



Publicado
Estructura ovárica y dinámica folicular de Liolaemus azarai (Squamata: Liolaemidae)
Ovarian structure and follicular dynamics of Liolaemus azarai (Squamata: Liolaemidae)
DOI:
https://doi.org/10.15446/caldasia.v39n2.64967Palabras clave:
Atresia, cuerpos lúteos, folículos, lagartija, ovario (es)Atresia, corpora lutea, follicles, lizard, ovary (en)
El estudio de la estructura ovárica y sus variaciones estacionales resulta fundamental para interpretar las estrategias reproductivas de los reptiles. Se analizó la morfología e histología del ovario de Liolaemus azarai a fin de caracterizar sus cambios y ofrecer información para determinar su ciclo reproductivo. Se registró, un nido germinal por ovario, con ovogonias y ovocitos primarios lo que evidencia que la capacidad proliferativa de la línea germinal se mantiene en el estado adulto. Durante la previtelogénesis, en todas las hembras capturadas, se verificó un incremento de hasta 30 veces el tamaño del ovocito debido al aumento del contenido citoplasmático, distinguiéndose una capa folicular con tres tipos celulares. La vitelogénesis se caracterizó por la incorporación del vitelo al ovocito y por las modificaciones que sufrió el epitelio folicular, el cual quedó formado por células planas, esta etapa se observó en septiembre, octubre, noviembre y diciembre. Se observaron cuerpos lúteos en estado avanzado del desarrollo, en octubre y en diciembre, coincidiendo con el grado de desarrollo embrionario intrauterino. Folículos atresicos se observaron en octubre y diciembre. La atresia temprana se evidenció fagocitosis del ovocito, abundantes células foliculares e invasión de células sanguíneas, mientras que en estados de atresia avanzada se observó la presencia de células autofágicas. Los resultados constituyen el primer aporte al conocimiento de la dinámica folicular de L. azarai y servirán como base para interpretar el ciclo reproductivo y desarrollo embrionario aún desconocidos para la especie.
The knowledge of ovarian morphology and its seasonal variations is essential to interpret the reproductive strategies of reptiles. The ovarian morphology and histology of Liolaemus azarai, was analyzed in order to characterize its changes and to provide information to determine their reproductive cycle. One germinal bed per ovary with oogonia and primary oocytes, it demonstrates that the proliferative activity of the germ line is maintained in adult. During previtellogenic stages, all of the females captured, the oocyte grows near 30 times due to increase of cytoplasmic contents, and follicular layer with three types of cells develops. The vitellogenic stages are characterized by the incorporation of the yolk platelets and important modifications in the follicular epithelium, is organized in a layer of flats cells, this time were observed in September, October, November and December. The corpora lutea was observed in an advanced stage of development, in October and December, according to the degree of intrauterine embryonic development. The atretic follicles were observed in October and December. The early atretic showed oocyte phagocytosis, abundant follicular cells and invation of blood cells; the advanced atretic follicles were characterized by autophagic cells. These results are the first contribution to the knowledge of follicular dynamics of L. azarai, and will be useful for to interpret the unknown reproductive cycle and embryonic developmental of this specie.
Recibido: 2 de junio de 2017; Aceptado: 2 de octubre de 2017
RESUMEN
El estudio de la estructura ovárica y sus variaciones estacionales resulta fundamental para interpretar las estrategias reproductivas de los reptiles. Se analizó la morfología e histología del ovario de Liolaemus azarai a fin de caracterizar sus cambios y ofrecer información para determinar su ciclo reproductivo. Se registró, un nido germinal por ovario, con ovogonias y ovocitos primarios lo que evidencia que la capacidad proliferativa de la línea germinal se mantiene en el estado adulto. Durante la previtelogénesis, en todas las hembras capturadas, se verificó un incremento de hasta 30 veces el tamaño del ovocito debido al aumento del contenido citoplasmático, distinguiéndose una capa folicular con tres tipos celulares. La vitelogénesis se caracterizó por la incorporación del vitelo al ovocito y por las modificaciones que sufrió el epitelio folicular, el cual quedó formado por células planas, esta etapa se observó en septiembre, octubre, noviembre y diciembre. Se observaron cuerpos lúteos en estado avanzado del desarrollo, en octubre y en diciembre, coincidiendo con el grado de desarrollo embrionario intrauterino. Folículos atresicos se observaron en octubre y diciembre. La atresia temprana se evidenció fagocitosis del ovocito, abundantes células foliculares e invasión de células sanguíneas, mientras que en estados de atresia avanzada se observó la presencia de células autofágicas. Los resultados constituyen el primer aporte al conocimiento de la dinámica folicular de L. azarai y servirán como base para interpretar el ciclo reproductivo y desarrollo embrionario aún desconocidos para la especie.
Palabras clave:
Atresia, cuerpos lúteos, folículos, lagartija, ovario.ABSTRACT
The knowledge of ovarian morphology and its seasonal variations is essential to interpret the reproductive strategies of reptiles. The ovarian morphology and histology of Liolaemus azarai, was analyzed in order to characterize its changes and to provide information to determine their reproductive cycle. One germinal bed per ovary with oogonia and primary oocytes, it demonstrates that the proliferative activity of the germ line is maintained in adult. During previtellogenic stages, all of the females captured, the oocyte grows near 30 times due to increase of cytoplasmic contents, and follicular layer with three types of cells develops. The vitellogenic stages are characterized by the incorporation of the yolk platelets and important modifications in the follicular epithelium, is organized in a layer of flats cells, this time were observed in September, October, November and December. The corpora lutea was observed in an advanced stage of development, in October and December, according to the degree of intrauterine embryonic development. The atretic follicles were observed in October and December. The early atretic showed oocyte phagocytosis, abundant follicular cells and invation of blood cells; the advanced atretic follicles were characterized by autophagic cells. These results are the first contribution to the knowledge of follicular dynamics of L. azarai, and will be useful for to interpret the unknown reproductive cycle and embryonic developmental of this specie.
Key words:
Atresia, corpora lutea, follicles, lizard, ovary.INTRODUCCIÓN
El estudio de la estructura ovárica y sus variaciones estacionales resulta fundamental para interpretar las características del ciclo reproductivo de una especie. La dinámica ovárica comprende distintos eventos como proliferación de ovogonias, foliculogénesis, crecimiento folicular, maduración del ovocito (previtelogénesis y vitelogénesis), atresia folicular, ovulación, y formación del cuerpo lúteo (Palmer et al. 1997). Dichos eventos implican cambios anatómicos e histológicos, verificables por aumento de tamaño de los folículos y modificaciones del epitelio folicular, entre otras, a lo largo del ciclo reproductivo.
Entre los reptiles existen marcadas diferencias en la morfología folicular. En Testudines y Crocodilia el epitelio folicular es cúbico simple (Callebaut et al. 1997, Romero De Pérez et al. 1999, Uribe y Guillette 2000, Calderón et al. 2004), mientras que en Squamata es polimórfico estratificado (Uribe et al. 1995, 2010) con variaciones a lo largo del ciclo reproductivo.
De todos los reptiles, el grupo de los escamados es el que presenta mayor variedad de modelos reproductivos (Ibargüengoytía 2008), por lo que los conocimientos que puedan generarse acerca de la morfología y dinámica ovárica tienen gran relevancia ya que aportan información de base para interpretar la actividad reproductiva.
El género Liolaemus se caracteriza por presentar una gran variedad de estrategias reproductivas incluyendo especies con ciclos anuales, bianuales o trienales, estacionales o continuos, sincrónico o asincrónicos entre sexos, con formas sexuales y partenogenéticas, con desarrollo ovíparo o vivíparo (Ibargüengoytía 2008). Esta particularidad ha impulsado a Schulte et al. (2000) a proponer al género como un excelente modelo para analizar la evolución de los patrones reproductivos. Liolaemus azarai Ávila, 2003 es una lagartija de pequeño tamaño cuya longitud hocico cloaca máxima es de 48,7 y 54,3 mm, para adultos hembras y machos, respectivamente. Presenta dimorfismo sexual en coloración y tamaño. Tiene desarrollo ovíparo y fecundidad de hasta cuatro huevos (Ávila 2003). Esta especie tiene una distribución restringida. Ha sido citada para Isla Yaciretá (Departamento Itapúa), San Sebastián (Departamento Ñeembucú) y Yabebyry (Departamento Misiones) en Paraguay (Zaracho et al. 2017) y para Isla Apipé (Departamento Ituzaingó), Curuzú Laurel (Departamento San Miguel) y Concepción (Departamento Concepción), en Corrientes, Argentina (Álvarez et al. 2003, Zaracho et al. 2014). Habita en dunas y suelos arenosos con vegetación principalmente de Butia paraguayensis (Barb. Rodr.) L.H. Bailey. Por sus características ecomorfológicas y su distribución restringida, ha sido categorizada como Vulnerable (Abdala et al. 2012). Debido a sus hábitos de vida y escasos registros en colecciones herpetológicas, poco se conoce de su biología y particularmente su reproducción. En el presente trabajo se describen los cambios en la morfología ovárica de hembras de L. azarai, a fin de contribuir con información de base que permita interpretar su ciclo reproductivo.
MATERIALES Y MÉTODOS
Obtención de ejemplares
Se utilizaron 15 hembras adultas de L. azarai de tamaño igual o superior 35 mm, en etapa no reproductiva (n: 3) y reproductiva (n: 12) de su ciclo de vida. Las mismas fueron capturadas en la Isla Apipé Grande (27°29' Sur, y 56°54' Oeste) en la Departamento de Ituzaingó, y en Curuzú Laurel (27°56' Sur, y 57°30' Oeste) Departamento de San Miguel, Corrientes, Argentina, durante el periodo 2007-2013 abarcando otoño (marzo y abril) primavera (septiembre, octubre y noviembre) y verano (diciembre).
La colecta de los ejemplares se realizó con la correspondiente autorización de Colecta Científica, dando cumplimiento a la disposición N° 845 de la Dirección de Recursos Naturales del Ministerio de Producción, Trabajo y Turismo de la provincia de Corrientes, Argentina. Los ejemplares fueron sacrificados por sobredosis de Carticaina-L-adrenalina, siguiendo las recomendaciones propuestas por The Herpetological Animal Care and Use Committee (American Society of Ichthyologists and Herpetologists) (Beaupre et al. 2004). Posteriormente fueron fijados en solución de Bouin por 12 horas y lavados en alcohol 70 %. Se midió la Longitud Hocico Cloaca (LHC) de cada ejemplar analizado con calibrador digital de 0,01 mm de precisión. El análisis macroscópico de los ovarios se realizó con lupa estereoscópica Leica ES2 (Wetzlar, Alemania). Se registraron los diámetros de todos los folículos ováricos observados utilizando un ocular micrométrico incorporado a los instrumentos ópticos y un calibrador digital de 0,01 mm de precisión. El ovario derecho de cada ejemplar fue aislado y procesado para la obtención de muestras histológicas.
Obtención de muestras histológicas
Los preparados histológicos se realizaron de acuerdo al protocolo convencional de deshidratación en soluciones crecientes de alcohol etílico (70, 80, 96 y 100 %), alcohol butílico 100 %, impregnación en alcohol butílico-parafina (50-50 %) e inclusión en parafina (100 %). Se realizaron cortes seriados de 7 μm, con micrótomo rotatorio tipo Spencer, los cuales fueron montados y teñidos con hematoxilina-eosina (H-E) y PAS. Las muestras se observaron mediante microscopio óptico (MO) Leica DM500 con cámara Leica ICC50HD (Wetzlar, Alemania). Se utilizó el software LAZ EZ.INK para la captura de las imágenes y medición del diámetro de ovogonias, ovocitos y folículos, en diferentes estadios, observados. Se realizó la caracterización histológica y la clasificación de los estadios de la ovogénesis y foliculogénesis siguiendo la clasificación propuesta por Uribe et al. (1995). Los estadios de los cuerpos lúteos se identificaron a partir de lo descripto por Martínez-Torres et al. (2003). Los estadios embrionarios (E.) fueron determinados a partir de la tabla de Dufaure y Hubert (1961) (D-H).
Los ejemplares analizados se incorporaron a la Colección Herpetológica de la Universidad Nacional del Nordeste (Corrientes, Argentina) (UNNEC: 7545, 9656, 9662, 9770, 9773, 10181, 10182, 10364, 11029, 11030, 11031, 11102, 11105, 11201, 11382).
RESULTADOS
Análisis macroscópico del ovario
Las hembras de L. azarai analizadas midieron en promedio 44,2 ± 6,8 mm de LHC. Presentaron ovarios pares, de posición asimétrica, siendo el derecho más craneal que el izquierdo. Los ovarios presentaron contorno irregular debido a la presencia de folículos en distintas fases de crecimiento. A nivel macroscópico se reconocieron sólo folículos previtelogénicos en hembras en etapa no reproductiva, y folículos previtelogénicos, vitelogénicos, cuerpos lúteos y folículos atrésicos durante el periodo de actividad reproductiva (Fig. 1a). Los folículos previtelogénicos (PV) se caracterizaron por su pequeño tamaño, no mayor a 1,5 mm de diámetro, y su aspecto translúcido o coloración blanquecina. Los folículos vitelogénicos (FV) fueron de tamaño y coloración variable según su etapa de maduración. Los FV iniciales alcanzaron hasta 3 mm de diámetro y fueron de color amarillo pálido. Los FV avanzados, cuyo diámetro varió entre 3,1 y 8,3 mm, tenían color amarillo-naranja intenso. Los folículos atrésicos se reconocieron por su forma irregular y aspecto translúcido. Los cuerpos lúteos tuvieron contorno irregular, coloración amarillo-naranja y presencia de una cicatriz longitudinal correspondiente al estigma.
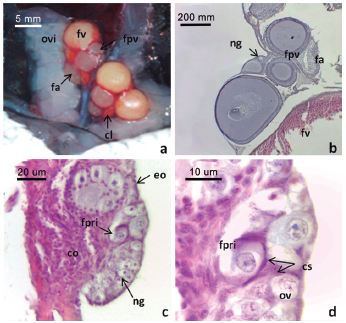
Figura 1: Morfología de sistema reproductor femenino y secciones histológicas del ovario de L. azarai.
a. Vista general del sistema reproductor en etapa reproductiva. b. Sección histológica del ovario en etapa reproductiva. c. Nido germinal. d. Detalle de un folículo primordial en etapas iniciales de la foliculogénesis. Convenciones: cl: cuerpo lúteo, co: corteza ovárica, cs: células de soporte, eo: epitelio ovárico, fa: folículo atrésico, fpri: folículo primordial, fpv: folículo previtelogénico, fv: folículo vitelogénico, ng: nido germinal, o: ovogonias, ov: ovocito primario, ovi: oviducto. Tinción: H-E.
Análisis histológico del ovario
El ovario presentó una corteza y una médula, rodeadas por un epitelio ovárico superficial plano simple. La médula fue lacunar con escaso tejido conjuntivo. La corteza ovárica presentó un nido germinal y folículos previtelogénicos en la etapa no reproductiva de los ejemplares. En la etapa reproductiva se reconocieron, además, folículos vitelogénicos, folículos atrésicos o cuerpos lúteos (Fig. 1b).
Nido germinal
Se observó un nido germinal por ovario en el epitelio de la cara dorsal del mismo (Fig. 1c). Estuvo formado por un número variable de ovogonias y ovocitos primarios en distintas fases de la primera división meiótica, intercalados con células de soporte o prefoliculares (Fig. 1d). Las ovogonias (7,4 ± 1,8 μm de diámetro, n = 10) fueron células voluminosas claras, con núcleo de posición central, con cromatina laxa y un nucléolo. Los ovocitos primarios, se diferenciaron de las ovogonias por su mayor tamaño (8,7 ± 1,5 μm de diámetro, n = 5) y por el aspecto fibrilar de la cromatina. Las células de soporte o prefoliculares fueron células eosinófilas, pequeñas, de contorno y núcleo ovoide con un nucléolo. Se observaron nidos germinales en todos los ovarios analizados.
Folículos previtelogénicos
Los folículos primordiales (estadio I) (14,9 ± 1,6 μm de diámetro, n = 4) estuvieron formados por el ovocito (8,9 ± 1,8 μm de diámetro, n = 4) en arresto meiótico en fase de diploteno, rodeado por una capa de células foliculares planas. Se localizaron en la periferia de la corteza ovárica, adyacentes a los nidos germinales (Fig. 1d, Fig. 2a).
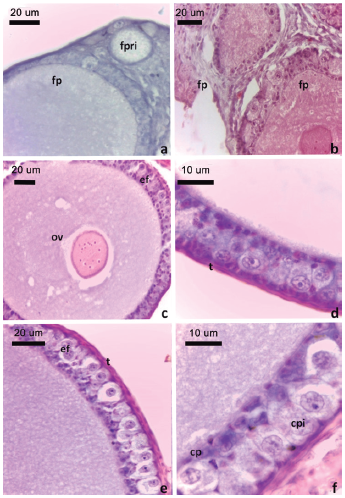
Figura 2: Folículos previtelogénicos de L. azarai.
a. Folículo primario estadio IIa, b. Folículos primarios estadio IIb, c. Folículo en estadio IIIa, d. Detalle del epitelio folicular de un folículo en estadio IIIb. e. Detalle del epitelio folicular de un folículo en estadio IV. f. Detalle de las células piriformes. Obsérvese la prolongación de la porción apical de la célula piriforme en contacto con la membrana plasmática del ovocito (flecha). Convenciones: cp: células pequeñas, cpi: células piriformes, ef: epitelio folicular, fp: folículo primario, fpri: folículo primordial, fpv: folículo previtelogénico, ov: ovocito, t: teca. Tinción H-E.
Los folículos primarios (estadio II) presentaron diferencias en cuanto al tamaño del ovocito y organización del epitelio folicular, diferenciándose en estadio IIa y IIb. Los folículos primarios en estadio IIa presentaron un epitelio folicular simple de células cúbicas (Fig. 2a) y los folículos primarios en estadio IIb presentaron una capa de células foliculares de diámetro variable (Fig. 2b).
Los folículos previtelogénicos, estadios III y IV, estuvieron formados por el ovocito primario en diploteno rodeado por un epitelio folicular cúbico estratificado y por una a dos capas de células tecales planas. Los folículos en estadio III presentaron tamaños variables entre 54,2 y 247,6 μm (n = 12). De acuerdo con características morfológicas del ovocito y del epitelio folicular se diferenciaron en dos tipos. El estadio IIIa (54,2 a 73,4 μm de diámetro), presentó el ovocito (45,3 ± 2,8 μm de diámetro, n = 9) con ovoplasma claro y vacuolado, el núcleo fue central con múltiples nucléolos. El epitelio folicular estuvo formado por células de diferentes tamaños (Fig. 2c). El estadio IIIb, el ovocito (146,7 ± 48,8 μm de diámetro, n = 3) presentó el ovoplasma perinuclear vacuolado y estuvo rodeado por la membrana vitelina PAS positiva. El epitelio folicular se organizó en tres estratos, uno interno formado por las células pequeñas (5,7 ± 0,85 μm de diámetro, n = 15) levemente eosinófilas y núcleo con un nucléolo, un estrato medio de células grandes (10,1 ± 0,9 μm de diámetro, n = 13), de ovoplasma claro y núcleo central voluminoso con varios nucléolos y un estrato externo de células pequeñas (Fig. 2d).
En el estadio IV el diámetro folicular promedio fue de 413,2 ± 5,4 μm (n = 4). Los ovocitos presentaron un diámetro promedio de 394,6 ± 56 μm (n = 4) y un núcleo de posición central con varios nucléolos. El ovoplasma perinuclear fue irregular y de aspecto vacuolado mientras que el ovoplasma cortical fue homogéneo. El epitelio folicular estuvo organizado en dos estratos, uno interno de células pequeñas y uno externo de células grandes piriformes, de núcleo voluminoso con un nucléolo prominente. Estas células contactaron mediante su extremo ensanchado con la membrana basal del epitelio folicular, y con la membrana plasmática del ovocito mediante su extremo apical (Figs. 2e-f). Los folículos primordiales y previtelogénicos se observaron en todas las hembras.
Folículos vitelogénicos
Los folículos vitelogénicos correspondieron a los estadios V, VI, VII y VIII. En el estadio V se observaron pequeños gránulos eosinófilos en el ovoplasma cortical, próximos a la membrana vitelina, correspondientes a plaquetas de vitelo. La membrana vitelina fue delgada, de aspecto homogéneo y refringente al microscopio de campo claro (Fig. 3a). La teca estuvo constituida por fibroblastos, matriz colágena y pequeños vasos sanguíneos.
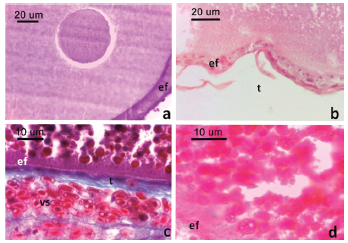
Figura 3: Folículos vitelogénicos de L. azarai.
a. Folículo en estadio V. Obsérvese los gránulos en la región cortical del citoplasma (flecha). b. Detalle del epitelio folicular de un folículo en estadio VI. c. Detalle del epitelio folicular de un folículo en estadio VII. d. Detalle la región cortical del ovocito de un folículo en estadio VIII. Obsérvese las plaquetas vitelinas. Convenciones: ef: epitelio folicular, pv: plaquetas vitelinas, t: teca, vs: vaso sanguíneo. Tinción H-E (a., b. y d.) y tricrómica de Gomori (c.).
Folículos en estadio VI (754,7 цгп de diámetro, n = 1), el ovocito (714,9 μm de diámetro, n = 1) presentó abundantes plaquetas vitelinas distribuidas en ovoplasma, siendo las corticales de menor tamaño que las localizadas en el ovoplasma central. El epitelio folicular estuvo formado por una o dos capas de células cúbicas de tamaño semejante, y no se observaron células piriformes (Fig. 3b). La teca fue semejante al estadio anterior.
Folículos en estadio VII presentaron el ovocito (1,3538 ± 74,3 mm de diámetro, n = 3) con plaquetas vitelinas organizadas centrípetamente de menor a mayor tamaño, excepto la región cortical que se observaron como una zona más clara y homogénea. El epitelio folicular estuvo formado por células planas (Fig. 3c). La teca se caracterizó por presentar varias capas de fibroblastos y abundantes vasos sanguíneos.
Folículos en el estadio VIII, el ovocito (3760 μm de diámetro, n = 1) presentó un núcleo excéntrico, próximo a la membrana plasmática, y ovoplasma completamente ocupado por plaquetas vitelinas e inclusiones lipídicas. El epitelio folicular tuvo una capa de células planas (Fig. 3d). La teca estuvo muy vascularizada.
Los folículos vitelogénicos fueron observados en hembras colectadas durante los meses de septiembre, octubre, noviembre y diciembre.
Cuerpo lúteo
Se observaron cuerpos lúteos en estadio III y IV, en dos hembras que poseían huevos oviductales. Cuerpos lúteos en estadio III (Fig. 4a) presentaron estructura compacta y contorno irregular. Se observó una zona interna de células luteínicas de forma oval y una zona externa de células tecales hipertrofiadas, inmersas en una matriz extracelular acidófila, y abundantes vasos sanguíneos (Fig. 4b). Estos fueron registrados en una hembra colectada en diciembre con tres embriones en estadio de gástrula inicial (E. 11 D-H).
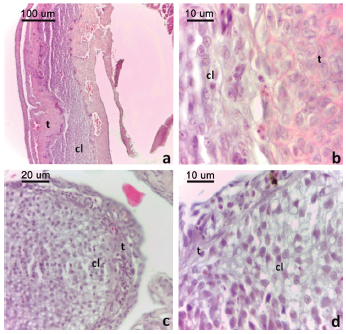
Figura 4: Cuerpo lúteo de L. azarai.
a. Cuerpo lúteo en fase III. b. Detalle de Fig. 4a. c. Cuerpo lúteo en fase IV. d. Detalle de 4c. Convenciones: cl: células luteínicas, t: teca. Tinción H-E.
En los cuerpos lúteos en estadio IV (Fig. 4c) se observaron células luteínicas invadidas por vasos sanguíneos y por septos de tejido conectivo de matriz acidófila. Las células luteínicas, de forma redondeada, contenían abundantes vacuolas hialinas e inclusiones acidófilas (Fig. 4d). Este estadio se registró en una hembra colectada en octubre con cuatro embriones en organogénesis temprana (E. 24-25 D-H).
Folículos atrésicos
Se observaron folículos en etapa temprana y avanzada del proceso de atresia. El folículo en atresia temprana (Fig. 5a), correspondiente a un folículo previtelogénico, presentó forma irregular con abundantes células foliculares fagocíticas, invadidas por células sanguíneas, y rodeadas por células tecales. Se observó una avanzada fagocitosis del ovoplasma, en el cual no se distinguió membrana vitelina ni membrana plasmática (Fig. 5b). Entre las células sanguíneas se observaron eritrocitos y leucocitos destacándose la presencia de eosinófilos (Fig. 5c).
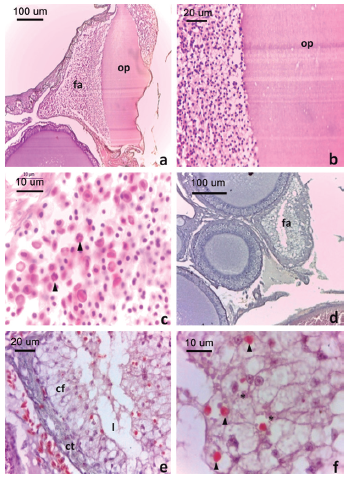
Figura 5: Folículos atrésicos de L. azarai.
a. Folículo en atresia temprana. Obsérvese la desorganización e hipertrofia de las células foliculares y rotura de la membrana plasmática del ovocito. b. Detalle las células foliculares invadiendo y fagocitando el citoplasma (flecha). c. Detalle de la invasión de células sanguíneas en el folículo atrésico (flecha indica eosinófilos). d. Folículo en atresia avanzada. e. Detalle de folículo atrésico. Obsérvese la reabsorción completa del ovoplasma. f. Detalle de las células foliculares de un folículo atrésico avanzado, obsérvese gránulos acidófilos (flecha) e inclusiones basófilas (asteriscos). Convenciones: cf: células foliculares, ct: células tecales, fa: folículo atrésico, l: lumen, op: ovoplasma. Tinción H-E.
Los folículos en atresia avanzada (Fig. 5d) presentaron forma irregular, un lumen reducido con restos de ovoplasma, células foliculares y células tecales. Las células foliculares estuvieron hipertofiadas, tuvieron forma irregular y presentaron vacuolas voluminosas que ocupan casi la totalidad del citoplasma (Fig. 5e) similar al estado de autofagia. Algunas de estas células presentaron, además, gránulos acidófilos y pequeñas inclusiones basófilas, compatibles con cuerpos apoptóticos (Fig. 5f). Entre las células tecales se observaron escasas células sanguíneas, principalmente eritrocitos y algunos eosinófilos. Los folículos atrésicos se registraron en hembras con folículos vitelogénicos colectadas en octubre y diciembre respectivamente.
DISCUSIÓN
Uno de los aspectos principales a considerar en el estudio de la biología reproductiva de los reptiles es la dinámica folicular a lo largo del ciclo reproductivo. El conocimiento de la morfología y fisiología ovárica es importante porque permite interpretar, no solo los mecanismos que regulan la actividad reproductiva de las hembras, sino también aspectos relacionados con el desarrollo embrionario. En cuanto a la morfología ovárica y folicular, L. azarai no mostró diferencias con lo descripto para otras especies del género, con las que difiere en su modo reproductivo (Ramírez Pinilla 1991, 1995), ni para otros lagartos (Uribe et al. 1996, 1998, Manes et al. 2007, Uribe et al. 2010).
En los reptiles la ovogénesis se mantiene durante toda la vida reproductiva de las hembras gracias a la presencia de nidos germinales de ovogonias con capacidad proliferativa. Jones y Sumers (1984) registran que el número de ovogonias en Hemidactylus frenatus Dumeril y Bribon, 1836 varía significativamente durante el ciclo reproductivo, siendo más bajo en la fase luteal. Guraya (1989), propone que las ovogonias tendrían un pico de actividad mitótica estacional postovulación, seguido de eventos de meiosis y foliculogénesis. En L. azarai se observaron nidos germinales con escasas ovogonias y folículos primordiales y primarios en hembras en fase vitelogénica. Hembras en fase luteínica muestran predominio de ovogonias siendo escaso el número de los ovocitos, lo que podría inferirse como un evento de proliferación e inicio de meiosis postovulación.
Jones et al. (1982) describen que los lagartos pueden tener uno, dos o varios nidos germinales, dependiendo de la fecundidad, siendo un carácter especieespecífico. Anolis sagrei Dúmeril y Bribon, 1837 (Sanz-Ochotorena y Uribe 1999) y A. porcatus Gray, 1840 (Sanz-Ochotorena et al. 2000, 2005) presentan un nido germinal por ovario y liberan un ovocito en cada ciclo reproductivo. Para Calotes versicolor Daudin, 1802, Shanbhag y Prasad (1993) reportan fecundidad de entre 13 y 24 huevos y describen la presencia de dos nidos germinales por ovario. Ctenosaura pectinata Wiegmann, 1834 (Uribe et al. 1996), cuya fecundidad puede ser de hasta 50 huevos y Mabuya brachypoda Taylor, 1956 (Hernández-Franyutti et al. 2005) que pare una sola cría por ciclo reproductivo, presentan dos nidos germinales por ovario (Ramírez-Bautista y González-Romero 2002). Lozano et al. (2014) describen para Sceloporus grammicus Wiegmann, 1828, la presencia de dos nidos germinales por ovario y la liberación de múltiples ovocitos por ovulación. Para el género Liolaemus dos nidos germinales por ovario es un rasgo común (Ramírez Pinilla 1991). Para L. azarai se registró una fecundidad máxima de cuatro huevos y un nido germinal por ovario lo que difiere de lo reportado para otras especies del género. De lo expuesto se destaca que, dentro de los lagartos, el número de huevos varía independientemente de la presencia de uno o dos nidos germinales por ovario. Según esto, la fecundidad no estaría condicionada por la cantidad de nidos germinales sino por la actividad mitótica de las ovogonias, análisis que requiere ser abordado en futuras investigaciones.
El desarrollo del epitelio folicular inicia a partir de la asociación temprana de células foliculares. En la fase de previtelogénesis hay un marcado crecimiento de la célula germinal, debido principalmente al aumento del contenido citoplasmático, e importantes cambios del epitelio folicular (Callebaut et al. 1997). Klosterman (1987) analiza Gerrhonotus coeruleus Wiegmann, 1828 e identifica, en las células intermedias y piriformes, características estructurales indicativas de intensa actividad de síntesis. Motta et al. (1995) describen que las células piriformes establecen puentes citoplasmáticos con el ovocito comportándose como células nodrizas que sintetizan ARN ribosómico y pequeñas moléculas que son transportadas hasta la célula germinal. Estos eventos contribuirían al aumento del contenido citoplasmático del ovocito, registrado en este estudio, el cual incrementó su diámetro hasta aproximadamente 30 veces. En L. azarai a partir del estadio III las células foliculares proliferan, diferenciándose células piriformes en el estadio IV lo que estaría indicando actividad de síntesis de moléculas.
La fase vitelogénica se caracteriza por un aumento exponencial del tamaño del ovocito, debido a la acumulación de plaquetas vitelinas e inclusiones lipídicas. Durante esta etapa, en L. azarai, el epitelio folicular presentó marcados cambios estructurales pasando de ser una capa estratificada polimórfica a constituirse como una monocapa de células planas. Motta et al. (1996, 2002) y Tammaro et al. (2006), describen regresión de las células piriformes al final de la previtelogénesis por procesos de apoptosis. El adelgazamiento de la capa folicular durante la vitelogénesis observado en L. azarai podría deberse a eventos de apoptosis como lo postulan dichos autores. Sin embargo, no se han registrado células con características apoptóticas en el epitelio folicular, por lo que es necesario profundizar este estudio incluyendo la detección de proteínas asociadas al proceso de apoptosis.
El cuerpo lúteo juega un rol fundamental en la producción de progesterona durante la gestación (Motta et al. 2002). Dentro de los reptiles existe gran variabilidad en cuanto a su morfología, su secreción y el tiempo de regresión, en relación con el modelo de desarrollo embrionario (Goldberg 1970, Guillete et al. 1981). En los escamados ovíparos está activo durante el periodo de gravidez y su regresión es previa a la oviposición, mientras que en las especies vivíparas su actividad es muy variable (Martínez-Torres et al. 2003). La estructura de los cuerpos lúteos de L. azarai fue semejante a la descripta para otros lagartos (Guillette et al. 1981, Gómez y Ramírez 2004, Manes et al. 2007), cuerpos lúteos en estadio III corresponden a un estado de desarrollo embrionario inicial, mientras que el estadio IV, donde se evidencia la máxima actividad secretoria, corresponde a la fase de organogénesis temprana. Con respecto a esto sería interesante profundizar el análisis de la actividad del cuerpo lúteo, su función en el mantenimiento de la retención intrauterina de huevos y la relación entre su regresión y la oviposición.
La atresia folicular es un proceso degenerativo, regulado por factores hormonales o ambientales, por medio del cual se eliminan folículos ováricos (Santos et al. 2008). Shanbhag y Prasad (1993) plantean que en los lagartos la atresia es más frecuente en folículos previtelogénicos, sugiriendo que estos serían más propensos a degenerar. En L. azarai se registró atresia en folículos previtelogénicos, aunque no se pudo determinar a qué estadio correspondían, en hembras preovulatorias. Por otra parte, en folículos atrésicos avanzados se observaron pequeñas inclusiones acidófilas en las células foliculares, comparables a las plaquetas vitelinas. Esto podría indicar un proceso de fagocitosis de dichas plaquetas, lo que demostraría eventos de atresia en folículos vitelogénicos.
Makrigiannakis et al. (2000) y Hussein (2005) sugieren que la apoptosis es el principal mecanismo por medio del cual se eliminan las células de la granulosa durante el proceso de atresia. Santos et al. (2008) analizan los eventos de atresia en Prochilodus argenteus Agassiz, 1829 y Leporinus taeniatus Lütken, 1875 y concluyen que la heterofagia, la autofagia y la apoptosis constituyen la secuencia de eventos característicos de la atresia folicular en teleósteos. En L. azarai, los folículos atrésicos tempranos presentaron abundantes células foliculares fagocíticas invadiendo el ovoplasma, comparable a un proceso de heterofagia. En la atresia avanzada las células foliculares mostraron características similares a células en procesos de muerte celular programada. Estas observaciones coinciden con lo reportado por Thomé et al. (2009) y Choi et al. (2010) y constituyen una evidencia concordante con la hipótesis de que la autofagia sería un evento previo a la apoptosis durante la atresia folicular. Sin embargo, es necesario corroborar esta información con la detección de proteínas implicadas en la muerte celular programada.
Este trabajo constituye la primera contribución al conocimiento de la actividad y dinámica ovárica de L. azarai a lo largo del ciclo reproductivo. En todos los meses del año se registraron ovogonias, ovocitos primarios y folículos previtelogénicos. La vitelogénesis ocurrió entre septiembre y diciembre, y a partir de octubre se registraron hembras grávidas con cuerpos lúteos. La atresia folicular se evidenció en folículos previtelogénicos y vitelogénicos, durante el periodo de actividad reproductiva. Estos resultados permiten inferir que el ciclo reproductivo de hembras de L. azarai correspondería a un patrón estacional.
PARTICIPACIÓN DE AUTORES
AMB Diseño, toma de datos, análisis, escritura del documento; SMT Diseño, análisis, escritura del documento; ABB Concepción y escritura del documento.
AGRADECIMIENTOS
Queremos extender nuestro agradecimiento a los revisores de dicho manuscrito por el aporte al mismo. A la Dirección de Recursos Naturales del Ministerio de Producción, Trabajo y Turismo de la provincia de Corrientes Argentina, por otorgar los correspondientes permisos de colecta científica y guía de tránsito para la colecta del material biológico. Un especial agradecimiento a los revisores anónimos del manuscrito, por los aportes y comentarios realizados. El presente trabajo fue financiado por la SGCyT-UNNE Proyecto 17/12F00 Taxonomía, biodiversidad y conservación de da herpetofauna de Corrientes, Chaco y Formosa (Argentina).
LITERATURA CITADA
Referencias
Abdala CS, Acosta JL, Acosta JC, Álvarez BB, Arias F, Avila LJ, Blanco MG, Bonino M, Boretto JM, Brancatelli G, Breitman MF, Cabrera MR, Cairo S, Corbalán V, Hernando A, Ibargüengoytía NR, Kacoliris F, Laspiur A, Montero R, Morando M, Pelegrin N, Pérez CHF, Quinteros AS, Semhan RV, Tedesco ME, Vega L, Zalba SM. 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuad. Herpetol. 26(1):215–248.
Álvarez BB, Aguirre R, Céspedez J, Hernando AB, Tedesco ME. 2003. Herpetofauna del Iberá: En: Álvarez BB, editores. Fauna del Iberá. Corrientes, Argentina: Editorial de la Universidad Nacional del Nordeste. p. 99–178.
Ávila LJ. 2003. New species of Liolaemus (Squamata: Liolaemidae) from Northeastern Argentina and Southern Paraguay. Herpetologica. 59(2):283–292. doi: 10.1655/0018-0831.
Beaupre SB, Jacobson ER, Lillywhite HB, Zamudio K. 2004. Guidelines for use of live amphibians and reptiles in field and laboratory research. 2nd Edition. Lawrence, Kansas: Herpetological Animal Care and Use Committee of the American Society of Ichthyologists and Herpetologists.
Calderón ML, De Pérez GR, Ramírez-P MP. 2004. Morphology of the ovary of Caiman crocodilus (Crocodylia: Alligatoridae). Ann. Anat. 186:13–24. doi: 10.1016/S0940-9602(04) 80113-4.
Callebaut M, Van Nassauw L, Harrison F. 1997. Comparison between oogenesis and related ovarian structures in a reptile, Pseudemys scripta elegans (turtle) and in a bird Coturnix coturnix japonica (quail). Reprod. Domest. Anim. 37:233–252.
Choi JY, Jo MW, Lee EY, Yoon BK, Choi DS. 2010. The role of autophagy in follicular development and atresia in rat granulosa cells. Fertil. Steril. 93(8):2532–2537. doi: 10.1016/j.fertnstert.2009.11.021.
Dufaure JP, Hubert J. 1961. Table de developpement du lezard vivipare: Lacerta (Zootoca) vivipara Jacquin. Arch. Anat. Microsc. Morphol. Exp. 50(3):309–328.
Goldberg SR. 1970. Seasonal ovarian histology of the ovoviparous iguanid lizard Sceloporus jarrovi cope. J. Morphol. 132(3):265–275. doi:10.1002/jmor.1051320303.
Gomez D, Rámirez-Pinilla MP. 2004. Ovarían histology of viviparous matrotrophic lizard Mabuya mabuya (Squamata: Scincide). J. Morphol. 259:90–105. doi: 10.1002/jmor.10170.
Guillette LJ, Spielvogel S, Moore FL. 1981. Luteal development, Placentation, and plasma progesterone concentration in the viviparous lizard Sceloporus jarrovi. Gen. Comp. Endocrinol. 43(1): 20–29. doi: 10.1016/0016-6480(81)90027-7.
Guraya SS. 1989. Ovarian follicles in reptiles and birds. Burggren W, Ishii S, Langer H, Neuweiler G, Randall DJ, editores. Zoophysology Volume 24. Springer-Verlag Berlin Heidelberg. p. 1–285.
Hernandez-Franyutti A, Uribe MC, Guillette LJ. 2005. Oogenesis of the viviparous matrotrophic lizard Mabuya brachyopoda. J. Morphol. 265:152–164. doi: 10.1002/jmor.10347.
Hussein MR. 2005. Apoptosis in the ovary: molecular mechanisms. Hum. Reprod. Update. 11(2): 162–178. doi: 10.1093/humupd/dmi001.
Ibargüengoytía NR. 2008. Estrategias reproductivas en reptiles. En: Vidal Maldonado MA, Labra Lillo A, Editores. Herpetología de Chile. Santiago de Chile: Science Verlag Chile. p. 391–425.
Jones RE, Swain T, Guillette LJ, Fitzgerald KT. 1982. The Comparative Anatomy of Lizard Ovaries, with Emphasis on The Number on Germinal Beds. J. Herpetol. 16(3):240–252. doi: 10.2307/1563717.
Jones RE, Summers CH. 1984. Compensatory follicular hypertrophy during the ovarian cycle of the house gecko, Hemidactylus frenatus. Anat. Rec. 209(1):59–65. doi: 10.1002/ar.1092090108.
Klosterman LL. 1987. Ultrastructural and quantitative dynamics of the granulosa of ovarian follicles of the lizard Gerrhonotus coeruleus (Family Anguidae). J. Morphol. 192(2):125–144. doi: 10.1002/jmor.1051920205.
Lozano A, Ramírez-Bautista A, Uribe MC. 2014. Oogenesis and Ovarian Histology in Two Populations of the Viviparous Lizard Sceloporus grammicus (Squamata: Prhynosomatidae) From the Central Mexican Plateau. J. Morphol. 275(8):949–960. doi: 10.1002/jmor.20275.
Makrigiannakis A, Coukos G, Christofidou-Solomidou M, Montas S, Coutifaris C. 2000. Progesterone is an autocrine/paracrine regulator of human granulosa cell survival in vitro. Ann. N. Y. Acad. Sci. 900(1):16–25. doi: 10.1111/j.1749-6632.2000.tb06212.x.
Manes ME, Noriega T, Campos Casal F, Apichela S. 2007. Ovarian changes during the reproductive cycle of the Tupinambis merianae lizard raised in a temperate environment. Cuad. Herpetol. 21(1):21–29.
Martínez-Torres M, Hernández-Caballero ME, Alvarez-Rodríguez C, Luis-Díaz J, Ortíz-López G. 2003. Luteal development and progesterone levels during pregnancy of the viviparous temperate lizard Barisia imbricata imbricata (Reptilia: Anguidae). Gen. Comp. Endocrinol. 132 55–65. doi: 10.1016/S0016-6480(02)00607-X.
Motta CM, Castriota-Scanderbeg M, Filosa S, Andreuccetti P. 1995. Role of Pyriform Cells During the Growth of Oocytes in the Lizard Podarcis sicula. J. Exp. Zool. 273: 247-256. doi: 10.1002/jez.1402730310.
Motta CM, Filosa S, Andreuccetti P. 1996. Regression of the epithelium in late previtellogenic follicle of Podarcis sicula: a case of apoptosis. J. Exp. Zool. 276(3):111–120. doi: 10.1002/(SICI)1097-0 1 0 X ( 1 9 9 6 1 0 1 5 ) 2 7 6 : 3 < 2 3 3 : : A I D -JEZ7>3.0.CO;2-O.
Motta PM, Nottola SA, Familiari G, Makabe S, Stallone T, Machiarelli G. 2002. Morphodynamics of the Follicular-luteal complex during early ovarian development and reproductive life. Int. Rev. Cytol. 223:177–288. doi: 10.1016/S0074-7696(05)23004-8
Palmer BD, Perkins MJ, Simon S, Massie K, Uribe MC. 1997. Reproductive anatomy and physiology: an ecological and evolutionary perspective. En: Ackerman L, editor. The Biology, Husbandry and Health Care of Reptiles and Amphibians. Vol 1. Biology of Reptiles. USA: TFH Publications, Inc. p. 54–87.
Ramirez-Pinilla MP. 1991. Reproductive and fat body cycle of the lizard Liolaemus wiegmanni. Amphib-Reptilia. 12(2):195–202. doi: 10.1163/156853891X00167.
Ramírez-Pinilla MP. 1995. Reproductive and fat body cycles of the oviparous lizard Liolaemus bitaeniatus (Sauria: Tropiduridae). J. Herpetol. 29(2):256–260. doi: 10.2307/1564562.
Ramírez-Bautista A, González-Romero A. 2002. Some reproductive and feeding characteristics of the viviparous Mexican lizard Sceloporus torquatus (Phrynosomatidae). The Southwestern Naturalist. 47(1):98–102. doi: 10.2307/ 3672808.
Romero de Pérez G, Ramírez-Pinilla MP, Calderón ML. 1999. Estudio preliminar de la ultraestructura de la pared del ovario y de folículos previtelogénicos y vitelogénicos tempranos de Caiman crocodilus fuscus. Rev. Acad. Colomb. Cienc. Exact. Fis. Nat. 23:453–464.
Santos HB, Thomé RG, Arantes FP, Sato Y, Bazzoli N, Rizzo E. 2008. Ovarian follicular atresia is mediated by heterophagy, autophagy and apoptosis in Prochilodus argenteus and Leporinus taeniatus (Teleostei: Characiformes). Theriogenology. 70(9):1449–1460. doi: 10.1016/j.theriogenology.2008.06.091.
Sanz-Ochotorena A, Uribe MC. 1999. Ciclo gonadal y de los cuerpos grasos de Anolis sagrei (Sauria: Iguanidae) en ciudad de La Habana, Cuba. Biología. 13: 26-30.
Sanz-Ochotorena A, Uribe MC, Coro Antich RM, Torres F. 2000. Ciclo reproductivo de Anolis porcatus (Sauria: Polychrotidae) en ciudad de La Habana, Cuba. Biología. 14(2):114-120.
Sanz-Ochotorena A, Uribe MC, Guillette LJ. 2005. Seasonal gametogenic cycles in a Cuban tropical lizards, Anolis porcatus. J. Herpetol. 39(3):443–454. doi: 10.1670/160-03A.1.
Shanbhag BA, Prasad BSK. 1993. Follicular dynamics and germinal bed activity during the annual ovarian cycle of the lizard Calotes versicolor. J. Morphol. 216(1):1–7. doi: 10.1002/jmor.1052160102.
Shulte JA, Macey JRM, Espinoza RE, Larzon A. 2000. Phylogenetic relationships in the iguanid lizard genus Liolaemus: multiple origins of viviparous reproduction and evidence for recurring Andean vicariance and dispersal. Biol. J. Linn. Soc. 69(1):75–102. doi: 10.1006/bijl.1999.0346.
Tammaro S, Simoniello P, Filosa S, Motta CM. 2006. Block of mitochondrial apoptotic pathways in lizard ovarian follicle cells as an adaptation to their nurse function. Cell. Tissue Res. 327(3):625–635. doi: 10.1007/s00441-006-0256-7.
Thomé RG, Santos HB, Arantes FP, Domingos FT, Bazzoli N, Rizzo E. 2009. Dual roles for autophagy during follicular atresia in fish ovary. Autophagy. 5(1):117–119. doi: 10.4161/auto.5.1.7302
Uribe MC, Mendéz-Omana EM, González-Quintero J, Gillette LJ. 1995. Seasonal variation in ovarian histology of the viviparous lizard Sceloporus torquatus torquatus. J. Morphol. 226(1):103–119. doi: 10.1002/jmor.1052260107.
Uribe MC, Portales GLB, Guillette LJ. 1996. Ovarian folliculogenesis in the viviparous Mexican lizard Ctenosaura pectinata. J. Morphol. 230(1):99–112. doi: 10.1002/(SICI)1097-4687(199610)230:1<99::AIDJMOR8>3.0.CO;2-I.
Uribe MC, González-Porter P, Palmer B, Guillette LJ. 1998. Cyclic histological changes of the oviductal-cloacal junction in the viviparous snake Toluca lineata. J. Morphol. 237(1):91–100. doi: 10.1002/(SICI)10974687(199808)237:2<91::AIDJMOR1>3.0.CO;2-R.
Uribe MC, Guillette LJ. 2000. Oogenesis and ovarian histology of the American alligator Alligator mississippiensis. J. Morphol. 245(3):225–240. doi: 10.1002/10974687(200009)245:3<225::AIDJMOR4>3.0.CO;2-M.
Uribe MC, Hernández-Franyutti AA, Sanz-Ochotorena A, González-Porter G. 2010. Estructura comparada de los folículos ováricos en reptiles: En: Hernández-G O, Méndez-DLC FR, Méndez-S JF, editores. Reproducción en Reptiles. México: Editorial UAEM. p. 176–201.
Zaracho VH, Ingaramo MR, Semhan RV, Etchepare EG, Acosta JL, Falcione AC, Álvarez BB. 2014. Herpetofauna de la Reserva Natural Provincial Isla Apipé Grande (Corrientes, Argentina). Cuad. Herpetol. 28:153–160.
Zaracho VH, Tedesco ME, Motte M, Yanoski A. 2017. First record of Liolaemus azarai and Hemidactylus mabouia for Ñeembucú Department, Paraguay. Cuad. Herpetol. 31(1):65–67.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
CrossRef Cited-by
1. Enely Maris da Silveira Firmiano, Clarice Machado-Santos, Amanda Oliveira Serra-Campos, Bernadete Maria de Sousa, Nadja Lima Pinheiro, Aparecida Alves do Nascimento. (2021). Histological study and immunohistochemical location of cytoskeletal proteins in the ovaries of the three species of lizards of the family Leiosauridae (Reptilia: Squamata). Tissue and Cell, 68, p.101477. https://doi.org/10.1016/j.tice.2020.101477.
2. Othman A. Aldokhi, Saleh Alwasel, Abdel Halim Harrath. (2019). Ultrastructural and histochemical study of previtellogenic oogenesis in the desert lizard Scincus mitranus (Squamata, Sauropsida). Journal of Morphology, 280(3), p.381. https://doi.org/10.1002/jmor.20950.
Dimensions
PlumX
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2017 Caldasia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















