



Publicado
New species of Leptocaris and a new record of Darcythompsonia inopinata (Harpacticoida: Darcythompsoniidae) from Colombia
Nuevas especies de Leptocaris y un nuevo registro de Darcythompsonia inopinata (Harpacticoida: Darcythompsoniidae) de Colombia
DOI:
https://doi.org/10.15446/caldasia.v39n2.61566Palabras clave:
Caribbean, Copepoda, Crustacea, taxonomy. (en)Caribe, Copepoda, Crustacea, taxonomía (es)
Descargas
Water samples taken from Rodadero Bay (Colombia) yielded three species of harpacticoid copepods of the family Darcythompsoniidae. Leptocaris colombiana sp. nov. is attributed to the brevicornis-group, and is characterized by the five-segmented antennule, discrete (not fused) female genital somite and third urosomite, six setae on the caudal rami, two setae on the antennal exopod, lack of abexopodal seta on the allobasis of the antenna, mandibular palp reduced to one seta, one inner seta only on the first endopodal segment of leg one and second segment with two apical elements, and by three setae on leg five. Leptocaris vicina sp. nov., is attributed to the mangalis-group, and is similar to L. stromatolicola in the armature formula of leg one to leg four, female leg five with two inner small subequal setae and one outer long element, five-segmented female antennule, antenna with one abexopodal seta on allobasis, antennal exopod reduced to two setae, mandibular palp reduced to one seta, and maxilla with two endites. These two species can be separated by the shape of the anal operculum, by the relative length of the endopodal segments of legs two and three, and by the innermost distal spine of the endopodal segment of the antenna. The record of Darcythompsonia inopinata from northern Colombia represents a continuum along the Caribbean coast of the Pacific dominion, Brazilian subregion.
Se hallaron tres especies de copépodos harpacticoides de la familia Darcythompsoniidae en muestras de agua tomadas de la Bahía del Rodadero (Colombia). Leptocaris colombiana sp. nov. pertenece al grupo brevicornis, y se caracteriza por la anténula de cinco segmentos, somita genital y tercer urosomita de la hembra discretos (no fusionados), ramas caudales con seis setas, exópodo de la antena representado por dos setas, alobase de la antena sin seta abexopodal, palpo mandibular reducido a una seta, primer segmento del endópodo de la pata uno con una seta interna y segundo segmento con dos setas apicales, y pata cinco con tres setas. Leptocaris vicina sp. nov. pertenece al grupo mangalis y es similar a L. stromatolicola en la fórmula de setas y espinas de las patas uno a cuatro, pata cinco de la hembra con dos setas internas pequeñas de la misma longitud y un elemento externo largo, anténula de la hembra de cinco segmentos, alobase de la antena con una seta abexopodal, exópodo de la antena reducido a dos setas, palpo mandibular reducido a una seta, maxila con dos enditos. Estas dos especies pueden distinguirse por la forma del opérculo anal, por la longitud relativa de los segmentos del endópodo de las patas dos y tres, y por la espina interna del segmento del endópodo de la antena. Finalmente, el registro de Darcythompsonia inopinata en el norte de Colombia representa un continuo a lo largo de la costa caribeña del dominio del Pacífico, subregión Brasileña.
Recibido: 14 de marzo de 2017; Aceptado: 30 de junio de 2017
ABSTRACT
Water samples taken from Rodadero Bay (Colombia) yielded three species of harpacticoid copepods of the family Darcythompsoniidae. Leptocaris colombiana sp. nov. is attributed to the brevicornis-group, and is characterized by the five-segmented antennule, discrete (not fused) female genital somite and third urosomite, six setae on the caudal rami, two setae on the antennal exopod, lack of abexopodal seta on the allobasis of the antenna, mandibular palp reduced to one seta, one inner seta only on the first endopodal segment of leg one and second segment with two apical elements, and by three setae on leg five. Leptocaris vicina sp. nov., is attributed to the mangalis-group, and is similar to L. stromatolicola in the armature formula of leg one to leg four, female leg five with two inner small subequal setae and one outer long element, five-segmented female antennule, antenna with one abexopodal seta on allobasis, antennal exopod reduced to two setae, mandibular palp reduced to one seta, and maxilla with two endites. These two species can be separated by the shape of the anal operculum, by the relative length of the endopodal segments of legs two and three, and by the innermost distal spine of the endopodal segment of the antenna. The record of Darcythompsonia inopinata from northern Colombia represents a continuum along the Caribbean coast of the Pacific dominion, Brazilian subregion.
Key words:
Caribbean, Copepoda, Crustacea, taxonomy.RESUMEN
Se hallaron tres especies de copépodos harpacticoides de la familia Darcythompsoniidae en muestras de agua tomadas de la Bahía del Rodadero (Colombia). Leptocaris colombiana sp. nov. pertenece al grupo brevicornis, y se caracteriza por la anténula de cinco segmentos, somita genital y tercer urosomita de la hembra discretos (no fusionados), ramas caudales con seis setas, exópodo de la antena representado por dos setas, alobase de la antena sin seta abexopodal, palpo mandibular reducido a una seta, primer segmento del endópodo de la pata uno con una seta interna y segundo segmento con dos setas apicales, y pata cinco con tres setas. Leptocaris vicina sp. nov. pertenece al grupo mangalis y es similar a L. stromatolicola en la fórmula de setas y espinas de las patas uno a cuatro, pata cinco de la hembra con dos setas internas pequeñas de la misma longitud y un elemento externo largo, anténula de la hembra de cinco segmentos, alobase de la antena con una seta abexopodal, exópodo de la antena reducido a dos setas, palpo mandibular reducido a una seta, maxila con dos enditos. Estas dos especies pueden distinguirse por la forma del opérculo anal, por la longitud relativa de los segmentos del endópodo de las patas dos y tres, y por la espina interna del segmento del endópodo de la antena. Finalmente, el registro de Darcythompsonia inopinata en el norte de Colombia representa un continuo a lo largo de la costa caribeña del dominio del Pacífico, subregión Brasileña.
Palabras clave:
Caribe, Copepoda, Crustacea, taxonomía.INTRODUCTION
The marine and brackish harpacticoid fauna from Colombia has received some attention recently, and 15 families, 28 genera, and 48 species of harpacticoid copepods are known (Fuentes-Reinés and Zoppi de Roa 2013a, b, Fuentes-Reinés and Gómez 2014, Fuentes-Reinés and Suárez-Morales 2014a, b, Fuentes-Reinés et al. 2015, Gómez and Fuentes-Reinés 2017, Suárez-Morales and Fuentes-Reinés 2015a, b), including the three representatives of the family Darcythompsoniidae reported herein. Although they are common inhabitants of mangrove ecosystems (Por 1983, Gee and Somerfield 1997, Boxshall and Halsey 2004, Huys et al. 2016), members of the family Darcythompsoniidae are also common in some other habitats (Zamudio-Valdéz and Reid 1990, Fiers 1986). Several harpacticoid copepods were gathered in water samples taken from mangrove ecosystems and oyster banks in Rodadero Bay, Magdalena, northern Colombia. Among others, we found specimens of Tisbintra (Tisbidae) and Geehydrosoma (Cletodidae), whose descriptions were published earlier (Gómez and Fuentes-Reinés 2017). Darcythompsoniids were scarce and only three specimens were retrieved. Two of them attributed to the genus Leptocaris and whose complete descriptions are given below, turned out to be new to science. Additionally, we report on the presence of Darcythompsonia inopinata Smirnov, 1934 in Rodadero Bay, Magdalena, northern Colombia.
MATERIALS AND METHODS
Eleven water samples were collected from mangrove ecosystems and at an oyster bank in Rodadero Bay, Magdalena, northern Colombia (11°14' North, 74°12' West). These samples were collected monthly from August 2015 to June 2016. Samples of 432 l each were taken using a 25 l bucket, sieved with a 45 μm zooplankton net, and preserved in 70% ethanol. Water salinity, pH, and temperature were measured with a Multi 350i handheld meter. Copepods were sorted manually and processed for taxonomic identification. Observations and drawings of the material presented herein were made from whole and dissected material. Dissected parts were mounted in lactophenol under a Leica compound microscope equipped with phase contrast and a drawing tube at a magnification of 1000X. Partially dissected material - habitus, right antennule, labrum, and P5 of Leptocaris colombiana sp. nov.- was left intact and preserved in alcohol.
The material examined was deposited in the Copepoda collection of the Instituto de Ciencias del Mar y Limnología, Mazatlán Marine Station (ICML-EMUCOP) and in the Centro de Colecciones Biológicas de la Universidad del Magdalena-Colombia (CBUMAG). Morphological terminology follows 0 except for the maxilla. Morphological terminology for the latter follows Ferrari and Ivanenko (2008). The following abbreviations are used throughout the text and figures: ae, aesthetasc; P1-P4 ENP/EXP1-3, first (proximal) to third (distal) endopodal/exopodal segment of first to fourth leg; P5, fifth leg.
Note also that the genus name Leptocaris is feminine (Art. 30.1.2; ICZN 1999) and Apostolov's (2007) and Huys' (2009) views have been adopted here for the masculine names given to some species of the genus. The biogeographical regionalization of the Neotropical region by Morrone (2014) was adopted.
RESULTS
Taxonomy
Order Harpacticoida Sars, 1903 Family Darcythompsoniidae Lang, 1936 Leptocaris Scott T., 1899 Leptocaris colombiana sp. nov. (Figs. 1-3)

Figure 1:
Leptocaris colombiana
sp. nov. Female. a. cephalothorax, dorsal, posterior long sensilla indicated by asterisks, b. free prosomites, dorsal, c. urosome, dorsal, d. urosome, ventral, e. posterior half of anal somite and caudal rami, dorsal, f. genital field and P5. Scale a,c,d,e = 100 μm, Scale b,f = 50 μm.
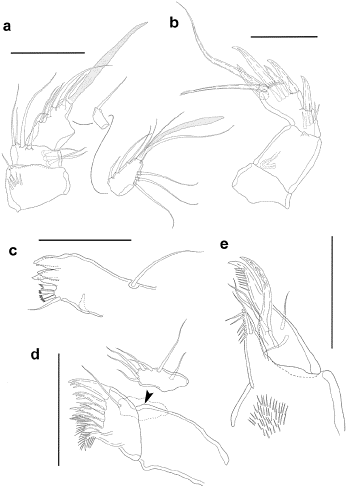
Figure 2:
Leptocaris colombiana
sp. nov. Female. a. antennule, b. antenna, c. mandible, d. maxillule, e. maxilla. Scale = 25 μm.
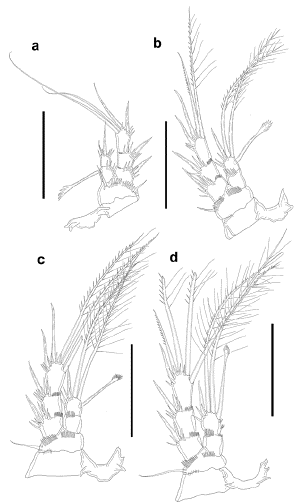
Figure 3:
Leptocaris colombiana
sp. nov. Female. a. P1, anterior, b. P2, anterior, c. P3, anterior, d. P4, anterior. Scale = 50 μm.
Type material. Adult female holotype partially dissected as follows: habitus, right antennule, labrum, and P5 left intact and preserved in alcohol (ICML-EMUCOP-010815-11); left antennule, antennae, mandibles, maxillulae, maxillae, P1-P4 dissected (ICML-EMUCOP-010815-12) and mounted onto four slides; Colombia, Magdalena, Rodadero Bay, 11°14' North, 74°12' West; August 2015; coll. J. M. Fuentes-Reinés.
Diagnosis
Habitus elongate, cylindrical. Rostrum minute. Female antennule five-segmented. Antenna with allobasis, without abexopodal seta; exopod represented by two setae; endopod one-segmented, with seven elements, of which apical inner spine smallest. Mandible with well-developed coxa and gnathobase; palpus represented by single seta. Maxillule with well-developed arthrite bearing four distal multicuspidate spines and three pinnate setae, and one surface seta; coxal endite, basis, exopod and endopod fused, with seven setae in all. Syncoxa of maxilla with one (probably coxal) endite armed with one stout spinulose element fused to endite basally, and one slender seta; allobasis with one (probably basal) strong claw with one accompanying seta, drawn out into strong curved claw with one accompanying seta (probably endopodal), endopod represented by two setae. Maxilliped absent. P1-P4 with three-segmented exopod and two-segmented endopod. Female P5 reduced, fused to somite ventrally, with three setae in all. Caudal rami with six setae.
Description of female
Habitus elongate, cylindrical, without a clear division between pro- and urosome. Total body length measured from anterior margin of rostrum to posterior margin of caudal rami, 620 μm. Cephalothorax (Fig. 1a) rounded anteriorly; with minute surface denticles posteriorly; with sensilla as shown; medial and posterior sensilla very long (indicated in Fig. 1a). Rostrum (Fig. 1a) minute, fused to cephalothorax, with two long sensilla. Surface of free prosomites (Fig. 1b) covered with small denticles; posterior margin of somites with coarser denticles and with some short sensilla; with two dorsal and two lateral long sensilla on posterior third of somites. First urosomite (P5-bearing somite; Fig. 1c) ornamented with denticles dorsally as previous prosomites; ventrally without denticles, with some sensilla along posterior margin only, with two small medial pores close to P5 (Fig. 1d). Genital somite and third urosomite discrete (Figs. 1c, d); genital somite with small denticles dorsally, posterior denticles not coarser than the rest (Fig. 1c); ventrally without denticles, with some sensilla along posterior margin only (Fig. 1d); genital field (Figs. 1d, f) located on anterior half of somite, with medial copulatory pore. Third urosomite with denticles and sensilla as in P5-bearing somite dorsally (Fig. 1c); ventrally with one row of slender spinules on proximal third, with one medial short row of slender spinules close to posterior margin, and one long row of slender spinules along posterior margin medially (Fig. 1d). Fourth urosomite as previous somite dorsally (Fig. 1c); ventrally without denticles, with long medial row of slender spinules along posterior margin (Fig. 1d). Fifth urosomite with small denticles dorsally, those along posterior margin not visibly coarser than the rest (Fig. 1c); without ventral ornamentation (Fig. 1d). Anal somite (Figs. 1c-e) elongate, conical, tapering distally, about as long as two previous somites combined; with crescentic anal operculum not reaching posterior margin of somite (Fig. 1e); the two sensilla associated to the anal operculum separated by a gap visibly wider than the width of the anal operculum, and displaced medially (Figs. 1c, e). Caudal rami (Figs. 1c-e) ovate, about twice as long as wide; with six setae as follows (seta I lost): seta II and III vestigial, situated laterally on distal half of ramus; seta IV well-developed, displaced dorsally close to vestigial setae II and III; apical seta V longest; inner seta VI arising distally; dorsal seta VII arising medially, articulated basally.
Antennule (Fig. 2a) short, five-segmented. Armature formula as follows: 1(1)-2(7)-3 (5+[1+ae])-4(1)-5(8+[1+ae]); all setae smooth; three setae articulated on last segment.
Antenna (Fig. 2b) with unornamented coxa. Allobasis and free endopodal segment subequal in length; the former without armature nor ornamentation. Exopod represented by two setae. Endopodal segment with one proximal and one subdistal transverse row of strong spinules, with two stout lateral spines on inner margin, and three spines and two setae distally, one of them with comb-like tip.
Mandible (Fig. 2c) with well-developed gnathobase bearing several multicuspidate teeth and one lateral seta. Palp represented by one long seta.
Maxillule (Fig. 2d) with unornamented praecoxa. Arthrite well-developed, with four multicuspidate spines and three pinnate setae; with one surface seta; coxal endite, basis, exopod and endopod fused, with two proximal, two subdistal and three distal setae.
Maxilla (Fig. 2e) with syncoxa ornamented with spinular patch as depicted, and with one syncoxal (probably coxal) endite with one stout spinulose element fused to endite basally, and one slender seta. Allobasis with one (probably basal) strong claw with one accompanying seta, drawn out into strong curved claw with one accompanying seta (probably endopodal); endopod represented by two setae.
Maxilliped absent.
P1 (Fig. 3a). Coxa seemingly without ornamentation. Basis with one subdistal spinular row medially, and one transverse spinular row close to exopod, with one inner spine-like element and one outer seta. Exopod three-segmented; all segments with outer spinules close to outer spine and distally; EXP1 and EXP2 without inner seta; EXP3 with one outer spine, two apical and one inner bare elements; all elements seemingly bare. Endopod two-segmented; reaching distal margin of EXP2; ENP1 with some subdistal spinules, with one inner seta oriented downwards and with comb-like tip; ENP2 with some subdistal spinules, with one apical outer bare spine and one apical inner bare short seta.
P2 (Fig. 3b). Coxa with some small spinules close to outer corner. Basis with one subdistal spinular row medially, and one transverse spinular row close to exopod; outer seta short, spine-like. Exopod three-segmented; all segments with outer spinules close to outer bare spine and distally; EXP1 and EXP2 without inner armature; EXP3 with two outer bare spines and two apical setae -outermost with outer spinules, innermost with outer spinules and inner setules-. Endopod two-segmented, barely reaching tip of EXP2, with spinular ornamentation as shown; ENP1 without armature; ENP2 with one strong inner seta with comb-like tip, two apical setae with outer spinules medially and with setules distally and along inner margin, and one outer apical bare spine.
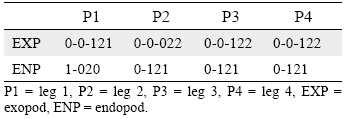
P3-P4 (Figs. 3c, d). Coxa with some small spinules close to outer corner. Basis with one subdistal spinular row medially, and one transverse spinular row close to exopod; outer element setiform, longer than in P2. Exopod three-segmented; all segments with outer spinules close to outer bare spine and distally; EXP1 and EXP2 without inner armature; EXP3 with two outer bare spines, two apical setae with outer spinules and inner setules, and one inner plumose element. Endopod two-segmented, barely reaching tip of EXP2, with spinular ornamentation as shown; ENP1 without armature; ENP2 with one strong inner seta with comb-like tip, two apical setae with outer spinules medially and with setules distally and along inner margin, and one outer apical bare spine. Armature formula of P1-P4 as in table 1.
Table 1. Armature formula of P1-P4 of Leptocaris colombiana sp. nov.
P5 (Figs. 1d, f) represented by small segment fused to somite ventrally; each leg with three bare setae, of which innermost and outermost subequal in length, medial shorter.
Etymology
The specific epithet, colombiana, refers to the country where the species was found.
Leptocaris vicina sp. nov.
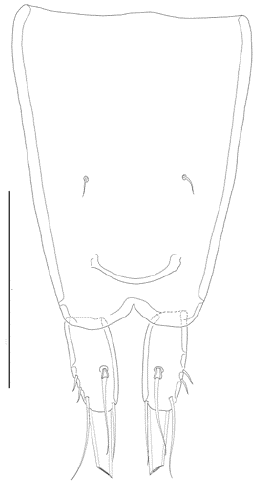
Figure 4:
Leptocaris vicina
sp. nov. Female. Anal somite and caudal rami, dorsal. Scale = 50 μm.

Figure 5:
Leptocaris vicina
sp. nov. Female. a. antennule, b. antenna, small inner apical spine indicated with an arrow, c. maxillule, d. maxilla. Scale = 25 μm.
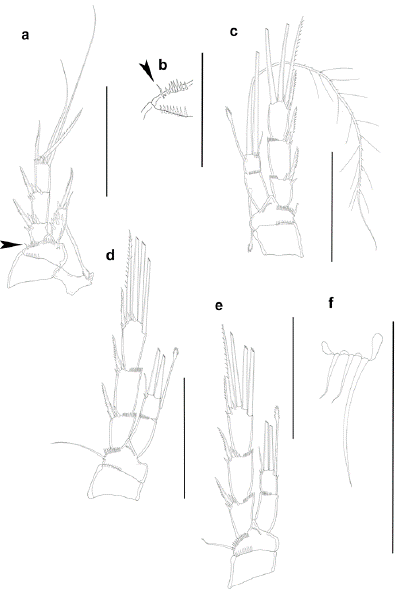
Figure 6:
Leptocaris vicina
sp. nov. Female. a. P1, anterior, outer basal seta indicated with an arrow, b. outer corner of basis of P1, outer basal seta indicated with an arrow, anterior, c. P2, anterior, d. P3, anterior, e. P4, anterior, f. P5. Scale a,c,d,e =50 μm, b,f =25 μm.
Type material. Adult female holotype partially dissected as follows: urosome left intact and preserved in alcohol (ICML-EMU-COP-010815-14); antennules, antennae, mouth parts, and P1-P4 dissected and mounted onto five slides (ICML-EMU-COP-010815-13); Colombia, Magdalena, Rodadero Bay, 11°14' North, 74°12' West, August 2015; coll. J. M. Fuentes-Reinés.
Diagnosis
Habitus elongate, cylindrical. Rostrum minute. Female antennule five-segmented. Antenna with allobasis, with abexopodal seta; exopod represented by two setae; endopod one-segmented, with seven elements, of which apical inner spine very small. Mandible with well-developed coxa and gnathobase; palpus represented by single seta. Maxillule with well-developed arthrite bearing four distal spines and two pinnate elements, with one surface seta; coxal endite, basis, exopod and endopod fused, with four setae. Syncoxa of maxilla with one (probably coxal) endite with one stout spinulose element fused to endite basally, and two slender setae; allobasis with one (probably basal) strong claw with two accompanying setae, drawn out into strong curved pinnate claw with one accompanying seta (probably endopodal), endopod represented by two setae. Maxilliped absent. P1-P4 with three-segmented exopod; P1 with one-segmented endopod; P2-P4 with two-segmented endopod. Female P5 reduced, fused to somite ventrally, with three setae in all, of which outermost longest. Caudal rami with six setae.
Description of female
Habitus as in L. stromatolicola Zamudio-Valdéz & Reid, 1990, elongate, cylindrical, without a clear division between pro- and urosome. Total body length measured from anterior margin of rostrum to posterior margin of caudal rami, about 533 μm. Dorsal surface of pro- and urosomites smooth, without ornamentation. Rostrum minute, fused to cephalothorax, with two sensilla. Genital somite and third urosomite fused, without trace of division, forming genital double-somite; genital field located on anterior half of somite, with medial copulatory pore. Posterior half of genital double-somite and following somite with medial row of minute spinules close to posterior margin. Fifth urosomite without ornamentation. Anal somite (Fig. 4) about 1.3 times as long as preceding somite, tapering posteriorly; with naked rounded anal operculum; the two sensilla associated to the anal operculum separated by a gap about as wide as the width of the anal operculum, and displaced medially. Caudal rami (Fig. 4) ovate, about twice as long as wide; with six setae as follows (seta I lost): seta II and III vestigial, situated laterally on distal half of ramus; seta IV well-developed, displaced dorsally close to vestigial setae II and III; apical seta V longest; inner seta VI arising distally; dorsal seta VII arising medially, articulated basally.
Antennule (Fig. 5a) short, five-segmented. Armature formula as follows: 1(1)-2(7)-3(4+[1+ae])-4(1)-5(8+[1+ae]); all setae smooth; three setae on last segment articulated.
Antenna (Fig. 5b) with allobasis, the latter with abexopodal seta with comb-like tip, and about as long as free endopodal segment. Exopod represented by two setae. Endopodal segment with one proximal and one subdistal transverse row of strong spinules, with two stout lateral spines on inner margin, distally with three spines (innermost small and can be mistaken for spinule, arrowed in Fig. 5b), and two setae, one of them with comb-like tip.
Mandible as in previous species.
Maxillule (Fig. 5c) similar to that of L. stromatolicola, with four distal spines and two pinnate setae on well-developed arthrite, and one surface seta. Coxal endite, basis, exopod and endopod fused, with two lateral and two distal setae.
Maxilla (Fig. 5d) with one syncoxal (probably coxal) endite with one stout spinulose element fused to endite basally, and seemingly two slender setae. Allobasis with one (probably basal) strong claw with two accompanying setae, drawn out into strong curved pinnate claw with one accompanying seta (probably endopodal), endopod represented by two setae.
Maxilliped absent.
P1 (Fig. 6a, b). Coxa with posterior row of small spinules close to basis. Basis with one subdistal spinular row medially, and one transverse spinular row close to exopod, with one inner spine-like element and one minute outer seta (the latter arrowed in Fig. 6a, b). Exopod three-segmented; all segments ornamented with subdistal and distal outer spinules; EXP1 and EXP2 without inner seta; EXP3 with two outer pinnate spines and two bare distal setae. Endopod one-segmented; reaching middle of EXP2, with stout spinules medially and subdistally, with one inner seta oriented downwards and with comb-like tip, distally with one inner small seta and one outer apical pinnate spine, the latter twice as long as the former.
P2 (Fig. 6c). Coxa with some posterior spinules close to outer corner. Basis with transverse row of small spinules close to exopod, and with minute spinules close to endopod, outer seta small. Exopod three-segmented; all exopodal segments with few distal and subdistal outer spinules; EXP1 and EXP2 without inner armature; EXP3 with two outer pinnate spines and two long apical setae with outer spinules and inner setules. Endopod two-segmented, reaching middle of EXP2; ENP1 about 1.8 times as long as ENP2, reaching beyond EXP1, with spinular ornamentation as shown and with one inner seta oriented downwards and with comb-like tip; ENP2 with some outer subapical spinules, distally with one outer small seta, two long setae with outer spinules and inner setules, and one inner seta with with comb-like tip.
P3-P4 (Figs. 6d, e). Coxa with some posterior spinules close to outer corner. Basis with transverse row of small spinules close to exopod, and with minute spinules close to endopod, outer seta well-developed, long. Exopod three-segmented; all exopodal segments with outer spinules as depicted; EXP1 and EXP2 without inner armature; EXP3 with two outer pinnate spines, two distal setae with outer spinules and inner setules, and one inner plumose element. Endopod two-segmented; ENP1 reaching tip of EXP1, about twice as long as ENP2, with some minute spinules apically, without armature; ENP2 reaching middle of EXP2, with one slender outer spine, two apical long setae and one inner element with comb-like tip. Armature formula of P1-P4 as in table 2.
Table 2. Armature formula of P1-P4 ofLeptocaris vicina sp. nov.

P5 (Fig. 6f) represented by small segment fused to somite ventrally; each leg with three bare setae, two innermost setae subequal in length, outermost longest.
Etymology
The specific epithet, vicina, from Latin vicinus, -α, -um, neighbor, makes reference to the fact that these two species were found in the same sample.
DISCUSSION
The marine and brackish harpacticoid fauna from Colombia is poorly known and have received some attention recently (e.g., Fuentes-Reinés and Zoppi de Roa 2013a, b, Fuentes-Reinés and Gómez 2014, Fuentes-Reinés and Suárez-Morales 2014a, b, Fuentes-Reinés et al. 2015, Gómez and Fuentes-Reinés 2017, Suárez-Morales and Fuentes-Reinés 2015a, b) The family Darcythompsoniidae, currently composed of the genera Darcythompsonia, Kristensenia, Leptocaris, and Pabellonia, can be found typically associated with decaying plant material in mangal ecosystems and coastal lagoons (Por 1983, Gee and Somerfield 1997, Boxshall and Halsey 2004, Huys et al. 2016), but Zamudio-Valdéz and Reid (1990) and Fiers (1986) noted that some genera of this family, e.g. Leptocaris and Darcythompsonia, are common inhabitants of places completely different to mangrove systems.
The genus Leptocaris contains 29 species (L. trisetosus (Kunz, 1935) and L. echinata Fiers, 1986 with three and two subspecies, respectively), of which 12 species (L. trisetosa and L. echinata with one subspecies each; L. trisetosa trisetosa (Kunz, 1935), L. echinata echinata Fiers, 1986, L. armata Lang, 1965, L. brevicornis (Douwe, 1904), L. doughertyi Lang, 1965, L. kunzi Fleeger & Clark, 1980, L. pori Lang, 1965, L. stromatolicola, L. glaber Fiers, 1986, L. mangalis Por, 1983, L. minima (Jakobi, 1954), and L. vermiculata (Oliveira, 1957)) have been described and/or reported from the New World. Of these, only seven species (L. echinata and L. trisetosa, with one subspecies each; L. echinata echinata, L. trisetosa trisetosa, L. brevicornis, L. glaber, L. mangalis, L. minima, and L. vermiculata) are known from the Neotropical Region. Interestingly, Huys et al. (2016) reported representatives of the brevicornis-group of the genus Leptocaris in early Miocene amber inclusions from northern Chiapas State, Southern Mexico.
Lee and Chang (2008), Song et al. (2012), and Köroğlu et al. (2014) presented a complete historical account of the family Darcythompsoniidae and its constituent genera, including the genus Leptocaris, as well as a brief revision of Kunz's (1978, 1983, 1994) species-groups, and Apostolov's (2007) reduction of species-groups from four as in Kunz (1994) to three. Fiers (1986; but see also Kunz 1994: 51) questioned the adequacy of the subdivision of the genus given the wide combination of characteristics in, for example, L. echinata echinata, and the wide variability in the general structure and armature complements of the antennal exopod, mandibular palp, presence/absence of an abexopodal seta in the antenna, fused/discrete condition of the female genital somite and third urosomite, and general structure of the maxilla, to name a few, within each species-group. Kunz's (1978, 1983, 1994) and Apostolov's (2007) subdivision of the genus are followed here for identification purposes only. However, as for many other species within the genus, the males of L. colombiana sp. nov. and L. vicina sp. nov. remain unknown, and the comparisons below are based solely on the females. The lack of males (see Lee and Chang 2008) and the wide variability observed in the number and shape of the setae on the female P5, among others, prevent any phylogenetical analysis (Fleeger and Clark 1980, Fiers 1986, Apostolov 2007).
Of particular interest here is the brevicornis-group, the most primitive species-group of the genus (Kunz 1983, Fiers 1986), defined by the presence of five setae/spines on P3-P4 EXP3, and four elements on P2-P3 ENP2 (Kunz 1983, 1994, Apostolov 2007), into which L. colombiana sp. nov., has been placed. Of interest is also the mangalis-group defined by the one-segmented endopod of P1 (Kunz 1994), into which L. vicina sp. nov. has been placed. The brevicornis-group is at present composed of L. brevicornis, L. trisetosa trisetosa, L. gurneyi (Nicholls, 1944), L. sibirica Borutsky, 1952, L. vermiculata, L. echinata echinata, L. mucronata Fiers, 1986, and L. itoi Kunz, 1994 (Song et al. 2012), but also of L. trisetosa breviseta Kunz, 1994, L. trisetosa pacifica Lee & Chang, 2008, L. echinata nuda Kunz, 1994, and L. colombiana sp. nov. Of these, only five species/subspecies are known from the New World and from the Neotropical Region, L. colombiana sp. nov., L. trisetosa trisetosa, L. echinata echinata, L. brevicornis, and L. vermiculata. The mangalis-group is composed by four species only, L. mangalis, L. stromatolicola, L. noodti Kunz, 1994, and L. vicina sp. nov. For a brief discussion on the mangalis-group see below.
As in the other species-groups of Leptocaris, an ample variety of combinations of characteristics is evident in the brevicornis-group, and any phylogenetic inference is at most speculative. This, as implicitly suggested by Fiers (1986), points to the inadequacy of the subdivision of the genus.
The brevicornis-group, as defined by Kunz (1983, 1994) and Apostolov (2007), is characterized by the presence of five elements on the third exopodal segment of P3 and P4, and four elements on the second endopodal segment of P2 and P3. However, more relevant differences are evident regarding, for example, the fused/discrete condition of the female genital field and third urosomite, number of setae on the caudal rami (less than six setae in L. brevicornis, L. trisetosa trisetosa, L. trisetosa breviseta, L. gurneyi and L. sibirica; six setae in L. vermiculata, L. echinata echinata, L. echinata nuda, L. mucronata, L. itoi and L. colombiana sp. nov.; seven setae in L. trisetosa pacifica), number of segments of the female antennule (four segments in L. mucronata; five segments in L. brevicornis, L. trisetosa trisetosa, L. trisetosa breviseta, L. trisetosa pacifica, L. gurneyi, L. vermiculata, L. echinata echinata, L. echinata nuda, and L. colombiana sp. nov.; five or six segments in L. sibirica; seven segments in L. itoi), number of setae of the antennal exopod (without setae in L. gurneyi, L. echinata echinata and L. echinata nuda; with one seta in L. sibirica; with two setae in L. brevicornis, L. trisetosa trisetosa, L. trisetosa breviseta, L. trisetosa pacifica, L. vermiculata, L. mucronata, and L. colombiana sp. nov.), presence of the abexopodal seta of the antenna in L. trisetosa pacifica, L. sibirica, L. echinata echinata, L. echinata nuda, and L. mucronata, and lack of such seta in L. trisetosa trisetosa, L. gurneyi, and L. colombiana sp. nov. (the presence/absence of such seta needs to be verified for L. brevicornis, L. trisetosa breviseta, L. vermiculata, and L. itoi), number of setae representing the mandibular palp (without seta in L. brevicornis, L. gurneyi, and L. sibirica; with one seta in L. vermiculata, L. echinata echinata, and L. colombiana sp. nov.; with two setae in L. trisetosa trisetosa, L. trisetosa pacifica, L. trisetosa breviseta, L. mucronata, and L. itoi), and armature formula of the endopod of P1 (1,111 in L. brevicornis, L. trisetosa trisetosa, L. breviseta, L. trisetosa pacifica, and L. gurneyi, L. echinata echinata, L mucronata and L. itoi; 0,020 in L. sibirica; 0,111 in L. vermiculata; 1,020 in L. echinata nuda and L. colombiana sp. nov.), P2 (0,121 in L. brevicornis, L. gurneyi, L sibirica, L. vermiculata, L. echinata echinata, L. echinata nuda, L. itoi, and L. colombiana sp. nov.; 1,121 in L. trisetosa trisetosa, L. trisetosa breviseta, L. trisetosa pacifica, and L. mucronata), P4 (0,121 in L. brevicornis, L. trisetosa trisetosa, L. trisetosa breviseta, L. trisetosa pacifica, L. gurneyi, L. sibirica, L. mucronata, L. itoi, and L. colombiana sp. nov.; 1,121 in L. vermiculata, L. echinata echinata, and L. echinata nuda), and P5 (without P5 in L. gurneyi; with two setae in L. brevicornis, L. trisetosa trisetosa, L. breviseta, and L. sibirica; three setae in L. trisetosa pacifica, L. echinata echinata, L. echinata nuda, L. mucronata, and L. colombiana sp. nov.; with four setae in L. vermiculata and L. itoi). These differences between species are not unique to the brevicornis-group, but are also observable in the other species-groups of the genus. These differences and the types of sexual dimorphism observed by Lee and Chang (2008) e.g., presence of normal setae or spiniform projections on P2ENP and P3ENP, might prove more reliable and could shed some light on the phylogenetic relationships of the species of Leptocaris.
Leptocaris colombiana sp. nov.
The fused condition of the genital somite and the third urosomite to form the double genital-somite is present in L. brevicornis, in the three subspecies of L. trisetosa, and in L. vermiculata and L. mucronata, but these somites are clearly discrete in L. sibirica, in both subspecies of L. echinata, and in L. colombiana sp. nov. The caudal rami possess less than six setae in L. brevicornis, L. trisetosa trisetosa and L. trisetosa breviseta, L. gurneyi, and L. sibirica; six setae have been observed for L. vermiculata, both subspecies of L. echinata, L. mucronata, L. itoi, and L. colombiana sp. nov.; L. trisetosa pacifica is unique by having a full complement of seven setae. The number of segments of the female antennule varies from four (only observed in L. mucronata) to seven (this condition was observed only for L. itoi), being the five-segmented the most common condition (though the five-or six-segmented condition of L. sibirica remains inconclusive; Borutzky 1952). The condition of the antennal exopod also varies among the species of the brevicornis-group. Two setae representing the antennal exopod have been observed in L. brevicornis, in the three subspecies of L. trisetosa, in L. vermiculata, L. mucronata, and L. colombiana sp. nov.; one seta has been observed for L. sibirica, while L. gurneyi, and both subspecies of L. echinata lack the antennal exopod. The antenna of L. itoi was omitted in Kunz (1994). The abexopodal seta on the allobasis of the antenna is present in L. vermiculata, L. trisetosa pacifica, L. sibirica, both subspecies of L. echinata, and in L. mucronata. This seta is absent in L. colombiana sp. nov., L. gurneyi, and in L. trisetosa trisetosa and L. trisetosa breviseta, and its presence/absence needs to be verified for L. brevicornis and L. itoi. Gurney (1920) redescribed L. brevicornis from several situations in the Norfolk Broads without abexopodal seta on the allobasis of the antenna, but noted also that one specimen bore a "blunt-pointed seta" on the allobasis of both antennae; Lee and Chang (2008) described their L. brevicornis from South Korea without abexopodal seta on the allobasis of the antenna; the antenna of L. itoi was not described in Kunz (1994). L. brevicornis, L. sibirica, and L. gurneyi lack the mandibular palp, but it is represented by one seta in both subspecies of L. echinata and in L. colombiana sp. nov., and by two setae in the three subspecies of L. trisetosa, in L. mucronata and L. itoi. Even though the brevicornis-group was defined based on the number of setae of the P3-P4 EXP3, and P2-P3 ENP2, the armature formula of the female P1-P2 ENP, P4 ENP, and P5 is somewhat variable, being the most common formulae 1,111/0,121/0,121/3 for P1 ENP, P2 ENP, P4 ENP, and P5, respectively. L. echinata nuda and L. colombiana sp. nov. possess one inner seta on P1 ENP1, but only two setae on ENP2; L. sibirica also possesses two setae only on P1 ENP2 but lacks the inner seta on ENP1; L. vermiculata possesses three setae on P1 ENP2 but lacks the inner element on ENP1. All the species/subspecies of the brevicornis-group possess four setae on the P2 ENP2, but only the three subspecies of L. trisetosa, and L mucronata possess one additional inner seta on ENP1. Similarly, all species/subspecies of this group bear four setae on the P4 ENP2, most species lack the inner seta on ENP1, and only L. vermiculata, and both subspecies of L. echinata bear an additional inner seta on ENP1. The number of elements on the female P5 is also variable within the brevicornis-group. The most drastic reduction was observed for L. gurneyi (without P5), while it is armed with two setae in L. brevicornis, L. trisetosa trisetosa and L. trisetosa breviseta, and L. sibirica. The female P5 of L. trisetosa pacifica bears three setae, casting some doubts about its relationship with the other two subspecies of L. trisetosa; three setae have been observed for both subspecies of L. echinata, for L. mucronata and L. colombiana sp. nov. Only L. vermiculata and L. itoi bear four setae on the female P5.
Leptocaris vicina sp. nov.
Kunz (1994) noted that Por's (1983) figures 48-51 and 57-61 showing the legs of L. mangalis do not correspond to Por's (1983: 148-149) table of the armature formula of P1-P4, and suggested to consider Por's (1983) table as wrong, and Por's (1983) figures as correct regarding the armature of swimming legs. Based on the rare condition of the one-segmented endopod of P1, Kunz (1994) suggested to stablish the mangalis-group for L. stromatolicola, L. noodti and L. mangalis, which is probably the only species group of the genus Leptocaris that reflects some phylogenetic relationships. Por (1983: 152) commented that Kunz (1983) would consider L. mangalis as a member of the ignavus-group, which was not the case. The mangalis-group was not considered by Apostolov (2007). Kunz (1994: 48, Tab. 1) presented a brief table with the main differences between L. stromatolicola, L. noodti, and L. mangalis. Note that Kunz (1994) described and illustrated a seven-segmented female antennule for L. noodti, but showed a four-segmented female antennule in his table 1 (Kunz 1994: 48). The one-segmented condition of the P1 ENP is very rare among the species of Leptocaris. To the best of our knowledge, this has been observed for four species only, L. stromatolicola from stromatolites in two evaporitic lakes and a marsh in the Cuatro Ciénegas basin, in central Coahuila, northern México (Zamudio-Valdéz and Reid 1990), L. noodti Kunz, 1994 from Hawaii (Kunz 1994), L. mangalis from mangrove forests of Southern Sinai (Por 1983), and L. vicina sp. nov. from Colombia; the latter was unequivocally attributed to Kunz's (1994) mangalis-group.
The mouthparts of the different species of Leptocaris have not received enough attention, probably due to their difficult dissection, and small sizes making difficult to observe their general structure and armature complements. Due to the simple structure of the mandible, and to the size of the antenna, these two appendages, along with the antennule, are by far the easiest appendages to observe and illustrate. Unfortunately, this is not the case for the maxillule and maxilla. The figures and text describing the maxilla and maxillule of L. noodti (see Kunz 1994: 47, Fig. 9E, F) and L. mangalis (see Por 1983: 148, Figs. 46-47) lack the necessary details. Nevertheless, L. noodti and L. mangalis seem to share the maxillule with coxa and basis separated. The structure of the maxilla of L. noodti and L. mangalis is misleading. The syncoxal endite of the maxilla of L. noodti appears as part of the allobasis, and the separation between the syncoxa and the allobasis is not evident in L. mangalis. These two species also share the lack of inner armature on the female P2 ENP1, and the presence of two setae only on the female P5, and can be easily separated by the number of segments of the female antennule (seven in L. noodti, but four in L. mangalis), by the presence, in L. noodti, of an abexopodal seta on the allobasis of the antenna (this seta is missing in L. mangalis), by the female armature formula of P1 EXP3 (with three setae in L. mangalis, but four elements in L. noodti), and P3-P4 EXP3 (with four setae/spines in L. mangalis, but five elements in L. noodti). Of interest, are the strong similarities between the general structure and armature complement of the mandible, maxillule and maxilla, and of P1-P5 of L. stromatolicola and L. vicina sp. nov. These two species share the combination of the following characters: armature formula of the female P1-P4 EXP/ENP (P1, 0-0121/120; P2, 0-0-121/1-121; P3, 0-0-122/0121; P4, 0-0-122/0-121), armature formula and relative length of the setae of the female P5 (with three setae; innermost elements subequal in length, half as long as outer seta), the five-segmented female antennule, and the general shape and armature of the antenna (allobasis with one abexopodal seta, exopod represented by two setae, endopodal segment with five spines and two setae), and mandible (palp represented by one seta). Furthermore, some striking similarities were observed between the general shape of the maxillule (arthrite with one surface seta, and with four spines and two pinnate setae distally), and the maxilla of L. stromatolicola and L. vicina sp. nov. Regarding the maxilla, besides the two setae representing the endopod, both species share the presence of two endites; the proximal endite of L. stromatolicola possesses one strong spinulose element only, while that of L. vicina sp. nov. possesses one strong spinulose element and two slender setae that are difficult to see and probably have been overlooked in L. stromatolicola; the second endite is armed with two slender setae and a curved strong element in both species. These two species can be separated by the shape of the anal operculum (crescentic in L. stromatolicola, but rounded in L. vicina sp. nov.), by the relative length of the segments of P2 ENP and P3 ENP (ENP1 visibly longer that ENP2 in L. vicina sp. nov., but nearly subequal in L. stromatolicola), and by the innermost distal spine of the endopodal segment of the antenna (reduced in L. vicina sp. nov., but well-developed in L. stromatolicola).
Darcythompsonia Scott T., 1906 Darcythompsonia inopinata Smirnov, 1934 Syn: for a complete list of synonyms see Fiers (1986)
Material examined. One dissected adult female (CBUMAG:MEI:0004); Colombia, Magdalena, Rodadero Bay, 11°14' N, 74°12' W, August, 2015; coll. J. M. Fuentes-Reinés.
REMARKS
Currently, there are four valid species within Darcythompsonia (D. inopinata, D. neglecta Redeke, 1953, D. fairliensis, and D. scotti Gurney, 1920). D. inopinata is the most widespread (Fiers 1986, Gómez 2000) and has been reported previously from Fiji Islands, Western Samoa, Papua New Guinea, Grand Comores Archipelago, and Solomon Islands (Fiers 1986 and references cited therein). This species is known also from the Neotropical Region and has been reported from Inagua (Bahamas), Aruba (Netherlands Antilles) (Fiers 1986) in the Antillean subregion, in Morocoy Peninsula (Venezuela) (Fiers 1986) and in the Cananéia Lagoon estuarine system (Brazil), Brazilian subregion (Por et al. 1984, Reid 1998). This is a benthic species occurring in shallow marine and brackish coastal waters (Reid 1998), but also has been reported from a variety of situations like wells, caves, shallow pools, cenotes, crabholes, and in coarse sand and shell debris (Fiers 1986). The present record of D. inopinata from northern Colombia represents a continuum along the Caribbean coast of the Pacific dominion (Brazilian subregion). The specimen from Colombia fits the diagnostic features of D. inopinata as described by Smirnov (1934) and resembles D. fairliensis in the armature formula of P1-P4, number of segments of antennule, and number of segments and setae on the antennal exopod. Nevertheless, they can be separated by the number of setae in the female P5 (three setae in D. inopinata, but four elements in D. fairliensis), and by the shape of the caudal rami (with angular extension in the proximal half in D. fairliensis, but without angular extension in D. inopinata).
AUTHORS PARTICIPATION
JFR collected the samples; SG identified the specimens; JFR and SG wrote the text. SG made the drawings
LITERATURE CITED
Referencias
Apostolov A. 2007. Harpacticoïdes marins (Copepoda, Harpacticoida) d’Islandie, 1. Le genre Halectinosoma Lang, 1944 et le genre Leptocaris T. Scott, 1899. Crustaceana 80(10):1153–1169. doi: 10.1163/156854007782321155.
Borutzky EV. 1952. Fauna SSSR. Rakoobraznye. Harpacticoida presnykh vod. Moscow: Zoologicheskii Institut Akademii Nauk SSSR. Boxshall GA, Halsey SH. 2004. An Introduction to Copepod Diversity. London: The Ray Society.
Ferrari FD, Ivanenko VN. 2008. The identity of protopodal segments and the ramus of maxilla 2 of copepods (Copepoda). Crustaceana 81(7):823–835. doi: 10.1163/156854008784771702.
Fiers F. 1986. Harpacticoid copepods from the West Indian Islands: Darcythompsoniidae (Copepoda, Harpacticoida). Bijdragen tot de Dierkunde 56:282–290.
Fleeger JW, Clark DR.1980. A revised key to Leptocaris (Copepoda: Harpacticoida), including a new species from a shallow estuarine Lake in Louisiana, USA. Northeast Gulf Science 3:53–59.
Fuentes-Reinés JM, Zoppi de Roa E. 2013a. Harpacticoid copepods from Ciénaga Grande de Santa Marta, Colombia. Métodos en Ecología y Sistemática 8:5–28.
Fuentes-Reinés JM, Zoppi de Roa E. 2013b. Occurrence of Elaphoidella grandidieri (Guerne and Richard, 1893) (Crustacea: Copepoda: Harpacticoida) in Ciénaga Grande de Santa Marta, Colombia. Check List 9(6):1580–1583. doi: 10.15560/9.6.1580.
Fuentes-Reinés JM, Gómez S. 2014. A new species of Schizopera (Copepoda: Harpacticoida: Miraciidae) from Colombia. Rev.MVZ Córdoba 19(3):4199–4213.
Fuentes-Reinés JM, Suárez-Morales E. 2014a. A new subspecies of Nitokra affinis Gurney, 1927 (Copepoda, Harpacticoida) from the Caribbean coast of Colombia. Zookeys 378:1–15. doi: 10.3897/zookeys.378.6695.
Fuentes-Reinés JM, Suárez-Morales E. 2014b. Annotated checklist and new records of Harpacticoida (Copepoda) from a coastal system of northern Colombia, South America. Crustaceana 87(2):212–255. doi: 10.1163/15685403-00003283.
Fuentes-Reinés JM, Zoppi de Roa E, Torres R. 2015. A new species of Cletocamptus Schmankewitsch, 1875 (Crustacea, Copepoda, Harpacticoida) and the description of the male of C. nudus from Colombia. Panam. J. Aquat. Sci. 10(1):1–18.
Gómez S. 2000. A new genus, a new species, and a new record of the family Darcythompsoniidae Lang, 1936 (Copepoda, Harpacticoida) from the Gulf of California, Mexico. Zool. J. Linn. Soc. 129(4): 515–536. doi: 10.1006/zjls.1999.0208.
Gómez S, Fuentes-Reinés JM. 2017. A new species of Tisbintra (Harpacticoida, Tisbidae) and range extension for Geehydrosoma brevipodum (Harpacticoida, Cletodidae) from northern Colombia. Caldasia 39(1):1–12. doi: 10.15446/caldasia.v39n1.64583.
Gee JM, Somerfield PJ. 1997. Do mangrove diversity and leaf litter decay promote meiofaunal diversity? J. Exp. Mar. Biol. Ecol. 218(1):13–33. doi: 10.1016/S0022-0981(97)00065-8.
Gurney R. 1920. A description of the copepod Cylindropsyllus brevicornis, Van Douwe, and of a new species of D’Arcythompsonia, Scott. Ann. Mag. Nat. Hist. Series 9 5(25):134–140. doi: 10.1080/00222932008632351.
Huys R. 2009. Unresolved cases of type fixation, synonymy and homonymy in harpacticoid copepod nomenclature (Crustacea: Copepoda). Zootaxa 2183:1–99.
Huys R, Boxshall GA. 1991. Copepod Evolution. London: The Ray Society.
Huys R, Suárez-Morales E, Serrano-Sánchez ML, Centeno-García E, Vega FJ. 2016. Early Miocene amber inclusions from Mexico reveal antiquity of mangrove-associated copepods. Sci Rep. 6:34872. doi: 10.1038/srep34872.
[ICZN] International Commission on Zoological Nomenclature. 1999 International code of zoological nomenclature. Fourth edition. London: The International Trust for Zoological Nomenclature.
Köroğlu NÖ, Kuru S, Karaytuğ S. 2014. Marine darcythompsoniids of the Turkish coasts with a description of Leptocaris emekdasi sp. nov. (Copepoda: Harpacticoida: Darcythompsoniidae) from the Aegean coast of Turkey. Mar. Biodiv. 45:383–390. doi: 10.1007/s12526-014-0295-3.
Kunz H. 1978. Zur Kenntnis der gattung Leptocaris (Copepoda, Harpacticoida, Darcythompsoniidae). Crustaceana 35(1):41–49. doi: 10.1163/156854078X00196.
Kunz H. 1983. Harpacticoiden (Crustacea: Copepoda) aus dem litoral der Azoren. Arquipelago 4:117–208.
Kunz H. 1994. Beitrag zur Kenntnis der Gattung Leptocaris (Copepoda, Harpacticoida) mit Beschreibung von zwei neuen Arten und zwei neuen Aunterarten. Mitt. Hambg. Zool. Mus. Inst. 91:35–52.
Lee JM, Chang CY. 2008. Copepods of the genus Leptocaris (Harpacticoida: Darcythompsoniidae) from Salt Marshes in South Korea. Korean J. Syst. Zool. 24(1):89–98. doi: 10.5635/KJSZ.2008.24.1.089.
Morrone JJ. 2014. Biogeographical regionalization of the neotropical region. Zootaxa 3782:1–110. doi: 10.11646/zootaxa.3782.1.1.
Por FD. 1983. Mangrove swamp-inhabiting Harpacticoida of the family Darcythompsonidae Lang. J. Crustac. Biol. 3:141–153.
Por FD, Prado-Por MSA, Oliveira EC. 1984. The mangal of the estuary and lagoon system of Cananeia (Brazil). In: Por FD, Por I, editors. Hydrobiology of the mangal. The Ecosystem of the Mangrove Forests. The Hague, Netherlands: Dr. W. Junk Publisher. p. 211–228.
Reid JW. 1998. Maxillopoda - Copepoda. Harpacticoida. In: Young PS, editor. Catalogue of Crustacea of Brazil. Rio de Janeiro: Museu Nacional. p. 75–127.
Smirnov S. 1934. Über eine neue D’Arcythompsonia-Art (Copepoda, Harpacticoida) aus dem Japanischen Meer. Zool. Anz. 108:118–122.
Song SJ, Dahms H-U, Khim JS. 2012. A review of Leptocaris including a description of L. ryukyuensis sp. nov. (Copepoda :Harpacticoida: Darcythompsoniidae). J. Mar. Biol. Assoc. U. K. 92(5):1073–1081. doi: 10.1017/S0025315411001949.
Suárez-Morales E, Fuentes-Reinés JM. 2015a. Two new species of ectinosomatid copepods (Harpacticoida: Ectinosomatidae) from the Caribbean coast of Colombia. Rev. Mex. Biodiv. 86(1):14–27. doi: 10.7550/rmb.46649.
Suárez-Morales E, Fuentes-Reinés JM. 2015b. A new species of Mesochra (Copepoda: Harpacticoida: Canthocamptidae) from a coastal system of northern Colombia with a key to the American species. J. Nat. Hist. 49(45–48): 2969–2982. doi: 10.1080/00222933.2015.1085604.
Zamudio-Valdéz JA, Reid JW. 1990. A new species of Leptocaris (Crustacea, Copepoda, Harpacticoida) from inland waters of Mexico. Anales del Instituto de Ciencias del Mar y Limnología 17: 47-54.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Juan M. Fuentes-Reinés, Eduardo Suárez-Morales, Pedro Eslava-Eljaiek, Daniel Serna-Macias, Deivis Suárez-Rivero. (2022). New records of Harpacticoids (Crustacea, Copepoda) from a coastal system of northern Colombia. Revista Peruana de Biología, 29(4), p.e23820. https://doi.org/10.15381/rpb.v29i4.23820.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2017 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















