



Publicado
Efectos de la conectividad local sobre los ensambles de peces en una planicie de inundación tropical
DOI:
https://doi.org/10.15446/caldasia.v38n2.60920Palabras clave:
Río Sogamoso, San Silvestre, El Llanito, peces de ciénagas, pulso de inundación (es)Las lagunas de las planicies de inundación tropicales albergan alta diversidad biológica, son fundamentales para los ciclos de vida de muchos organismos y de importancia económica para las poblaciones locales. Este estudio evaluó el efecto de la conectividad local en los ensambles de peces en las lagunas San Silvestre (conectividad alterada) y El Llanito (conectividad no alterada), en la planicie de inundación del río Sogamoso, afluente del río Magdalena, Colombia. Se realizaron tres muestreos durante un ciclo hidrológico y se analizaron la abundancia, riqueza y equidad de los ensambles de peces y los aspectos fisicoquímicos de las lagunas. Se registraron 37 especies, 28 en El Llanito y 31 en San Silvestre. Aunque no se encontraron diferencias en los índices de riqueza y equitatividad, hay tendencias claras que indican menores valores en San Silvestre comparada con El Llanito. No hubo diferencias en la abundancia entre las lagunas, pero fue mayor y con poca variación en San Silvestre, mientras que en El Llanito hubo un pico en el último muestreo. En las dos lagunas las abundancias se concentraron en pocos taxones con variaciones temporales moderadas. En El Llanito la temperatura, el pH y la conductividad fueron mayores, mientras que en San Silvestre los valores fueron más elevados en profundidad y velocidad de la corriente. Los resultados pueden ser útiles para la conservación de los recursos ícticos dada su importancia social y ecológica para esta cuenca que está sometida a una gran presión por las actividades humanas.
doi: https://dx.doi.org/10.15446/caldasia.v38n2.60920
Ecología
Efectos de la conectividad local sobre los ensambles de peces en una planicie de inundación tropical
Effects of the local connectivity on fish assemblages in a tropical floodplain
Angela L. Gütiérrez-C.
Departamento de Biología, Universidad Nacional de Colombia. Bogotá D.C., Colombia. angelagutierrezc@gmail.com
Gabriel A. Pinilla-A.
Departamento de Biología, Universidad Nacional de Colombia. Edif. 405, of. 205, Bogotá D.C., Colombia. gapinillaa@unal.edu.co
RESUMEN
Las lagunas de las planicies de inundación tropicales albergan alta diversidad biológica, son fundamentales para los ciclos de vida de muchos organismos y de importancia económica para las poblaciones locales. Este estudio evaluó el efecto de la conectividad local en los ensambles de peces en las lagunas San Silvestre (conectividad alterada) y El Llanito (conectividad no alterada), en la planicie de inundación del río Sogamoso, afluente del río Magdalena, Colombia. Se realizaron tres muestreos durante un ciclo hidrológico y se analizaron la abundancia, riqueza y equidad de los ensambles de peces y los aspectos fisicoquímicos de las lagunas. Se registraron 37 especies, 28 en El Llanito y 31 en San Silvestre. Aunque no se encontraron diferencias en los índices de riqueza y equitatividad, hay tendencias claras que indican menores valores en San Silvestre comparada con El Llanito. No hubo diferencias en la abundancia entre las lagunas, pero fue mayor y con poca variación en San Silvestre, mientras que en El Llanito hubo un pico en el último muestreo. En las dos lagunas las abundancias se concentraron en pocos taxones con variaciones temporales moderadas. En El Llanito la temperatura, el pH y la conductividad fueron mayores, mientras que en San Silvestre los valores fueron más elevados en profundidad y velocidad de la corriente. Los resultados pueden ser útiles para la conservación de los recursos ícticos dada su importancia social y ecológica para esta cuenca que está sometida a una gran presión por las actividades humanas.
Palabras clave. Río Sogamoso, San Silvestre, El Llanito, peces de ciénagas, pulso de inundación.
ABSTRACT
Lagoons of tropical floodplains harbor high levels of biological diversity, play a fundamental role in the life cycles of many organisms and are of great economic importance to local people. This study evaluated the effect of local connectivity on fish assemblages of the San Silvestre (altered connectivity) and El Llanito (unaltered connectivity) lagoons of the Sogamoso River floodplain (Magdalena River basin, Colombia). Samples were collected at three different times during the course of the hydrological cycle and abundance, richness and evenness of fish assemblages were analyzed, along with physicochemical aspects of the lagoons. A total of 37 species were recorded, 28 in El Llanito and 31 in San Silvestre. None of the richness and evenness indices showed statistically significant differences; however clear trends towards higher dominance, lower diversity, and less evenness in San Silvestre compared to El Llanito were observed. There was no significant difference in abundance between the lagoons, but it was greater with lower variation in San Silvestre, while in El Llanito there was a peak in the last round of sampling. Abundance in the two lagoons was concentrated in a few taxa and showed moderate variation over the sampling periods. Temperature, pH, and conductivity values were higher in El Llanito lagoon, while the San Silvestre lagoon had greater depth and flow velocity. This study provides useful information for the conservation of fish resources in the Magdalena River basin, a region of great social and ecological importance that is under considerable pressure from human activities.
Key words. Sogamoso River, San Silvestre, El Llanito, swamps fish, flood pulse.
INTRODUCCIÓN
Las lagunas de las planicies de inundación son áreas fundamentales para la regulación hídrica debido a que amortiguan el incremento de los niveles de los ríos, los cuales están asociados a las precipitaciones periódicas en las cabeceras de las cuencas (Galvis & Mojica 2007, Montoya & Aguirre 2009, Junk & Wantzen 2004). Este proceso dinámico se manifiesta en la planicie a través del pulso de inundación y de la variación estacional de la conectividad entre los diferentes sistemas ecológicos (Thomaz et al. 2007, Agostinho et al. 2009). A pesar de que se sabe que la conectividad derivada del pulso de inundación juega un papel determinante y estructurador en las comunidades de estos ecosistemas (Lasne et al. 2007, Thomaz et al. 2007, Agostinho et al. 2009, Hurd et al. 2016), en Colombia aún existen pocos estudios que evalúen este efecto.
Durante el periodo de caudales altos, las lagunas de inundación se convierten en depósitos temporales de agua que posteriormente retorna a los ríos en el periodo de sequía. El carácter bidireccional del agua, entre río y lagunas, crea un flujo de nutrientes que enriquece la oferta alimenticia de estos ecosistemas lénticos (Winemiller & Jepsen 1998). Esto los convierte en ambientes de gran biodiversidad, en áreas de desove y apareamiento, así como en lugares obligados de visita para especies migratorias y reofílicas de importancia pesquera. Por esta razón, las lagunas de inundación ofrecen alimento y refugio a los peces mientras llega el periodo de aguas bajas, época en la cual se desplazan al canal principal del río durante sus migraciones reproductivas o alimenticias (Winemiller & Jepsen 1998, Claro et al. 2004, Agostinho et al. 2007, Lasne et al. 2007, Casatti et al. 2009, Burgess et al. 2012). La magnitud de las migraciones depende de la especie, algunas pueden desarrollar todo su ciclo de vida dentro del canal principal del río, otras permanecen en dicho canal durante la época de sequía y pasan a la planicie durante la inundación, y otras permanecen dentro de las lagunas durante la sequía y entran al río durante la inundación (Winemiller & Jepsen 1998, Agostinho et al. 2007, Galvis & Mojica 2007). Por lo tanto, estas lagunas cumplen funciones ecológicas únicas que en la actualidad no han sido diagnosticadas exhaustivamente en Colombia.
Una de las principales amenazas a la bio-diversidad de estos ecosistemas acuáticos continentales es la introducción de especies. La cuenca del río Magdalena es la más amenazada del país con 96 especies de peces introducidas y trasplantadas (IDEAM 2004). La presencia de varias especies de cíclidos en los ambientes naturales afecta las pesquerías de especies nativas (Gutiérrez et al. 2010). Esta situación es particularmente importante en la zona de estudio, pues se conoce ampliamente que las lagunas de los planos de inundación son fuente de alimento y recursos financieros para las poblaciones humanas locales, derivadas de la pesca artesanal, la caza y los cultivos de tierras inundables (Rincón & Rivas 2002, Patrón 2004, Galvis & Mojica 2007). A pesar de su importancia, presentan serios problemas relacionados con actividades pecuarias, agrícolas, urbanísticas y construcción de represas, que llevan a la deforestación de la vegetación ribereña, la contaminación de suelos y aguas, el aumento de sedimentos y la pérdida de conectividad hídrica (Lasne et al. 2007). Todos estos factores generan un acelerado deterioro de la diversidad y la funcionalidad de estos ecosistemas acuáticos, así como de los sistemas terrestres que los circundan (Díaz-Granados et al. 2001, Patrón 2004, Granado-Lorencio et al. 2005).
En este contexto, el presente estudio evaluó el efecto del grado de conectividad local en los ensambles de peces en dos lagunas de una planicie de inundación tropical, una de ellas con conectividad no alterada y la otra sí. Con este objetivo, se analizaron atributos de la estructura de las comunidades ícticas, tales como riqueza, abundancia y equidad de especies, y variables ambientales para establecer las diferencias entre los dos sistemas. La hipótesis planteada fue que la pérdida de conectividad local afecta negativamente la estructura de la comunidad de peces debido a la pérdida de vías de dispersión entre las lagunas y otros cuerpos de agua de la planicie.
MATERIALES Y MÉTODOS
Área de estudio
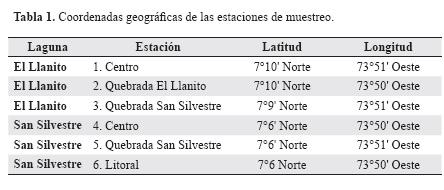
Las lagunas El Llanito (807 ha; 7°10' Norte, 73°51' Oeste) y San Silvestre (1.849 ha; 7°5' Norte, 73°48' Oeste) pertenecen a la planicie de inundación del río Sogamoso (Castellanos et al. 2011) y se caracterizan por tener aguas turbias (Mojica et al. 2006). Estas dos lagunas, también conocidas como ciénagas, tenían conexiones naturales que las comunicaban con el canal principal del río a través del caño San Silvestre. No obstante, en 1978 la construcción de una compuerta interrumpió dicha conexión en la laguna San Silvestre, alterando así los cambios de nivel de los periodos de inundación y sequía derivados del ciclo hidrológico (Pava et al. 2006). El río Sogamoso se forma por la confluencia de los ríos Chicamocha y Suárez y desemboca en el río Magdalena (Castellanos et al. 2011). El río recorre un valle profundo y estrecho con dirección norte-este, con substrato rocoso y alta corriente. En la región media, el río es trenzado pero cerca de la desembocadura el canal principal tiene playas de arena fina y baja pendiente (Ramírez & Pinilla 2012) (Figura 1).

En esta región ocurre un periodo de sequía y aguas bajas de diciembre a marzo, seguido de un periodo de lluvias y aguas altas de abril a mayo. Posteriormente se presenta un periodo de aguas en descenso de junio a agosto y finalmente una segunda etapa de aguas altas de septiembre a noviembre (Granado-Lorencio et al. 2005, Guzmán 2005).
Muestreos
Los muestreos se realizaron en junio (aguas en descenso), noviembre (aguas altas) y diciembre (aguas bajas) de 2009. En cada campaña se ejecutaron cuatro jornadas de pesca de cuatro horas cada una en tres puntos de cada laguna, dos en la mañana y dos en el crepúsculo. Los puntos de pesca se seleccionaron considerando diferentes microambientes dentro las lagunas (Montoya & Aguirre 2009) (Tabla 1). En la laguna El Llanito se hicieron las capturas en el centro del sistema y en dos sitios correspondientes a las desembocaduras de las quebradas El Llanito y San Silvestre, esta última une las dos lagunas analizadas. En la laguna San Silvestre se tomó un punto cercano a la desembocadura de la quebrada San Silvestre, próximo a la compuerta que limita la conectividad de la laguna, otro en el centro del cuerpo de agua y otro próximo a región litoral. Las capturas se realizaron con atarraya de 2,5 cm de ojo de malla. Todos los ejemplares capturados se identificaron mediante manuales y soporte de los pescadores de la región y se liberaron, a excepción de aquellos de taxonomía dudosa, en cuyo caso se fijaron unos pocos individuos con una solución de formaldehido al 10% y se trasladaron al Laboratorio de Ecología de la Universidad Nacional de Colombia, Bogotá. Adicionalmente, se registraron in situ la temperatura del agua, el pH y la conductividad eléctrica (µScm-1) con equipos de campo Extech-EC500 y Extech-DO600. También se midieron la profundidad (cm) y la velocidad de la corriente superficial (ms-1).
Para la identificación del material biológico se siguió la clasificación propuesta por Nelson (2006), utilizando claves especializadas (Schultz 1944, Dahl 1971, Géry 1977, Maldonado et al. 2005, Covain & Fisch-Muller 2007). Los registros de abundancia se estandarizaron como datos de captura por unidad de esfuerzo (CPUE), es decir igual a individuos capturados en 4 horas/12,56 m2 de red.
Análisis de datos
Con el fin de describir la estructura de los ensambles de peces, se determinaron los índices de riqueza o número de especies, abundancia y equidad J', para lo cual se empleó el programa Past 3.09 (Hammer et al. 2001). Posteriormente, estos valores se compararon mediante un análisis descriptivo, debido a que la baja cantidad de muéstreos no permitió alcanzar los requerimientos necesarios para realizar pruebas parámetricas. De la misma manera, no se pudo aplicar una técnica como PERMANOVA dada la baja cantidad de datos. Finalmente, para estudiar el comportamiento de los parámetros fisicoquímicos de las lagunas se tomaron todos los datos de las variables medidas para realizar un análisis de componentes principales (ACP), en el que se seleccionaron para la interpretación los componentes con los mayores autovalores generados por el modelo aleatorio de "broken-stick" (Jackson 1993) con el software PCORD 6 (McCune & Mefford 2011).
RESULTADOS
Estructura y composición
Se capturaron 2391 individuos, distribuidos en 37 especies, 28 en El Llanito y 31 en San Silvestre, seis órdenes, cinco en El Llanito y seis en San Silvestre, 16 familias, catorce en El Llanito y quince en San Silvestre y 32 géneros, 26 en El Llanito y 27 en San Silvestre (Tabla 2). Las familias mejor representadas fueron Characidae y Pimelodidae. La familia Cichlidae estuvo compuesta por tres especies. Siete familias presentaron solo dos especies (Curimatidae, Prochilodontidae, Anostomidae, Loricariidae, Auchenipteridae, Sternopygidae y Poeciliidae) y cinco estuvieron representadas por una sola especie (Potramotrygonidae, Ctenoluci-dae, Erythrinidae, Heptapteridae, Doradidae). Las especies más abudantes y frecuentes fueron los carácidos Astyanax magdalenae Eigenmann & Henn, 1916 y Leporinus muyscorum Steindachner, 1901, y los siluriformes Pimelodus blochii Valenciennes, 1840 y Trachelyopterus insignis (Steindachner, 1878).
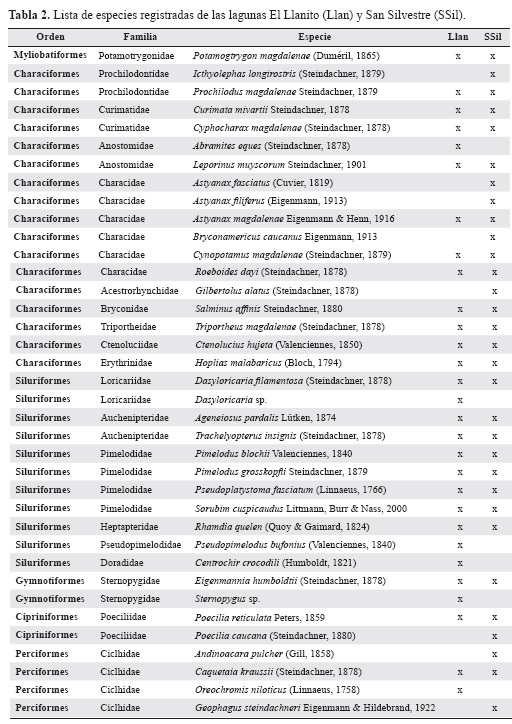
La abundancia total presentó patrones diferentes entre las lagunas. En El Llanito la abundancia se duplicó en el último muestreo frente a los periodos anteriores. En San Silvestre la abundancia fue relativamente mayor y más homogénea a lo largo de los tres muestreos (Figura 2). En El Llanito la mayor cantidad de individuos por especie estuvo concentrada en ocho taxones, cuyas abundancias tendieron a aumentar a lo largo de los muestreos, siendo más altas en la época de mayor sequía, es decir en diciembre (Figura 3 a). Las abundancias de las especies mejor representadas (A. magdalenae, L. muyscorum, P. blochii y T. insignis), tuvieron una amplia variación entre muestreos (Figura 3b), con un claro aumento en diciembre. Solo la arenca (Triportheus magdalenae (Steindachner, 1878)) evidenció un notable aumento en noviembre. Otras especies alcanzaron cantidades considerables, tales como Cy-phocharax magdalenae (Steindachner, 1878) y Prochilodus magdalenae (Stein-dachner, 1879), pero sus abundancias variaron moderadamente a lo largo de los muestreos. Las demás especies tuvieron bajas abundancias con poca variación entre muestreos. En la laguna San Silvestre la mayor parte de las especies tuvieron bajas abundancias que tendieron a disminuir a lo largo de los muestreos. En la época de menor nivel del agua, es decir diciembre, se capturaron menos ejemplares de la mayoría de especies (Figura 3c), a excepción de T. magdalenae que presentó un aumento en las capturas. Otras especies como Roeboides dayi (Steindachner, 1878) y C. magdalenae tuvieron abundancias moderadas con un pico en junio que son las aguas en descenso. Por su parte, Andinoacara pulcher (Gill, 1858) y P. magdalenae aumentaron en noviembre y diciembre, respectivamente, aunque con bajas abundancias (Figura 3d).


La mayor riqueza ocurrió en junio y la menor en diciembre en ambos ecosistemas (Figura 4a). La equidad en El Llanito tuvo valores más altos en comparación con San Silvestre, en los dos casos presentó un aumento en el periodo de aguas altas, es decir noviembre (Figura 4b).

Parámetros ambientales
Se seleccionaron los ejes uno y dos del ACP para la interpretación de las variables abióticas. Estos ejes explicaron en conjunto el 64,3% de la varianza acumulada (Figura 5). Los parámetros asociados positivamente al eje 1 (autovalor del eje uno = 2,007, explicación del 40,14%) fueron la temperatura del agua, la conductividad eléctrica y el pH, y negativamente la velocidad de la corriente y la profundidad. Al eje dos (autovalor del eje dos = 1,207, explicación de 24,15%) se asociaron positivamente la conductividad eléctrica y negativamente la temperatura del agua y la profundidad. El ordenamiento indica que desde el punto de vista fisicoquímico, las dos lagunas son diferentes. Mientras que en El Llanito la temperatura, el pH y la conductividad fueron más altos, la laguna San Silvestre tuvo mayores valores de profundidad y de velocidad de la corriente (Figura 5). Otro aspecto importante que se observa en el ACP es la tendencia a una mayor dispersión de los puntos correspondientes a la laguna El Llanito, mientras que aquellos pertenecientes a San Silvestre están más congregados, lo cual es indicio de una menor variabilidad ambiental en este último ecosistema.

DISCUSIÓN
Las lagunas de las planicies de inundación de los ríos tropicales son ecosistemas esenciales para la manutención de la diversidad de diferentes grupos de flora y fauna (Junk & Wantzen 2004, Lasne et al. 2007, Agostinho et al. 2009, Fearnside 2014). Estos ambientes son fundamentales en el desarrollo de los ciclos de vida de los peces, especialmente en los procesos de alimentación (Winemiller & Jepsen 1998), de reproducción y de protección de sus larvas y juveniles (Claro et al. 2004, Lasne et al. 2007, Casatti et al. 2009, Montoya & Aguirre 2009, Granado-Lorencio et al. 2012b).
La riqueza de especies encontrada durante este estudio (El Llanito 28 spp y San Silvestre 31 spp), coincide con Granado-Lorencio et al. (2012a, 2012b) quienes reportaron un promedio de 23 especies para las lagunas de la planicie de inundación del río Magdalena y 31 para El Llanito. La mayor riqueza de la laguna San Silvestre puede deberse a su mayor tamaño, pues hay una relación positiva entre el área y la riqueza de especies (Gaston 2000, Tetleta-Rangel et al. 2013). Las abundancias en El Llanito tendieron a ser más variables que en San Silvestre. Taxones como C. magdalenae, T. insignis y Caquetaia krausii (Steindachner, 1878), consideradas especies no migratorias, presentaron abundancias moderadas con poca variación. Especies migratorias como P. magdalenae, Curimatta mivartii Steindachner, 1878, L. muyscorum, T. magdalenae y P. blochii (Granado-Lorencio et al. 2005, Granado-Lorencio et al. 2012b), tuvieron las mayores abundancias y las variaciones temporales más amplias. Estos cambios probablemente estén asociados a las fluctuaciones de nivel del agua en este sistema, producto de su mayor conectividad, lo que permite que el ciclo hidrológico desencadene los movimientos migratorios en los meses de sequía (Agostinho et al. 2004, Thomaz et al. 2007, Luz-Agostinho et al. 2008, Montoya & Aguirre 2009).
En la laguna San Silvestre por su parte, muchas especies tuvieron bajas cantidades y poca variación entre los muestreos. Solo tres de ellas mostraron abundancias altas y cambios importantes en su densidad; ellas son: T. magdalenae, de hábitos migratorios, y R. dayi y A. pulcher, especies pequeñas que contribuyeron con abundancias moderadamente elevadas, siendo menores en la época de sequía. Al parecer, durante los periodos de aguas altas la disgregación de sus poblaciones en un mayor volumen de agua las hace menos vulnerables a la depredación (Winemiller & Jepsen 1998, Luz-Agostinho et al. 2008). Las especies migratorias P. magdalenae y A. magdalenae y la sedentaria C. magdalenae, tuvieron abundancias intermedias, con variaciones moderadas. Si esta laguna tuviera una conectividad más activa, se esperaría que los peces migratorios mostraran fluctuaciones mucho más amplias.
De las 37 especies registradas en este estudio, nueve (24%) presentan algún grado de amenaza (Mojica et al. 2012). Varias de ellas son de interés para la población local, ya sea para la venta o para autoconsumo. Dentro de las especies registradas como Casi Amenazadas (NT) están Cynopotamus magdalenae (Steindachner, 1879) y Pota-motrygon magdalenae (Duméril, 1865); las Vulnerables son Abramites eques (Steindachner, 1878), Ageneiosus pardalis (Lütken, 1874), C. mivartii, L. muyscorum y Salminus affinis (Mojica et al. 2012). Las especies P. magdalenae y Pseudoplatystoma magdaleniatum (Buitrago-Suárez & Burr, 2007), se catalogan como Vulnerables (A2c,d) y en Peligro Crítico (A1d) (ver explicación de las abreviaturas en Mojica et al. 2012). Estas especies se pueden considerar como extremadamente sensibles, tanto por sobrepesca como por los efectos negativos de la pérdida de conectividad entre los ambientes de la planicie de inundación, debido a que los aspectos reproductivos y tróficos de sus ciclos de vida dependen de la ocurrencia de los periodos de inundación. Estos pulsos son necesarios para que sus migraciones puedan darse (Granado-Lorencio et al. 2005, Galvis et al. 2007, Lasne et al. 2007, Hurd et al. 2016).
A pesar de la alteración de la conexión natural de la laguna San Silvestre, no fue posible establecer diferencias consistentes entre los dos cuerpos de agua en los índices ecológicos de riqueza, abundancia o equidad. En el presente estudio los índices ecológicos no revelaron efectos sobre el ensamble de peces por la falta de conectividad en San Silvestre. No obstante, cuando se observan los cambios en los taxones individuales, se puede ver que en el sistema con menor conectividad las especies representativas fueron diferentes y los cambios en abundancia fueron menores, debido muy probablemente a una menor variación fisicoquímica, como lo evidenció el ACP.
Existen numerosos trabajos que han resaltado el papel fundamental que tiene la conectividad entre los ambientes de las planicies de inundación en los ciclos de vida de los peces (p.e, reproducción, alimentación, dispersión en los momentos de inundación) (Agostinho et al. 2004, Lasne et al. 2007, Thomaz et al. 2007, Luz-Agostinho et al. 2008, Agostinho et al. 2009, Martin et al. 2011, Burgess et al. 2012, Hurd et al. 2016). Por lo tanto, existe una alta probabilidad de que la desconexión de San Silvestre con el río Sogamoso provoque cambios significativos en la estructura del ensamble de peces, los cuales serían visibles en una escala temporal mayor.
A pesar de hallarse mayor riqueza en la laguna de San Silvestre, tuvo menores valores de equidad debido a la marcada dominancia de algunas especies. En El Llanito se encontró un comportamiento opuesto, con baja dominancia de algunas especies, lo que podría indicar un efecto homogeneizador de la conectividad con el río (Thomaz et al. 2007, Hurd et al. 2016). En general, la riqueza de especies de peces está influenciada por varios factores, entre los que se encuentran aspectos históricos y biogeográficos, la conectividad hídrica, las condiciones físicas y químicas de las lagunas y el tamaño de los ambientes acuáticos (Granado-Lorencio et al. 2005, Lasne et al. 2007, Granado-Lorencio et al. 2012a). Nuestros resultados sugieren que la falta de conectividad reduce la riqueza debido a una mayor homogenización ambiental y a la imposibilidad para lo peces de desplazarse entre los cuerpos de agua de la planicie. La menor equidad en San Silvestre indica que la compuerta modifica los efectos del pulso de inundación reduciendo la heterogeneidad ambiental y el flujo bidireccional de peces entre la laguna y el río, lo que podría favorecer a las especies más competitivas y reducir la diversidad local (Thomaz et al. 2007, Agostinho et al. 2009, Martin et al. 2011).
La falta de conectividad puede favorecer a algunas especies y perjudicar a otras, pues las más sensibles no se pueden desplazar en búsqueda de cuerpos de agua con mejores condiciones ambientales (Heino et al. 2015) o con interacciones bióticas menos estresantes (Hurd et al. 2016).
A pesar de que las abundancias tampoco tuvieron diferencias significativas en las dos lagunas, el incremento de individuos en El Llanito durante la época de aguas bajas se podría deber a la reducción del nivel de agua. Los cambios de nivel son notorios en esta laguna debido a su conectividad con el sistema fluvial. El aumento de la abundancia en diciembre muestra principalmente un efecto de concentración de los organismos durante el periodo seco, lo que favorece su captura (Winemiller & Jepsen 1998, Luz-Agostinho et al. 2008). En San Silvestre la compuerta atenúa los cambios estacionales de nivel del río, lo que se traduce en abundancias relativamente constantes a lo largo del año.
Las características fisicoquímicas sugieren que la alteración de la conexión entre la laguna San Silvestre y el canal del río mantiene condiciones limnológicas relativamente constantes a lo largo del ciclo hidrológico. En términos ambientales, esto significa que San Silvestre no presenta patrones fisicoquímicos estacionales, producidos por la alternancia de periodos secos y de inundación, como sí ocurre en otros sistemas lénticos con mejor conectividad (Thomaz et al. 2007, Agostinho et al. 2009). En la laguna El Llanito por su parte, se observa una mayor heterogeneidad ambiental, determinada por las diferencias en los flujos de agua de los distintos momentos del ciclo hidrológico. Esta variabilidad hidrológica favorece una comunidad íctica más diversa y equitativa (Thomaz et al. 2007).
Los resultados aportan información importante sobre la ecología de las lagunas inundables de Colombia, en especial de las que se localizan en la cuenca del río Magdalena. Dicha cuenca representa el 25% del territorio nacional, en el cual está asentado el 80% de la población, lo que genera una presión humana intensa y permanente sobre sus recursos naturales (Galvis & Mojica 2007). Esta cuenca constituye un ejemplo claro del conflicto entre el desarrollo y la conservación de la biodiversidad. La cuenca ha experimentado una disminución de aproximadamente el 90% de las capturas de peces desde la década de los años setenta hasta la fecha (Galvis & Mojica 2007). Una de las consecuencias más evidentes de esta disminución ha sido la inclusión en las pesquerías de especies que antiguamente eran desechadas debido a su tamaño o sabor y que actualmente generan interés comercial para los pescadores, como es el caso del T. magdalenae y L. muyscorum.
Se puede concluir que la laguna San Silvestre, que tiene conexión limitada, presenta menor equidad de especies y menor variación ambiental. Es de suma importancia comprobar en futuros estudios estas diferencias con muestreos más intesivos y prolongados en el tiempo que permitan evaluar el efecto sostenido de la disminución de la conectividad en los ensambles de peces. De esta manera se tendría certeza sobre la aparente disminución de la diversidad íctica de San Silvestre, la cual a largo plazo puede generar extinciones locales de especies que históricamente dependen de la intermitencia natural del pulso de inundación, tanto para su reproducción y alimentación, como para su dispersión (Burgess et al. 2012, Granado-Lorencio 2012a, 2012b, Hurd et al. 2016).
PARTICIPACIÓN DE LOS AUTORES
ALGC concepción y diseño, toma de datos, ALGC y GAPA análisis de datos, escritura del documento.
AGRADECIMIENTOS
Los autores agradecen a la Dirección de Investigaciones y a la Facultad de Ciencias de la Universidad Nacional de Colombia, Sede Bogotá, por la financiación. A Angélica Ramírez por el apoyo y las contribuciones durante la ejecución y análisis de los resultados del proyecto. Al grupo de investigación Biodiversidad, Biotecnología y Conservación de Ecosistemas por el apoyo logístico.
LITERATURA CITADA
Agostinho, A.A., L.C. Gomes, S. Veríssimü & E.K. Okada. 2004. Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11-19.
Agostinho, A.A., F.M. Pelicice, A.C. Petry, L.C. Gomes & H.F. Júlio-Jr. 2007. Fish diversity in the upper Paraná River basin: habitats, fisheries, management and conservation. Aquatic Ecosystem Health & Management 10(2):174-186.
Agostinho, A.A., C.C. Bonecker, & L.C. Gomes. 2009. Effects of water quantity on connectivity: the case of the upper Paraná River floodplain. Ecohydrology and Hydrobiology 9(1): 99-113.
Burgess, O.T., W.E. Pine Iii & S.J. Walsh. 2012. Importance of floodplain connectivity to fish populations in the Apalachicola river, Florida. River Research and Applications 29(6): 718-733.
Casatti, L., J.L. Veronezi Júnior & C.P. Ferreira. 2009. Diet of the armored catfish Aspidoras fuscoguttatus (Ostariophysi, Callichthyidae) in streams with different limnological and structural features. Biota Neotropica 9(1): 113-21.
Castellanos-M, Ca., N. Chamarravi, E. Castañeda, F. Galvis & E. Cepeda. 2011. Variación espacio temporal de la cobertura vegetal en la microcuenca Ciénaga El Llanito, Santander, Colombia. Boletín Científico Centro de Museos, Museo de Historia Natural 15(2): 60-68.
Clarü-Jr, L., E. Ferreira, J. Züanon & C. Araüjo-Lima. 2004. Efeito da floresta alagada na alimentação de três espécies de peixes onívoros em lagos de várzea da Amazônia Central, Brasil. Acta Amazônica 34(1): 133-137.
Covain, R. & S. Fisch-Müller. 2007. The genera of the Neotropical armored catfish subfamily Loricariinae (Siluriformes: Loricariidae): a practical key and synopsis. Zootaxa 1462: 1-40.
Dahl, G. 1971. Los peces del norte de Colombia. Ministerio de Agricultura, Instituto de los Recursos Naturales Renovables INDERENA, Bogotá. 391 pp.
Díaz-Granados, M., L. Camacho & A. Maestre. 2001. Modelación de balances hídricos de ciénagas fluviales y costeras colombianas. Revista de Ingeniería. Universidad de los Andes. Facultad de Ingeniería 13: 12-20.
Fearnside, P.M. 2014. Impacts of Brazil's Madeira River Dams: Unlearned lessons for hydroelectric development in Amazonia. Environmental Science & Policy 38: 164-172.
Galvis, G. & J.I. Mojica. 2007. The Magdalena River fresh water fishes and fisheries. Aquatic Ecosystem Health & Management 10(2): 127-139.
Gaston, K.J. 2000. Global patterns in biodiversity. Nature 405: 220-227.
Géry, J. 1977. Charachoids of the World. T. F. H. Publications, New Jersey. 294 pp.
Granado-Lorencio, C., C.R.M. Araüjo-Lima & J. Lobôn-Cerviá. 2005. Abundance-distribution relationships in fish assembly of the Amazonas floodplain lakes. Ecography 28(4): 515-520.
Granado-Lorencio, C., A. Gülfo, F. Alvarez, L.F. Jiménez-Segura, J.D. Carvajal-Quintero & A. Hernández-Serna. 2012a. Fish assemblages in floodplain lakes in a Neotropical river during the wet season (Magdalena River, Colombia). Journal of Tropical Ecology 28: 271-279.
Granado-Lorencio, C., A. Hernández-Serna, J.D. Carvajal, L.F. Jiménez-Segura, A. Gulfo & F. Álvarez. 2012b. Regionally nested patterns of fish assemblages in floodplain lakes of the Magdalena River (Colombia). Ecology and Evolution 2(6): 1296-303.
Gutiérrez, F.P., C.A. Lasso, P. Sánchez-Duarte & D.L Gil. 2010. Análisis de riesgo para especies acuáticas continentales y marinas. En: Baptiste, M.P., N. Castaño, D. Cárdenas, F.P. Gutiérrez, D. Gil y C.A. Lasso (eds) Análisis de riesgo y propuesta de categorización de especies introducidas en Colombia. 73-148 pp. Instituto de Investigación de Recursos Biológicos «Alexander von Humboldt», Bogotá, Colombia.
Guzmán, A. 2005. Aspectos históricos y técnicos del río Magdalena. Una visión sobre experiencias en el río. Hojas Universitarias 57: 35-44.
Hammer, 0., D.A.T. Harper & P.D. Ryan. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1): 1-9.
Heino, H., A.S. Melo, T. Siqueira, J. Soininen, S. Valanko & L.M. Bini. 2015. Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshwater Biology 60(5): 845-869.
Hurd, L.E., R.G.C. Sousa, F.K. Siqueira-Souza, G.J. Cooper, G.J. Kahn-J.R. & C.E.C. Freitas. 2016. Amazon floodplain fish communities: Habitat connectivity and conservation in a rapidly deteriorating environment. Biological Conservation 195: 118-127.
Ideam. 2004. Guía técnico científica para la ordenación y manejo de cuencas hidrográficas en Colombia. Instituto de Hidrología, Meteorología y Estudios Ambientales. 2004. (decreto 1729 de 2002), Bogotá, Colombia, 100 pp.
Jackson, D.A. 1993. Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology, 74: 2204-2214.
Junk, W.J. & K.M. Wantzen. 2004. The flood pulse concept: new aspects, approaches and applications - an update. In: R.L. Welcomme & T. Petr (eds). Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries (Volume II): 117-140.
Lasne, E., S. Lek & P. Laffaille. 2007. Patterns in fish assemblages in the Loire floodplain: The role of hydrological connectivity and implications for conservation. Biological Conservation 139 (2007) 258-268.
Luz-Agostinho, K.D.G., A.A. Agostinho, L.C. Gomes, & H.F. Júlio-Jr. 2008. Influence of flood pulses on diet composition and trophic relationships among piscivorous fish in the upper Paraná River floodplain. Hydrobiologia 607: 187-198.
Maldünado, J.A., A. Ortega, J.S. üsma, G. Galvis, F.A. Villa, G.L Vásqüez, S. Prada & R.C. Ardila. 2005. Peces de los Andes de Colombia. Instituto de Investigación de Recursos Biológicos «Alexander von Humboldt», Bogotá. 346 pp.
Martin, S.M., K. Lorenzen, R. I. Arthur, P. Kaisone & K. Soüvannalangsy, K. 2011. Impacts of fishing by dewatering on fish assemblages of tropical floodplain wetlands: A matter of frequency and context. Biological Conservation 144: 633-640.
Mccune, B. & M.J. Mefford. 2011. PC-ORD. Multivariate analysis of ecological data. Version 6. MjM Software, Gleneden Beach, Oregon. 28 pp.
Mojica, J.I., G. Galvis, P.C. Sánchez-Duarte, C. Castellanos & F.A. Villa-Navarro. 2006. Peces del valle medio del río Magdalena, Colombia. Biota Colombiana 7(1) 23-28.
Mojica, J.I., J.S. üsma, R. Álvarez-León & C.A. Lasso (Eds). 2012. Libro rojo de peces dulceacuícolas de Colombia. 2012. Instituto de Investigación de Recursos Biológicos «Alexander von Humboldt», Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, WWF Colombia, Universidad de Manizales. Bogotá, Colombia. 319 pp.
Montoya, Y. & N. Aguirre. 2009. Estado del arte de la limnología de lagos de planos inundables (Ciénagas) en Colombia. Gestión y Ambiente 12(3): 85-106.
Nelson, J.S. 2006. Fishes of the World. 4th ed. Jhon Wiley & Sons, Hoboken (New Jersey, USA). 601 pp.
Patrón, M. 2004. Uso inadecuado y deterioro de los sistemas cenagosos del medio y bajo Sinú y la cuenca del río San Jorge. En: J.C. Mira (ed). Resolución de conflictos ambientales. Metodología y estudios de caso en el Caribe colombiano: 55-66.
Ecofondo, Ministerio de Ambiente, Vivienda y Desarrollo Territorial, Bogotá. 111 pp.
Pava, E., J. Carrasquilla & W. López. 2006. Caracterización de la comunidad fito-planctónica de un plano de inundación del río Sogamoso en la cuenca media del río Magdalena (ciénaga de San Silvestre, Colombia). Revista Intrópica 3: 43-57.
Ramírez, A. & G.A. Pinilla. 2012. Hábitos alimentarios, morfometría y estados gonadales de cinco especies de peces en diferentes períodos climáticos en el río Sogamoso (Santander, Colombia). Acta Biológica Colombiana 17(2): 241-258.
Rincón, C. & T. Rivas. 2002. Estudio de peces promisorios en dos ciénagas de la cuenca del río Atrato, Chocó. Revista Institucional Universidad Tecnológica del Chocó: Investigación, Biodiversidad y Desarrollo 16: 42-46.
Schultz, L. 1944. A new loricariid catfish from the Rio Truando, Colombia. Copeia 1944(3): 155-156.
Tetleta-Rangel, E., J.L. Hernández-Stefanoni & J.M. Dupuy. 2013. Patterns of rare woody species richness: the influence of environment, landscape attributes and spatial structure across different spatial scales. Biodiversity and Conservation 22(6): 1435-1450.
Thomaz, S.M., L.M. Bini & R.L. Bozelli. 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579: 1-13.
Winemiller, K.O. & D.B Jepsen. 1998. Effects of seasonality and fish movement on tropical river food webs. Journal of Fish Biology 53 (Supplement A), 267-296.
Recibido: 11/12/2015 Aceptado: 01/11/2016
Referencias
Agostinho, A.A., L.C. Gomes, S. Veríssimo & E.K. Okada. 2004. Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11–19.
Agostinho, A.A., F.M. Pelicice, A.C. Petry, L.C. Gomes & H.F. Júlio-Jr. 2007. Fish diversity in the upper Paraná River basin: habitats, fisheries, management and conservation. Aquatic Ecosystem Health & Management 10(2):174–186.
Agostinho, A.A., C.C. Bonecker, & L.C. Gomes. 2009. Effects of water quantity on connectivity: the case of the upper Paraná River floodplain. Ecohydrology and Hydrobiology 9(1): 99-113.
Burgess, O.T., W.E. Pine Iii & S.J. Walsh. 2012. Importance of floodplain connectivity to fish populations in the Apalachicola river, Florida. River Research and Applications 29(6): 718–733.
Casatti, L., J.L. Veronezi Júnior & C.P. Ferreira. 2009. Diet of the armored catfish Aspidoras fuscoguttatus (Ostariophysi, Callichthyidae) in streams with different limnological and structural features. Biota Neotropica 9(1): 113-121.
Castellanos-M, Ca., N. Chamarravi, E. Castañeda, F. Galvis & E. Cepeda. 2011. Variación espacio temporal de la cobertura vegetal en la microcuenca Ciénaga El Llanito, Santander, Colombia. Boletín Científico Centro de Museos, Museo de Historia Natural 15(2): 60-68.
Claro-Jr, L., E. Ferreira, J. Zuanon & C. Araujo-Lima. 2004. Efeito da floresta alagada na alimentação de três espécies de peixes onívoros em lagos de várzea da Amazônia Central, Brasil. Acta Amazônica 34(1): 133-137.
Covain, R. & S. Fisch-Muller. 2007. The genera of the Neotropical armored catfish subfamily Loricariinae (Siluriformes: Loricariidae): a practical key and synopsis. Zootaxa 1462: 1-40.
Dahl, G. 1971. Los peces del norte de Colombia. Ministerio de Agricultura, Instituto de los Recursos Naturales Renovables INDERENA, Bogotá. 391 pp.
Díaz-Granados, M., L. Camacho & A. Maestre. 2001. Modelación de balances hídricos de ciénagas fluviales y costeras colombianas. Revista de Ingeniería. Universidad de los Andes. Facultad de Ingeniería 13: 12-20.
Fearnside, P.M. 2014. Impacts of Brazil’s Madeira River Dams: Unlearned lessons for hydroelectric development in Amazonia. Environmental Science & Policy 38: 164-172.
Galvis, G. & J.I. Mojica. 2007. The Magdalena River fresh water fishes and fisheries. Aquatic Ecosystem Health & Management 10(2): 127-139.
Gaston, K.J. 2000. Global patterns in biodiversity. Nature 405: 220-227.
Géry, J. 1977. Charachoids of the World. T. F. H. Publications, New Jersey. 294 pp.
Granado-Lorencio, C., C.R.M. Araujo-Lima & J. Lobón-Cerviá. 2005. Abundance-Distribution relationships in fish assembly of the Amazonas floodplain lakes. Ecography 28(4): 515-520.
Granado-Lorencio, C., A. Gulfo, F. Alvarez, L.F. Jiménez-Segura, J.D. Carvajal-Quintero & A. Hernández-Serna. 2012a. Fish assemblages in floodplain lakes in a Neotropical river during the wet season (Magdalena River, Colombia). Journal of Tropical Ecology 28: 271-279.
Granado-Lorencio, C., A. Hernández-Serna, J.D. Carvajal, L.F. Jiménez-Segura, A. Gulfo & F. Álvarez. 2012b. Regionally nested patterns of fish assemblages in floodplain lakes of the Magdalena River (Colombia). Ecology and Evolution 2(6): 1296-303.
Gutiérrez, F.P., C.A. Lasso, P. Sánchez-Duarte & D.L Gil. 2010. Análisis de riesgo para especies acuáticas continentales y marinas. En: Baptiste, M.P., N. Castaño, D. Cárdenas, F.P. Gutiérrez, D. Gil y C.A. Lasso (eds) Análisis de riesgo y propuesta de categorización de especies introducidas en Colombia. 73-148 pp. Instituto de Investigación de Recursos
Biológicos «Alexander von Humboldt», Bogotá, Colombia.
Guzmán, A. 2005. Aspectos históricos y técnicos del río Magdalena. Una visión sobre experiencias en el río. Hojas Universitarias 57: 35-44.
Hammer, Ø., D.A.T. Harper & P.D. Ryan. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1): 1-9.
Heino, H., A.S. Melo, T. Siqueira, J. Soininen, S. Valanko & L.M. Bini. 2015. Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshwater Biology 60(5): 845-869.
Hurd, L.E., R.G.C. Sousa, F.K. Siqueira-Souza, G.J. Cooper, G.J. Kahn-J.R. & C.E.C. Freitas. 2016. Amazon floodplain fish communities: Habitat connectivity and conservation in a rapidly deteriorating environment. Biological Conservation 195: 118-127.
Ideam. 2004. Guía técnico científica para la ordenación y manejo de cuencas hidrográficas en Colombia. Instituto de Hidrología, Meteorología y Estudios Ambientales. 2004. (decreto 1729 de 2002), Bogotá, Colombia, 100 pp.
Jackson, D.A. 1993. Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology 74: 2204-2214.
Jun k, W.J. & K.M. Want zen. 2004. The flood pulse concept: new aspects, approaches and applications - an update. In: R.L. Welcomme & T. Petr (eds). Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries (Volume II): 117-140.
Lasne, E., S. Lek & P. Laffaille. 2007. Patterns in fish assemblages in the Loire floodplain: The role of hydrological connectivity and implications for conservation. Biological Conservation 139: 258-268.
Luz-Agostinho, K.D.G., A.A. Agostinho, L.C. Gomes, & H.F. Júlio-Jr. 2008. Influence of flood pulses on diet composition and trophic relationships among piscivorous fish in the upper Paraná River floodplain. Hydrobiologia 607: 187-198.
Maldonado, J.A., A. Ortega, J.S. Usma, G. Galvis, F.A. Villa, G.L Vásquez, S. Prada & R.C. Ardila. 2005. Peces de los Andes de Colombia. Instituto de Investigación de Recursos Biológicos «Alexander von Humboldt», Bogotá. 346 pp.
Martin, S.M., K. Lorenzen, R. I. Arthur, P. Kaisone & K. Souvannalangsy, K. 2011. Impacts of fishing by dewatering on fish assemblages of tropical floodplain wetlands: A matter of frequency and context. Biological Conservation 144: 633-640.
Mccune, B. & M.J. Mefford. 2011. PC-ORD. Multivariate analysis of ecological data. Version 6. MjM Software, Gleneden Beach, Oregon. 28 pp.
Mojica, J.I., G. Galvis, P.C. Sánchez-Duarte, C. Castellanos & F.A. Villa-Navarro. 2006. Peces del valle medio del río Magdalena, Colombia. Biota Colombiana 7(1): 23-28.
Mojica, J.I., J.S. Usma, R. Álvarez-León & C.A. Lasso (Eds). 2012. Libro rojo de peces dulceacuícolas de Colombia. 2012. Instituto de Investigación de Recursos Biológicos «Alexander von Humboldt», Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, WWF Colombia, Universidad de Manizales. Bogotá, Colombia. 319 pp.
Monto ya, Y. & N. Aguirre. 2009. Estado del arte de la limnología de lagos de planos inundables (Ciénagas) en Colombia. Gestión y Ambiente 12(3): 85106.
Nelson, J.S. 2006. Fishes of the World. 4th ed. Jhon Wiley & Sons, Hoboken (New Jersey, USA). 601 pp.
Patrón, M. 2004. Uso inadecuado y deterioro de los sistemas cenagosos del medio y bajo Sinú y la cuenca del río San Jorge. En: J.C. Mira (ed). Resolución de conflictos ambientales. Metodología y estudios de caso en el Caribe colombiano: 55-66. Ecofondo, Ministerio de Ambiente, Vivienda y Desarrollo Territorial, Bogotá. 111 pp.
Pava, E., J. Carrasquilla & W. López. 2006. Caracterización de la comunidad fitoplanctónica de un plano de inundación del río Sogamoso en la cuenca media del río Magdalena (ciénaga de San Silvestre, Colombia). Revista Intrópica 3: 43-57.
Ramírez, A. & G.A. Pinilla. 2012. Hábitos alimentarios, morfometría y estados gonadales de cinco especies de peces en diferentes períodos climáticos en el río Sogamoso (Santander, Colombia). Acta Biológica Colombiana 17(2): 241-258.
Rincón, C. & T. Rivas. 2002. Estudio de peces promisorios en dos ciénagas de la cuenca del río Atrato, Chocó. Revista Institucional Universidad Tecnológica del Chocó: Investigación, Biodiversidad y Desarrollo 16: 42-46.
Schultz, L. 1944. A new loricariid catfish from the Rio Truando, Colombia. Copeia 1944(3): 155-156.
Tetleta-Rangel, E., J.L. Hernández-Stefanoni & J.M. Dupuy. 2013. Patterns of rare woody species richness: the influence of environment, landscape attributes and spatial structure across different spatial scales. Biodiversity and Conservation 22(6): 1435-1450.
Thomaz, S.M., L.M. Bini & R.L. Bozelli. 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579: 1-13.
Winemiller, K.O. & D.B Jepsen. 1998. Effects of seasonality and fish movement on tropical river food webs. Journal of Fish Biology 53 (Supplement A): 267-296.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Raniere Garcez Costa Sousa, Nathalya Sousa Oliveira, Fábio Ricardo da Rosa. (2022). The flood pulse regulates the longitudinal distribution of fish assemblages in the amazonian floodplain lakes. Boletim do Instituto de Pesca, 48 https://doi.org/10.20950/1678-2305/bip.2022.48.e688.
2. Juan Camilo Delgado Gaona. (2025). Extractivismo petrolero, degradación ambiental y resistencia comunitaria: defensa de los caños Rosario y San Silvestre en Barrancabermeja, Magdalena Medio colombiano. Revista Controversia, (225), p.1. https://doi.org/10.54118/controver.vi225.1386.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2016 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















