
Berger, U., M. Adams, V. Grimm & H. Hildenbrand. 2006. Modelling secondary succession of neotropical mangroves: Causes and consequences of growth reduction in pioneer species. Perspectives in Plant Ecology and Systematics 7: 243-252.
Berger, U., V.H. Rivera-Monroy, T. Doyle, F, Dahdouh-Guebas, N. Duke, M. Fontalvo-Herazo, H. Hildenbrand, N. Koedam, U. Mehlig, C. Piou & R.R. Twilley. 2008. Advances and limitations of individual-based models to analyze and predict dynamics of mangrove forests: A review. Aquatic Botany 89: 260-274.
Blanco, J., E. Viloria & J. Narváez. 2006. ENSO and salinity changes in the Ciénaga Grande de Santa Marta coastal lagoon system, Colombian Caribbean. Estuarine, Coastal and Shelf Science 66: 157-167.
Blanco, J. 2013. Modelos ecológicos: descripción, explicación y predicción. Ecosistemas 22(3): 1-5.
Borja, A., D. Dauer, E. Michael & C. Simens-tad. 2010. Medium- and long-term recovery of estuarine and coastal ecosystems: patterns, rates and restoration effectiveness ecosystems: patterns, rates and restoration effectiveness. Estuaries and Coasts 33: 1249-1260.
Botero, L. & H. Salzwedel. 1999. Rehabilitation of the Ciénaga Grande de Santa Marta, a mangrove-estuarine system in the Caribbean coast of Colombia. Ocean and Coastal Management 42: 243-256.
Botkin, D., J. James & J. Wallis. 1972. Some ecological consequences of a computer model of forest growth. Journal of Ecology 60 (3): 849-872.
Castañeda-M, E., V.H. Rivera-Monroy & R.R. Twilley. 2006. Mangrove zonation in the dry life zone of the Gulf of Fonseca, Honduras. Estuaries and Coasts 29: 751-764.
Cardona, P. & L. Botero. 1998. Soil characteristics and vegetation structure in a heavily deteriorated mangrove forest in the Caribbean coast of Colombia. Biotropica 30: 24-34.
Cardona-Olarte, P., K, Krauss & R.R. Twilley. 2013. Leaf gas exchange and nutrient use efficiency help explain the distribution of two neotropical mangroves under contrasting flooding and salinity, International Journal of Forestry Research, Article ID 524625: 1-10.
Campillo, F. & N. Champagnat. 2012. Simulation and analysis of an individual-based model for graph-structured plant dynamics. Ecological Modelling 234: 93-105.
Chen, R. & R.R. Twilley. 1998. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrients resources. Journal of Ecology 86: 37-51.
Cintrón, G., A. Lugo, D. Pool & G. Morris. 1978. Mangrove in arid environments in Puerto Rico and adjacent Islands Biotrópica 10: 110-121.
Cotes, G. 2004. Gestión institucional para la rehabilitación de la ecorregión Ciénaga Grande de Santa Marta y sus bosques de manglar. En: Garay, J., J. Restrepo, O. Casas, O. Solano y F. Newmark (eds.). Los manglares de la ecorregión Ciénaga Grande de Santa Marta: pasado, presente y futuro: 41-58 INVEMAR-Serie de publicaciones especiales No.11, Santa Marta.
Feller, I., K. Mckee, D. Whigham & J. O'neill. 2003. Nitrogen vs. phosphorus limitation across an ecotonal gradient in a mangrove forest. Biogeochemistry 62: 145-175.
Feller, I., C. Lovelock, U. Berger, K. Mckee, S. Joyce & M. Ball. 2010. Biocomplexity in mangrove ecosystems. Annual Review of Marine Science 2: 395-417.
Fitz, C., G. Kiker & J. Kim. 2011. Integrated ecological modeling and decision analysis within the Everglades Landscape. Critical Reviews in Environmental Science and Technology 41: 517-547.
Giraldo, E. 1995. Regeneración natural del manglar en el sector occidental (Isla Salamanca-complejo Pajarales) de la Ciénaga Grande de Santa Marta, Caribe Colombiano. Tesis Biología Marina. Universidad del Valle. Cali, 116 pp.
Hoyos, R., E. Urrego. & A. Lema. 2013. Respuesta de la regeneración natural en manglares del Golfo de Urabá (Colombia) a la variabilidad ambiental y climática intra-anual. Revista de Biología Tropical 61 (3): 1445-1461.
Huston, M. 1979. A general hypothesis of species diversity. American Naturalist. 113:81-101.
Invemar-Instituto De Investigaciones Marinas Y Costeras "José Benito Vives De Andreis". 2014. Monitoreo de las condiciones ambientales y los cambios estructurales y funcionales de las comunidades vegetales y de los recursos pesqueros durante la rehabilitación de la Ciénaga Grande de Santa Marta. INVEMAR. Santa Marta. 160 pp + anexos.
Martínez-E., A. Nejadhashemi, S. Woznicki & B. Love. 2014. Modeling the hydrological significance of wetland restoration scenarios. Journal of Environmental Management 133: 121-134.
Obeysekera, J., L. Kuebler, S. Ahmed, M. Chang, V. Engel, C. Langevin, E. Swain & Y. Wan. 2011. Use of hydrological and hydrodynamic modelling for ecosystem restoration. Critical Reviews in Environmental Science and Technology 41: 447-488.
Odum, W.E., C. Mcivor & T. Smith. 1982. The ecology of the mangroves of south Florida: a community profile. FWS/OBS-81/24. Washington, DC, United States Fish and Wildlife Service, Office of Biological Services.
Porté, A. & H. Bartelink. 2002. Modelling mixed forest growth: a review of models for forest management. Ecological Modelling 105: 141-188.
Rodhouse, T., K. Irvine, K. Vierling & L. Vierling. 2011. Estimating temporal trend in the presence of spatial complexity: a bayesian hierarchical model for a wetland plant population undergoing restoration. PLoS ONE 6 (12): 1-9.
Rodríguez-R, J.A. 2015. Trayectorias de rehabilitación del bosque de manglar de la Ciénaga Grande de Santa Marta, luego de su reconexión con el Río Magdalena. Tesis como requisito parcial para obtener el título de Magister en Ciencias, Biología, Línea Biología Marina. Universidad Nacional de Colombia, Santa marta. 96 pp.
Rivera-Monroy, V.H, J.E. Mancera-Pineda, R.R. Twilley, O. Casas, E. Castañeda, J. Restrepo, F. Daza, L. Perdomo, P. Reyes, E. Campos, M. Villamil, F. Pinto, P. Cardona, A. Vidal., W. Troncoso, D. Fonseca, E. Viloria, G. Sanchez, P. Rojas, J Narváez, J. Blanco, G. Ramírez, C. Henry, J. Fernández, F. Newmark, E. Carbono, C. Hernández, G. Cotes, H. Sanchez, Y. Herrera, A. María, R. Zuñiga, I. Acosta & A. Eguren. 2001. Estructura y función de un ecosistema de manglar a lo largo de una trayectoria de restauración: El caso de la región de la Ciénaga Grande de Santa Marta. University of Louisiana at Lafayette, Instituto de Investigaciones Marinas y Costeras (INVEMAR), Santa Marta. 285 pp.
Twilley, R.R., V.H. Rivera-Monroy, R. Chen & L. Botero. 1998. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Marine Pollution Bulletin 37: 404-419.
Twilley, R. R. & V. H. Rivera-Monroy. 2005. Developing performance measure of Mangrove wetlands using simulation models of hydrology, nutrient, biochemistry, and community dynamics. Journal of Coastal Research 21: 79-93.
Visser, J., S. Duke-Sylvester, J. Carter & W. Broussard. 2013. A computer model to forecast wetland vegetation changes resulting from restoration and protection in coastal Louisiana. En: Peyronnin, N. y Reed, D. (eds.). Louisiana's 2012 Coastal Master Plan Technical Analysis, Journal of Coastal Research, Special Issue No. 67: 51-59. Coconut Creek, Florida.
Wang, X., S. Shang, Z. Qu, T. Liu, A. Melesse & W. Yang. 2010. Simulated wetland conservation-restoration effects on water quantity and quality at watershed scale. Journal of Environmental Management 91: 1511-1525.
Recibido: 10/02/2016 Aceptado: 22/09/2016
Referencias
Ball, M. 1980. Patterns of secondary succession in a mangrove forest of southern Florida. Oecologia 44: 226-235.
Berger, U. & H. Hildenbrandt. 2000. A new approach to spatially explicit modeling of forest dynamics: spacing, ageing and neighborhood competition of mangrove trees. Ecological Modelling 132: 297-302.
Berger, U., M. Adams, V. Grimm & H. Hildenbrand. 2006. Modelling secondary succession of neotropical mangroves: Causes and consequences of growth reduction in pioneer species. Perspectives in Plant Ecology and Systematics 7: 243-252.
Berger, U., V.H. Rivera-Monroy, T. Doyle, F. Dahdouh-guebas, N. Duke, M. Fontalvoherazo, H. Hildenbrandt, N. Koedam, U. Mehlig, C. Piou & R.R. Twilley. 2008. Advances and limitations of individual-based models to analyze and predict dynamics of mangrove forests: A review. Aquatic Botany 89: 260-274.
Blanco, J., E. Viloria & J. Narváez. 2006. ENSO and salinity changes in the Ciénaga Grande de Santa Marta coastal lagoon system, Colombian Caribbean. Estuarine, Coastal and Shelf Science 66: 157-167.
Blanco, J. 2013. Modelos ecológicos: descripción, explicación y predicción. Ecosistemas 22(3): 1-5.
Borja, A., D. Dauer, E. Michael & C. Simenstad. 2010. Medium- and long-term recovery of estuarine and coastal ecosystems: patterns, rates and restoration effectiveness ecosystems: patterns, rates and restoration effectiveness. Estuaries and Coasts 33: 1249-1260.
Botero, L. & H. Salzwedel. 1999. Rehabilitation of the Ciénaga Grande de SantaMarta, a mangrove-estuarine system in the Caribbean coast of Colombia. Ocean and Coastal Management 42: 243-256.
Botkin, D., J. James & J. Wallis. 1972. Some ecological consequences of a computer model of forest growth. Journal of Ecology 60 (3):849-872.
Castañeda-M, E., V.H. Rivera-Monroy & R.R. Twilley. 2006. Mangrove zonation in the dry life zone of the Gulf of Fonseca, Honduras. Estuaries and Coasts 29: 751-764.
Cardona, P. & L. Botero. 1998. Soil characteristics and vegetation structure in a heavily deteriorated mangrove forest in the Caribbean coast of Colombia. Biotropica 30: 24–34.
Cardona-Olarte, P., K, Krauss & R.R. Twilley. 2013. Leaf gas exchange and nutrient use efficiency help explain the distribution of two neotropical mangroves under contrasting flooding and salinity, International Journal of Forestry Research, Article ID 524625: 1-10.
Campillo, F. & N. Champagnat. 2012. Simulation and analysis of an individual-based model for graph-structured plant dynamics. Ecological Modelling 234: 93-105.
Chen, R. & R.R. Twilley. 1998. A gap dynamic model of mangrove forest development along gradients of soil salinity and nutrients resources. Journal of Ecology 86: 37-51.
Cintrón, G., A. Lugo, D. Pool & G. Morris. 1978. Mangrove in arid environments in Puerto Rico and adjacent Islands Biotrópica 10: 110-121.
Cotes, G. 2004. Gestión institucional para la rehabilitación de la ecorregión Ciénaga Grande de Santa Marta y sus bosques de manglar. En: Garay, J., J. Restrepo, O. Casas, O. Solano y F. Newmark (eds.). Los manglares de la ecorregión Ciénaga Grande de Santa Marta: pasado, presente y futuro: 41-58 INVEMAR-Serie de publicaciones especiales No.11, Santa Marta.
Feller, I., K. Mckee, D. Whigham & J. O’neill. 2003. Nitrogen vs. phosphorus limitation across an ecotonal gradient in a mangrove forest. Biogeochemistry 62: 145-175.
Feller, I., C. Lovelock, U. Berger, K. Mckee, S. Joyce & M. Ball. 2010. Biocomplexity in mangrove ecosystems. Annual Review of Marine Science 2: 395-417.
Fitz, C., G. Kiker & J. Kim. 2011. Integrated ecological modeling and decision analysis within the Everglades Landscape. Critical Reviews in Environmental Science and Technology 41: 517-547.
Giraldo, E. 1995. Regeneración natural del manglar en el sector occidental (Isla Salamanca-complejo Pajarales) de la Ciénaga Grande de Santa Marta, Caribe Colombiano. Tesis Biología Marina. Universidad del Valle. Cali, 116 pp.
Hoyos, R., E. Urrego. & A. Lema. 2013. Respuesta de la regeneración natural en manglares del Golfo de Urabá (Colombia) a la variabilidad ambiental y climática intra-anual. Revista de Biología Tropical 61 (3): 1445-1461.
Huston, M. 1979. A general hypothesis of species diversity. American Naturalist. 113: 81-101.
Invemar-Instituto De Investigaciones Marinas y Costeras “José Benito Vives De Andreis”. 2014. Monitoreo de las condiciones ambientales y los cambios estructurales y funcionales de las comunidades vegetales y de los recursos pesqueros durante la rehabilitación de la Ciénaga Grande de Santa Marta. INVEMAR. Santa Marta. 160 pp + anexos.
Martínez-E., A. Nejadhashemi, S. Woznicki & B. Love. 2014. Modeling the hydrological significance of wetland restoration scenarios. Journal of Environmental Management 133: 121-134.
Obeysekera, J., L. Kuebler, S. Ahmed, M. Chang, V. Engel, C. Langevin, E. Swain & Y. Wan. 2011. Use of hydrological and hydrodynamic modelling for ecosystem restoration. Critical Reviews in Environmental Science and Technology 41: 447–488.
Odum, W.E., C. Mcivor & T. Smith. 1982. The ecology of the mangroves of south Florida: a community profile. FWS/OBS-81/24. Washington, DC, United States Fish and Wildlife Service, Office of Biological Services.
Porté, A. & H. Bartelink. 2002. Modelling mixed forest growth: a review of models for forest management. Ecological Modelling 105: 141-188.
Rodhouse, T., K. Irvine, K. Vierling & L. Vierling. 2011. Estimating temporal trend in the presence of spatial complexity: a Bayesian hierarchical model for a wetland plant population undergoing restoration. PLoS ONE 6 (12): 1-9.
Rodríguez-R, J.A. 2015. Trayectorias de rehabilitación del bosque de manglar de la Ciénaga Grande de Santa Marta, luego de su reconexión con el Río Magdalena. Tesis como requisito parcial para obtener el título de Magister en Ciencias, Biología, Línea Biología Marina. Universidad Nacional de Colombia, Santa marta. 96 pp.
Rivera-Monroy, V.H, J.E. Mancera-Pineda, R.R. Twilley, O. Casas, E. Castañeda, J. Restrepo, F. Daza, L. Perdomo, P. Reyes, E. Campos, M. Villamil, F. Pinto, P. Cardona, A. Vidal., W. Troncoso, D. Fonseca, E. Viloria, G. Sanchez, P. Rojas, J Narváez, J. Blanco, G. Ramírez, C. Henry, J. Fernández, F. Newmark, E. Carbonó, C. Hernández, G. Cotes, H. Sanchez, Y. Herrera, A. María, R. Zuñiga, I. Acosta & A. Eguren. 2001. Estructura y función de un ecosistema de manglar a lo largo de una trayectoria de restauración: El caso de la región de la Ciénaga Grande de Santa Marta. University of Louisiana at Lafayette, Instituto de Investigaciones Marinas y Costeras (INVEMAR), Santa Marta. 285 pp.
Twilley, R.R., V.H. Rivera-Monroy, R. Chen & L. Botero. 1998. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Marine Pollution Bulletin 37: 404-419.
Twilley, R. R. & V. H. Rivera-Monroy. 2005. Developing performance measure of Mangrove wetlands using simulation models of hydrology, nutrient, biochemistry, and community dynamics. Journal of Coastal Research 21: 79-93.
Visser, J., S. Duke-Sylvester, J. Carter & W. Broussard. 2013. A computer model to forecast wetland vegetation changes resulting from restoration and protection in coastal Louisiana. En: Peyronnin, N. y Reed, D. (eds.). Louisiana’s 2012 Coastal Master Plan Technical Analysis, Journal of Coastal Research, Special Issue No. 67: 51-59. Coconut Creek, Florida.
Wang, X., S. Shang, Z. Qu, T. Liu, A. Melesse & W. Yang. 2010. Simulated wetland conservation-restoration effects on water quantity and quality at watershed scale. Journal of Environmental Management 91: 1511-1525.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Juan F. Blanco-Libreros, Ricardo Álvarez-León. (2019). Mangroves of Colombia revisited in an era of open data, global changes, and socio-political transition: Homage to Heliodoro Sánchez-Páez. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 43(166), p.84. https://doi.org/10.18257/raccefyn.780.
2. Blanca Castellanos-Basto, Jorge Herrera-Silveira, Érick Bataller, Rodolfo Rioja-Nieto. (2021). Local Drivers Associated to Temporal Spectral Response of Chlorophyll-a in Mangrove Leaves. Sustainability, 13(9), p.4636. https://doi.org/10.3390/su13094636.
3. Jefferson Brooks, Miguel Chen Austin, Dafni Mora, Nathalia Tejedor-Flores. (2021). A Critical Review on Mathematical Descriptions to Study Flux Processes and Environmental-Related Interactions of Mangroves. Sustainability, 13(12), p.6970. https://doi.org/10.3390/su13126970.
4. Hudson DeYoe, Robert I. Lonard, Frank W. Judd, Richard Stalter, Ilka Feller. (2020). Biological Flora of the Tropical and Subtropical Intertidal Zone: Literature Review for Rhizophora mangle L.. Journal of Coastal Research, 36(4), p.857. https://doi.org/10.2112/JCOASTRES-D-19-00088.1.
5. Baltazar Sánchez-Díaz, Angel Sol-Sánchez, J. C. Cabrales-De La Cruz, Gloria Isela Hernández-Melchor, Lucy Dalia Aguilar-Sanchez. (2023). Análisis estadístico del crecimiento de mangle blanco (Laguncularia racemosa) durante la época de seca, Ejido la Solución Somos todos, Paraíso Tabasco. Rev. iberoam. bioecon. cambio clim., 9(18), p.2163. https://doi.org/10.5377/ribcc.v9i18.16617.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2016 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















