


Published
ESTRUCTURA POBLACIONAL DE LA PALMA IRIARTEA DELTOIDEA, EN UN BOSQUE DE TIERRA FIRME DE LA AMAZONIA COLOMBIANA
Downloads
ROY GONZÁLEZ-M.
ÁNGELA PARRADO-ROSSELLI
RENÉ LÓPEZ-CAMACHO
Grupo Uso y Conservación de la Diversidad Forestal, Ingeniería Forestal, Facultad del Medio Ambiente y Recursos Naturales, Universidad Distrital Francisco José de Caldas. Avenida Circunvalar-Venado de Oro, Bogotá, Colombia. rgonzalezm@udistrital.edu.co, aparrador@udistrital.edu.co,rlopezc@udistrital.edu.co
RESUMEN
El objetivo de esta investigación fue caracterizar la densidad y la estructura poblacional de la palma bombonaIriartea deltoidea en un bosque de tierra firme de la zona sur del Parque Nacional Natural Amacayacu, en la Amazonia Colombiana. En seis parcelas de 1 ha y 90 transectos de 50 x 1 m, se censaron todos los individuos de la especie. Para cada individuo, se registró la altura del tallo, el estado morfológico de las hojas (i.e. hojas enteras, hojas divididas, forma de las pinnas), la formación de raíz fúlcrea, la altura del cono de la raíz y el estado fenológico (brácteas, inflorescencias e infrutescencias). Se encontraron 2819 individuos (376±58 ind/ha) que se agruparon en tres grandes categorías de edad significativamente diferentes (e.g. plántulas, juveniles y adultos). La gran categoría Plántulas se caracterizó por individuos con hojas no divididas, sin formación de raíces fúlcreas y comprendió el 96.6% de los individuos muestreados. Esta se subdividió en seis categorías de plántulas estadísticamente significativas dadas por cambios en la altura, número y morfología de las hojas. La gran categoría Juvenil consistió en individuos con hojas divididas en pinnas, formación de raíces fúlcreas con altura de cono radicular menor a 1 m, sin estructuras reproductivas y con 1.5% (42) de los individuos totales. Esta se subdividió en dos categorías de juveniles (I y II). La categoría Adultos incluyó todos los individuos con hojas divididas en pinnas, formación de raíces fúlcreas con altura del cono radicular mayor a 1 m y con presencia de estructuras reproductivas. Estos correspondieron al 1.9% (54) de los individuos y se subdividieron en dos categorías: Adultos I y Adultos II, dadas principalmente por diferencias significativas en la altura. La densidad de la población presentó una distribución típica de poblaciones naturales de plantas tropicales, siguiendo un modelo de Poisson en donde abundan individuos de los primeros estadios de vida (plántulas) y se van reduciendo a medida que aumenta la edad (juveniles y adultos). Dentro de las 10 categorías etarias, la de Plántulas II fue la más densa (129±38 ind/ha), mientras que Juveniles I fue la menos densa (3±2 ind/ha). La densidad de los adultos fue ligeramente mayor a la de los Juveniles. Se discute la importancia de una refinada categorización etaria frente a categorizaciones realizadas en estudios previos.
Palabras clave. Categorías etarias, densidad poblacional, Palma solitaria, Parque Nacional Natural Amacayacu, plántulas.
ABSTRACT
The goal of this research was to characterize the population structure and density of the bombona palm Iriartea deltoidea in forests of terra firme in the southern areas of the Amacayacu National Park, in the Colombian Amazon. In six 1-ha plots and 90 transects of 50 x 1 m, all individuals, from seedlings to adults, were censused. For each individual, the trunk height, leaf morphology (i.e. entire or divided leafs, pinnae form), stilt root formation and cone height, as well as phenological condition (bracts, inflorescences, infrutescences) were recorded. A total of 2819 individuals (376±58 ind/ha) were found and grouped into three broad age categories(Seedlings, Juveniles, Adults). The category Seedlings, consisting of 96.6% of the individuals, was characterized by individuals with entire leafs and stilt root absence. This category was subdivided into six sub-categories that were statistically different based on height, number and leaf morphology. The Juvenile category consisted of 1.5% of the total number of individuals, which have pinnate leaves, stilt root cone lower than 1 m, and lack of reproductive structures. This category was divided into two subcategories (Juvenile I and II). The Adult category included pinnate individuals with stilt root cone above 1 m height and with reproductive structures. It corresponded to 1.9% individuals and was subdivided into two subcategories (Adults I and II) based on significant differences in the trunk height. The population density of this palm exhibited the common pattern found in other natural populations of tropical plants, that is, following a Poisson model where individuals at early life stages (Seedlings) are very abundant, decreasing in number with age (Juveniles and Adults). Within the ten age categories and subcategories described, Seedlings II was the most abundant (129±38 ind/ha) while Juveniles I was the least (3±2 ind/ha). Adult density was slightly higher than that of Juveniles. We discuss the importance of using a more refined age categorization.
Key words. Age categories, population density, solitary palm, Amacayacu National Park, seedlings.
Recibido: 22/04/2011
Aceptado: 15/05/2012
INTRODUCCIÓN
Las palmas son reconocidas por su abundancia e importante papel que desempeñan en la composición, la estructura y la función de los bosques húmedos neotropicales (Peres 1994, Aguilar & Jiménez 2009). En la Amazonia noroccidental la familia Arecaceae es uno de elementos naturales más abundantes y dominantes de las diferentes masas boscosas, llegando a mostrar altos índices de valor de importancia (Duivenvoorden 1995, Terborgh & Andresen 1998, Duque et al. 2003, Vormisto et al. 2004, Montufar & Pintaud 2006, Cantillo 2007). Además, las palmas son una de las fuentes alimenticias más importantes para la fauna debido a su producción periódica y sincrónica de frutos, lo que las hace predecibles para las especies que de ellas dependen (Peres 1994). También, los altos contenidos nutricionales de sus frutos, las hacen elementos fundamentales en las dietas de los animales frugívoros presentes en los bosques (Smythe 1989, Peres 1994, Beck 2005, 2006).
En el mismo sentido, las palmas son un componente importante en la cultura de las comunidades locales que habitan los bosques, bien sea para usos de subsistencia o como una importante fuente de ingresos económicos (Kahn 1991, Peres 1994, Anderson & Putz 2002, López et al.2006, Byg et al. 2007). Galeano & Bernal (2010) han expuesto la importancia y la relación constante que tienen los humanos con las diferentes especies de palmas puesto que ofrecen una amplia gama de bienes tanto no elaborados (e.g. frutos, semillas, hojas), como materia prima que se emplea para la elaboración de un gran número de productos (e.g. elementos para construcción, elaboración de muebles y artesanías, producción industrial de alimentos).
Ante esta demostrada importancia, diferentes estudios han tenido que definir categorías de "edad" para cada especie (e.g. Durán & Franco 1992, Pinard 1993, Bernal 1998, Rodríguez-Buriticá et al. 2005, Arango et al. 2010) con el fin de evaluar aspectos como la demografía poblacional de las especies altamente utilizadas (e.g. Bernal 1998, Arango et al. 2010, Navarro et al. 2010), los efectos de la extracción, las dinámicas de regeneración y el estado de conservación de las poblaciones naturales (TNC 2000). Sin embargo, los mecanismos para la definición de dichas categorías de edad se han basado, por lo general, en divisiones sistemáticas (rangos o marcas de clase con amplitudes similares), sin combinar con los rasgos morfológicos de la especie, aspecto que puede generar la inclusión de individuos en una categoría a la que no pertenece o traslapes de individuos con determinados rasgos en más de una categoría definida.
La palma Iriartea deltoidea Ruiz & Pav. (1978), es una especie mencionada por ser ampliamente abundante y representativa de los bosques de tierra firme de la amazonia (Valencia et al. 2004, Vormisto et al. 2004, Macia & Svenning 2005). Es muy utilizada por diferentes comunidades humanas en la construcción de pisos, paredes y muebles. Los frutos son empleados para la alimentación de animales y el palmito y almidón obtenidos son utilizados para consumo humano. A partir de las raíces, el tallo y las semillas se elaboran elementos artesanales con fines comerciales (Bernal 1992, Acero 2000, Anderson & Putz 2002, López et al. 2006, Bernal et al. 2007, Galeano & Bernal 2010).
Dada esta importancia, algunos autores han evaluado aspectos relacionados con el estado y el manejo de sus poblaciones, el impacto de su aprovechamiento y su conservación (i.e. Pinard 1993, Pitman 2000, Anderson & Putz 2002, Anderson 2004). Sin embargo, aun es deficiente el conocimiento sobre las categorías etáreas de esta especie, por lo que estimaciones sobre la demografía, el estado de conservación de sus poblaciones y el impacto de su aprovechamiento pueden estar siendo inexactas. Por lo tanto, con el fin de refinar los criterios y las metodologías para la obtención de las categorías etarias de I. deltoidea en los bosques de tierra firme de la amazonia colombiana, el objetivo general de este artículo fue caracterizar la estructura poblacional de la especie en los bosques de tierra firme de la zona sur del Parque Nacional Natural Amacayacu. Esto se realizó a través de dos objetivos específicos. En primer lugar, se definieron las categorías etarias de la población con base en los caracteres morfológicos de la especie y rasgos estructurales. En segundo lugar, se evalúo la densidad poblacional en cada una de las categorías obtenidas y los cambios de esta a medida que aumenta la categoría etaria. Finalmente, se discuten las implicaciones de la metodología de establecimiento de categorías etarias empleada en este trabajo frente a propuestas realizadas en otros estudios para la misma especie en diferentes sectores de la amazonia.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en el Parque Nacional Natural Amacayacu, localizado en el denominado trapecio Amazónico, en el departamento del Amazonas, Colombia. Éste cuenta con una extensión de 2935 km2 y las elevaciones oscilan entre 80 y 200 msnm (Rudas 1996, Barrera et al. 2008). La zona de muestreo se ubica en el municipio de Leticia sobre el área no traslapada con los resguardos de las comunidades indígenas Tikuna (San Martín, Palmeras y Mocagua). Las coordenadas geográficas centrales de la zona de muestreo son 3°47'38.4" Sur y 70°16'8.4" Oeste. La zona presenta una temperatura media anual de 26°C, una humedad relativa promedio multianual de 86%, con un régimen de lluvias monomodal y 2836 mm de precipitación media anual. La evaporación y evapotranspiración reportan un valor medio multianual de 1127.6 y 1506.2 mm respectivamente (Rudas, 1996). Los suelos se caracterizan por presentar bajos niveles de fertilidad principalmente en la fase mineral, alto grado de acidez, baja saturación de bases y contenidos bajos de calcio, magnesio, potasio y fósforo disponibles (Martínez 1998).
El muestreo se realizó en los bosques de tierra firme de la zona sur del parque, los cuales son la unidad dominante tanto de la Amazonia colombiana como del parque (Rudas & Prieto 2005, Ruiz et al. 2007). Las familias más representativas son Annonaceae, Araceae, Arecaceae, Melastomataceae, Mimosaceae, Poaceae y Rubiaceae, mientras que los géneros más importantes según el número de especies son Inga, Psychotria, Miconia, Ficus,Anthurium, Piper y Guarea (Rudas 1996). Algunas de las especies con amplia distribución en el parque sonTapirira guianenis, Annona montana, Duguetia spixiana, Guatteria spp., Protium spp., Pseuldolmedia laevis, I. deltoidea, Virola spp., Iryanthera spp. (Rudas 1996). El territorio del PNN Amacayacu es tradicionalmente indígena, principalmente de la etnia Tikuna, y una porción considerable de la zona sur se encuentra traslapada bajo la figura de resguardo. Por lo tanto, a pesar de ser un área protegida de nivel nacional, las poblaciones de la palma han estado históricamente sometidas a ciertos niveles de extracción.
Especie en estudio
Iriartea deltoidea Ruiz & Pav. (Arecaceae), es una especie común en los bosques de tierra firme de la amazonia (Pitman 2000, Anderson & Putz 2002, Vormisto et al. 2004) que crece de forma solitaria. Su distribución geográfica va desde Nicaragua hasta el sur de América (pasando por Centroamérica, la Guyana la Amazonia, la Orinoquia y los Andes, encontrándose en Bolivia, Brasil, Colombia, Perú y Venezuela) con una distribución altitudinal que va desde los 0 hasta los 1350 m de altitud y se encuentra habitando bosques primarios, secundarios y ribereños, chagras y áreas de pastizales (Pinard 1993, López et al. 2006, Carvajal & Murillo-A 2007, Galeano & Bernal 2010).
Según López et al. (2006) y Galeano & Bernal (2010), en estado adulto presenta un tallo solitario color grisáceo-blanquecino que alcanza entre 25 y 30 m de altura, el diámetro del estípite varía entre 25 y 30 cm, con un engrosamiento (barriga) hacia la mitad del tallo que puede llegar a presentar hasta 100 cm de diámetro. Las raíces son fúlcreas con diámetros cercanos a los 5 cm que forman una especie de cono denso con altura de hasta 3.5 m; la copa o corona está formada por 5-7 hojas, horizontales o levemente erectas con un raquis de 3- 4 m de largo y 20-30 pinnas por cada lado divididas en 2-20 partes. Las inflorescencias pueden alcanzar hasta 1.5 m de largo, con el pedúnculo de 30 cm de largo y 5 cm de diámetro como máximo; los frutos tienen forma globosa o esférica, color café amarillento al madurar y la cáscara es lisa, brillante y quebradiza con diámetros entre 2- 3 cm; las semillas color café, poseen diámetros que alcanzan los 2 cm y se encuentran cubiertas de fibras blanquecinas (Link & Stevenson 2004, López et al. 2006, Galeano & Bernal 2010).
Toma de datos
El muestreo se realizó durante los meses de julio y agosto de 2009. Se instalaron 6 parcelas de 1 ha separadas desde 250 m hasta 3 km entre sí. Adicionalmente se instalaron 90 transectos de 50 x 1 m ( 0.005 ha) cada uno, para un área total de muestreo de 6.45 ha. En cada sitio se censaron todos los individuos existentes de I. deltoidea. Para cada individuo se midió la altura del tallo, adaptando la metodología propuesta por Vallejo et al.(2005), donde la altura de individuos ≥ 2 m fue estimada con base en un tubo de aluminio estandarizado a 1.5 m y observador por encima del nivel del suelo, los individuos con alturas < 2 m y ≥ 30 cm se midieron con flexometro de precisión al centímetro y los individuos con alturas < 30 cm se midieron con escala métrica de precisión al milímetro, removiendo la hojarasca existente alrededor del tallo. Adicionalmente, se registró el estado morfológico de las hojas (hojas enteras, hojas divididas, forma de las pinnas), la formación de la raíz fúlcrea, la altura del cono de la raíz y el estado fenológico (brácteas, inflorescencias, infrutescencias (con base en lo sugerido por Durán & Franco 1992, Bernal 1998, Rodríguez-Buriticá et al. 2005). Se colectaron muestras de material vegetal de diferentes individuos las cuales se procesaron en el Herbario Forestal Gilberto Emilio Mahecha de la Universidad Distrital Francisco José de Caldas (UDBC).
Análisis de datos
Posterior a la toma de datos en campo, mediante diagramas de Box & Whiskers (Quinn & Keough 2002), los individuos se agruparon en categorías y subcategorías con base en rasgos morfológicos (i.e. forma de la hoja, división y disposición de las pinnas, formación de raíces fulcreas y presencia de estructuras reproductivas) y rasgos estructurales (i.e. altura del tallo o estípite, altura del cono radicular y número de hojas). Con el fin de aceptar o no la división de estas categorías se construyeron intervalos de confianza y se realizaron pruebas de significancia a partir del estadístico t-Student (Z a un nivel de confianza del 95%; Muria & Gil 1998, Álvarez 2007). Paralelamente, los individuos de este muestreo se agruparon en las clases o categorías propuestas por Pinard (1993) y Anderson & Putz (2002), basadas en rangos de altura con intervalos de similar amplitud. Así, se elaboraron modelos de distribución Poisson para la aproximación teórica de la distribución poblacional con base en las abundancias registradas para cada una de las categorías y las propuestas de clasificación. La distribuciónPoisson ha sido establecida como una distribución de probabilidad discreta adecuada para interpretar la frecuencia de organismos en un área determinada (Quinn & Keough 2002); así mismo, considera la probabilidad de que los individuos (plantas) se distribuyan al azar en un medio (Gómez 1999). Este procedimiento permite ajustar la información compilada en campo para la especie en estudio teniendo en cuenta que los valores usados son discretos, puntuales y medidos en un espacio definido "área" (Steel & Torrie 1988, Gómez 1999, Quinn & Keough 2002). De acuerdo con Ledesma (2009), el estadístico lambda (.), construido mediante los valores observados en campo, se compara con las expectativas teóricas arrojadas por el modelo (Ecuación 1).
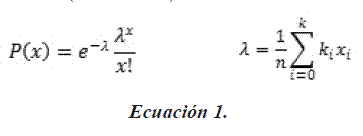
De donde P(x) corresponde a la probabilidad del número de individuos esperados para determinada categoría; x es el número de individuos en una marca de clase k. Esta marca de clase en cada categoría va en orden ascendente (e.g. planta-juvenil-adulto) y n corresponde al número total de individuos. La variación entre los valores esperados frente a los valores observados generados por cada modelo, indican cambios significativos en las tendencias de distribución de la población y se evidencian con el aumento o la reducción en los valores de ... Esta variación es corroborada mediante el estadístico para la bondad de ajuste entre curvas (Chi cuadrado X2), a un nivel de confianza del 99% (Sokal & Rohlf 1998), el cual se considera un criterio de prueba apropiado para evaluar la bondad de ajuste de estas curvas (Steel & Torrie 1988).
RESULTADOS
Grandes categorías etarias
Se encontraron 2819 individuos de I. deltoidea en 6.45 ha (376 ± 58 ind/ha) del bosque de tierra firme de la zona sur del PNN Amacayacu, agrupados en tres grandes categorías significativamente diferentes en términos de altura y los caracteres morfológicos definidos para la categorización (Figura 1).

La gran categoría plántulas comprendió todos los individuos con hojas no divididas en pinnas y sin formación de raíces fúlcreas. Esta reunió el 96.6% (2723 de los individuos muestreados. Los juveniles correspondieron a todos los individuos con hojas divididas en pinnas, formación de raíces fúlcreas con altura de cono radicular menor a 1 m y sin estructuras reproductivas. Estos representaron el 1.5% (42 individuos) de lo encontrado, mientras que los adultos el 1.9% (54) individuos (Figura 1). Estos últimos fueron todos los individuos que presentaban hojas divididas en pinnas, formación de raíces fúlcreas con altura del cono radicular mayor a 1 m y con presencia de estructuras reproductivas.
Categorías de plántulas
Las seis categorías de Plántulas obtenidas se discriminaron principalmente por diferencias significativas en la altura y el número de hojas (Tabla 1, Figura 2a).

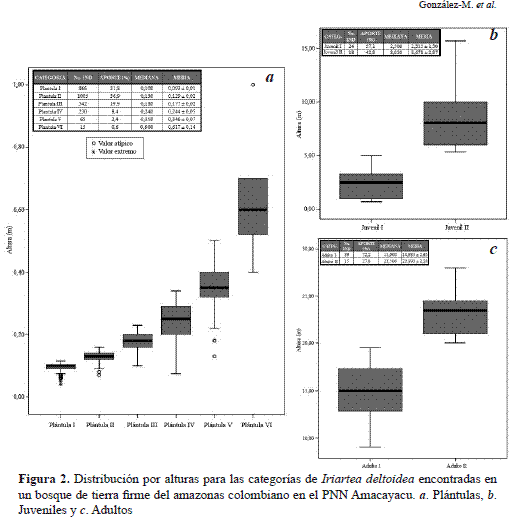
Así, las Plántulas I son individuos con altura promedio de 9.3 cm con una sola hoja no dividida. Las Plántulas II se caracterizan por presentar una altura promedio de 12.9 cm y pueden presentar una o dos hojas no divididas. Sin embargo, cuando presentan una sola hoja, la altura mínima de los individuos es de 12.0 cm (Figura 2a, Figura 3,Tabla 2).
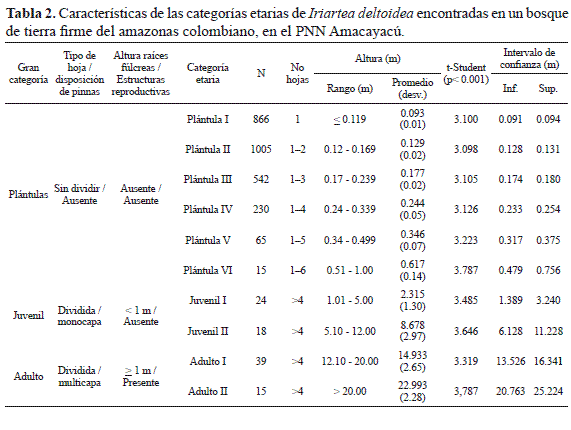
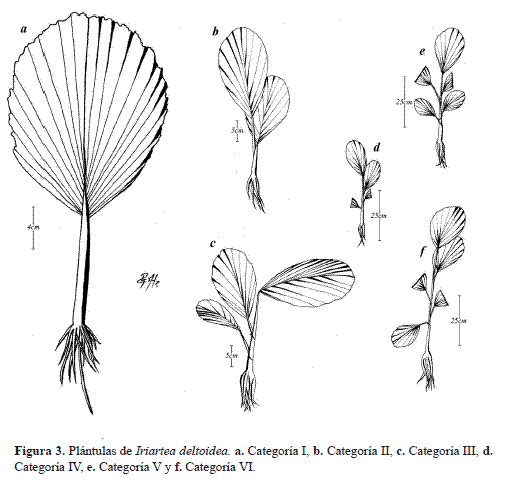
Las Plántulas III se caracterizan por presentar alturas promedio de 17.7 cm con un máximo de tres hojas no divididas. Los individuos de esta categoría con una o dos hojas tienen alturas superiores a 17.0 cm de altura. La categoría de Plántula IV presentó individuos con una altura promedio de 24.4 cm y 4 hojas no divididas como máximo y el límite inferior para ingresar a la categoría es de 24.0 cm. Las plántulas V presentan una altura promedio de 34.6 cm con un desarrollo foliar de 5 hojas no divididas como máximo y nunca una altura menor a 34.0 cm (Figura 2a, Figura 3, Tabla 2). Las plántulas VI presentan individuos con un máximo de 6 hojas no divididas, altura promedio de 61,7 cm y altura mínima de 51.0 cm.
Categorías de juveniles
Aunque las dos categorías de juveniles de I. deltoidea (Juvenil I y Juvenil II)encontradas, presentan individuos con hojas divididas en pinnas (obovadas a triangulares, dispuestas en monocapa), raíces fúlcreas inferiores a 1 m y una reducida formación de la corona o capitel, estas se reconocen por diferencias significativas en la altura (Zt(22, p<0.05)= 2.074; Zc=8.505; p<0.0001) y por qué no evidencian estructuras reproductivas (Tabla 2, Figura 2b, Figura 4). Los individuos pertenecientes a la categoría Juvenil I presentan una altura promedio de 2.3 m y llegan a alcanzar los 5 m cuando se evidencia mejor el desarrollo radicular (Figura 4). Los Juveniles II son individuos con una altura promedio de 8.7 m y llegan a alcanzar los 12 m; el capitel se encuentra más desarrollado al igual que el cono radicular, pero no presenta una altura superior a 1 m desde el suelo (Figura 2b, Figura 4,Tabla 2).

Categorías de adultos
Al igual que la gran categoría juvenil, los adultos presentaron dos categorías etarias (Adulto I y II). Los individuos de estas categorías se caracterizan por presentar hojas mayores a 2.0 m de longitud divididas en pinnas alargadas, irregulares y dispuestas en multicapa, que estructuran claramente el capitel o corona de la palma (Figura 5).

El cono de la raíz fúlcrea tiene una altura igual o mayor a 1 m y se encuentran evidencias de estructuras reproductivas (brácteas, inflorescencias, infrutescencias). Adicionalmente, se observa un claro engrosamiento del estípite (barriga) a una altura inferior a la localización de las estructuras reproductivas (Figura 5). La categoría Adulto I, presentan una altura promedio de 14.9 m y alcanzan un máximo de 20 m (Tabla 2, Figura 2c). En contraste, los Adultos II presentan una significativa diferencia en los rangos de altura promedio siendo esta última de 23 m (Zt(29; p<0.05)= 2.045; Zc=11.107; p<0.0001; Tabla 2, Figura 2c).
Densidad y modelo de distribución de la población
En promedio se encontraron 376±58 individuos por hectárea. La población presenta una distribución típica de poblaciones naturales de plantas tropicales, en donde abundan individuos de los primeros estadios de vida (plántulas) y se van reduciendo a medida que aumenta la edad (Juveniles y Adultos; Figura 6a). Sin embargo, se observó que los juveniles presentaron una menor densidad que los adultos.
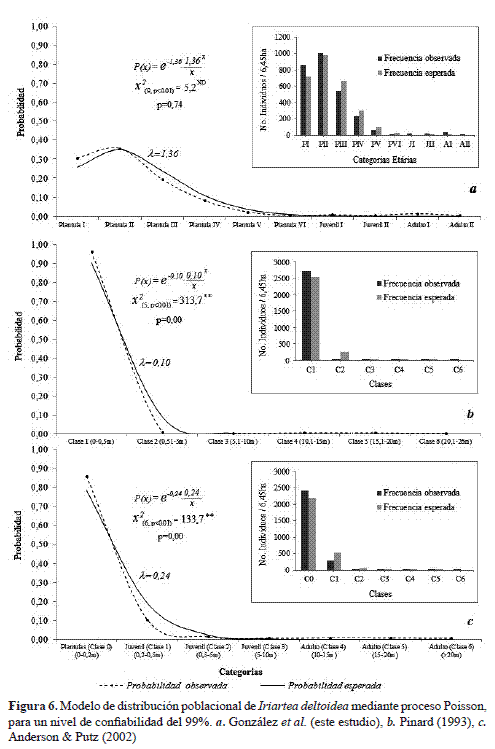
Respecto a cada una de las categorías, la gran categoría plántulas presentó los mayores valores de individuos por ha (360±55), siendo las plántulas II la categoría más densa con 129±38 ind/ha, mientras que las categorías de plántulas V y VI fueron las de más baja densidad de individuos por hectárea (10±5 y 3±1 ind/ha, respectivamente; Figura 6a). En segundo lugar, la densidad de los Juveniles II fue menor a la de Juveniles I (3±2 ind/ha, 4±2 ind/ha, respectivamente). En tercer lugar, aunque también baja (9±3 ind/ha) la densidad de los adultos fue ligeramente mayor a la de los Juveniles, en donde los Adultos I presentaron 7±4 ind/ha, mientras que los Adultos II presentaron 3±2 ind/ha (Figura 6a).
A pesar de las menores densidades de juveniles respecto de adultos, el modelo probabilístico de distribución dePoisson muestra que la probabilidad de la distribución es similar frente a los valores teóricos del modelo con un =1.36 (Figura 6a). Al realizar la prueba de bondad de ajuste (X2), no se encontraron diferencias significativas entre la distribución probabilística observada y la esperada (X2=5.20 ≤ X2(6, p<0.01)= 21.7; p=0.74) para la distribución por categorías etarias sobre 6.45 ha, por lo cual se corrobora la tendencia de que la población de I. deltoidea sigue una distribución Poisson.
De manera contrastante al categorizar la población muestreada en este estudio con base en las clases definidas por Pinard (1993; Figura 6b) y Anderson & Putz (2002; Figura 6c) se encontraron valores de <1 y diferencias altamente significativas entre los valores observados y los valores esperados (p< 0,000) de las curvas probabilísticas que exhiben la estructura poblacional de la especie bajo el modelo de Poisson, aspecto que corrobora una vez más las categorías obtenidas para I. deltoidea en el bosque de tierra firme estudiado en la amazonia colombiana.
DISCUSIÓN
Estructura etaria y morfología de I. deltoidea
En este estudio se definieron diez categorías etarias de I. deltoidea, diferenciables a partir de caracteres morfológicos como la división de las hojas en pinnas, la formación de raíces fúlcreas, la presencia o ausencia de estructuras reproductivas y variables estructurales como la altura, el número de hojas y la altura del cono radicular. Esto coincide con otras investigaciones sobre diferentes especies de palmas, en las que los caracteres morfológicos han sido elementos fundamentales para la definición de grupos de edad. Por ejemplo, Rodríguez-Buriticá et al. 2005 definieron para Geonoma orbignyana once categorías de edad basados en el número de venas y desarrollo del tallo. Igualmente, Bernal (1998) se apoyó en el desarrollo del tallo y en el número de pinnas para realizar la clasificación etaria de Phytelephas seemannii, estableciendo cinco categorías etarias, mientras que Navarro et al. 2010 definieron las categorías para Lepidocaryum tenue a partir de la relación entre el desarrollo del tallo, la formación de anillo y el número de venas. A pesar de lo anterior, dada la gran variabilidad morfológica que presenta I. deltoidea principalmente en la forma en las hojas a medida que aumenta la altura (Rich et al.1995, Terborgh & Davenport 2001), se resalta necesidad de incluir otras variables cualitativas y estructurales para una depuración de las categorías estarías obtenidas en el presente estudio.
La clasificación etaria de la población de I. deltoidea realizada aquí, a partir de parámetros estadísticos con las variables mencionadas y corroborada mediante un modelo de distribución Poisson, presentó buen un ajuste de los datos observados frente a las expectativas teóricas generadas por el modelo. De esta forma, se resalta la ventaja de conformación de categorías después de la toma de datos en campo (con base en caracteres morfológicos y estructurales), mientras que se muestra que la división de las clases de edad en intervalos similares de altura, como lo propuesto por Pinard (1993) y Anderson & Putz (2002), presenta inconsistencias al aplicarse a la estructura de la población de los bosques de tierra firme de amazonia colombiana. Esto se debe a que individuos con similares características morfológicas pueden quedar agrupados en diferentes clases de alturas o viceversa, por lo que se aumenta la incertidumbre sobre la real categoría de edad de un individuo particular, tal y como lo encontraron Terborgh & Davenport (2001). Ellos muestran que pese a la fuerte relación entre el incremento en altura y la variación en la forma de las hojas, existe incertidumbre en el rango de 10- 20 m donde la especie presenta traslapes en los valores de altura y la disposición multicapa de las pinnas en la hoja dada la alta flexibilidad morfológica de la especie.
No obstante lo anterior, los resultados de este estudio coinciden con los valores de altura y rasgos morfológicos de las hojas obtenidos por Rich et al. (1995) en Costa Rica. Ellos mencionan que los individuos juveniles poco desarrollados de I. deltoidea en Costa Rica presentan alturas promedio de 1.47± 0.9 m y 3.43± 0.19 m y poseen hojas divididas en pinas dispuestas en una sola capa. También establecen que individuos medianamente desarrollados presentan una altura promedio de 8.10± 0.4 m con hojas divididas en pinas dispuestas en una leve multicapa y que los individuos desarrollados o adultos presentan alturas promedio de 15.17± 0.53 m y 25.7 m con hojas divididas en pinnas y dispuestas claramente en multicapa. Adicionalmente, el punto de división entre juveniles (poco desarrollados según el autor) y adultos (bien desarrollados), basados en los rasgos de las hojas, se genera a una altura de 12 m, elemento que confirma nuevamente estas categorías encontradas para la amazonia colombiana.
En este trabajo, el incorporar los valores de altura de las raíces fúlcreas ayuda a una más clara definición entre adultos y juveniles lo cual no había sido considerado anteriormente. Además, coincide con lo afirmado por Avaloset al. (2005), acerca de la fuerte correlación entre el cono radicular y la altura del tallo de la palma, en donde los individuos con mayor tamaño presentan conos más grandes. Adicionalmente, el tamaño del área de muestreo y la utilización de múltiples variables sintetizadas en diagramas de Box & Whiskers, complementados con la obtención de intervalos de confianza fortalece las categorías definidas, hizo posible identificar si las variables de decisión con la cual se generaron las categorías era apropiada, al evaluar si se presentaba o no traslape de datos entre los grupos (González et al. 2007).
Densidad poblacional de I. deltoidea
Aunque I. deltoidea es una especie dominante en los bosques tropicales bajos de Latinoamérica (Suatunce et al.2003, Valencia et al. 2004, Vormisto et al. 2004, Macia & Svenning 2005, Paniagua-Zambrana 2005, Silman et al.2005, Cabrera & Wallace 2007, Barreto et al. 2010), se encuentran importantes variaciones en el número de individuos por unidad de área entre sitios (Cordero et al. 2003). Así, el número de individuos encontrados (376 ± 58 ind/ha) presenta un valor intermedio frente a los reportes realizados por Vormisto et al. (2004) quienes mencionan que en la región de Yasuní (Ecuador) sobre 2.5 ha se hallan 1999 individuos (780 ind/ha aproximadamente), mientras que en la región de Iquitos-Pebas (Perú) se encuentran 793 ind en 2.75 ha (288 ind/ha aproximadamente). Adicionalmente, aunque existen otros estudios en la amazonia colombiana, estos incluyen solamente individuos de DAP ≥10cm, por lo que los resultados de densidad no son comparables con los presentados en este estudio (Peña et al. (2010). Por lo tanto, este trabajo aporta nueva información sobre la densidad de esta palma desde plántulas hasta adultos en bosques de tierra firme de la amazonia colombiana.
Al evaluar los valores de densidad por categorías, se encontró que a nivel de plántulas los valores son superiores a los reportados por Kahn & Mejía (1991) en bosques de quebrada en la amazonia peruana (153 ind / 1.6 ha, individuos entre 0- 1 m de altura). En contraste, la densidad de juveniles, según estos autores, es mayor (individuos entre 1-10m, 104 ind / 1.6 ha, 64 ind/ha aproximadamente). Respecto a los adultos, Barreto et al.(2010) y Peña et al. (2010), reportan 104 individuos con DAP ≥ 10 cm en 5 ha ubicadas en los mismos bosques en donde se realizó el presente trabajo, mientras que nosotros presentamos 96 individuos en las categorías juveniles y adultos (de los cuales el 89% son equiparables al rango de DAP ≥ 10cm) en 6 ha discontinuas, por lo que se evidencia una similitud en los diferentes trabajos, y contrastan con lo encontrado con Kahn & Mejía (1991) y por Vormisto et al. (2004) para la amazonia peruana.
Implicaciones para el manejo
Una adecuada categorización etaria realizada de forma estática a partir de variables morfológicas y estructurales permite caracterizar mejor el estado de las poblaciones de especies con alta demanda. Además, permite una mejor caracterización del estado de los ecosistemas pues la estructura etaria ha sido sugerida como un indicador de perturbación de los ecosistemas (TNC 2000, Onaindia et al. 2004). Adicionalmente, la metodología expuesta y sustentada en un modelo de distribución Poisson, puede ser utilizada para otras palmas de porte similar (e.g.Socratea exorrhiza, Wettinia spp.).
Por otro lado, a partir de modelo de distribución Poisson se pudo establecer que tan sólo un 3% de la población deI. deltoidea pasa a categorías superiores (i.e. juveniles y adultos), lo que sugiere que la palma tiene una estrategia de crecimiento "r", lo cual coincide en diferentes estudios realizados sobre otras palmas, en las que se presenta una gran producción de nuevos individuos con valores muy altos en mortalidad de estas nuevas generaciones (Durán & Franco 1992, Bernal 1998, Conte et al. 2003, Rodríguez-Buritica et al. 2005, Arango et al.2010). Por lo tanto, teniendo en cuenta que la I. deltoidea es una palma ampliamente utilizada en la Amazonia colombiana (Anderson & Putz 2002, López et al. 2006, Bernal et al. 2007, Galeano & Bernal 2010) y que los juveniles I y II fueron los estadios que menores densidades presentaron, es importante comenzar a plantear estrategias de manejo de sus poblaciones naturales que eviten la marcada disminución de juveniles y adultos y que incidan sobre el 97% de plántulas de las categorías I-VI que no lograron pasar a estadios superiores.
CONCLUSIONES
Teniendo en cuenta que I. deltoidea es una especie con amplia variabilidad en la morfología de las hojas, con el fin establecer una adecuada división entre categorías de un grupo poblacional es necesario incluir variables adicionales a la altura del tronco como la altura del cono radicular, el número de hojas y el número de pinas.
A partir de un modelo de distribución Poisson, se logra una refinada agrupación de la población en categorías etarias con base en rasgos estructurales y morfológicos, lo cual provee datos robustos sobre la estructura poblacional de alto valor de uso en los bosques de tierra firme de la amazonia colombiana.
Dada las altas densidades de plántulas de la especie en relación a otros estadios como juveniles y adultos, se sugiere emplear el rescate de plántulas como estrategia de propagación bien sea con fines comerciales o de restauración y prestar especial atención a los estadios juveniles que presentaron una menor densidad respecto a las otras categorías etarias encontradas.
Este estudio provee nueva información sobre los valores de densidad de I. deltoidea en bosques de tierra firme de la amazonia, en especial a nivel de plántulas donde los reportes son nulos para el caso de la región sur del Amazonas colombiano.
AGRADECIMIENTOS
Agradecemos al Centro de investigaciones y Desarrollo Científico de la Universidad Distrital Francisco José de Caldas y al Parque Nacional Natural Amacayacú por el apoyo financiero y logístico al proyecto marco de donde proviene esta investigación. Muchas gracias a los habitantes de la comunidad Tikuna en Mocagua (Leticia), Bartolomé Moran y Casimiro del Águila. A los funcionarios del PNN Amacayacu Alexander Puentes, Alberto Parente, Estela Chota. Los herbarios Amazónico (COAH) y Forestal (UDBC) facilitaron sus instalaciones para la caracterización preliminar de la especie y la identificación de las muestras botánicas. Dos evaluadores anónimos ayudaron a mejorar versiones previas de este manuscrito.
LITERATURA CITADA
1. ACERO, D. 2000. Arboles, Gentes y Costumbres. Universidad Distrital Francisco José de Caldas. Plaza & Janes Editores Colombia S.A. Bogotá, D.C. Pg. 387.
2. AGUILAR, V. & F. JIMÉNEZ. 2009. Diversidad y distribución de palmas (Arecaceae) en tres fragmentos de bosque muy húmedo en Costa Rica. Biología Tropical 1: 83-92.
3. ÁLVAREZ, R. 2007. Estadística aplicada a las ciencias de la salud. Ediciones Díaz de Santos. Madrid. Pg. 996.
4. ANDERSON, P.J., PUTZ, F.E., 2002. Harvesting and conservation: are both possible for the palm, Iriartea deltoidea? Forest Ecology and Management 170: 271–283.
5. ANDERSON, P.J. 2004. The Social Context for Harvesting Iriartea deltoidea (Arecaceae). Economic Botany 58:410-419.
6. ARANGO, D., A. DUQUE & E. MUÑOZ. 2010. Dinámica poblacional de la palma Euterpe oleracea (Arecaceae) en bosques inundables del Chocó, Pacífico colombiano. Revista de Biología Tropical 58: 465-481.
7. BARRERA, V., J. ZAMBRANO & P. STEVENSON. 2008. Diversity of regenerating plants and seed dispersal in two canopy trees from Colombian Amazon forests with different hunting pressure. Revista de Biología Tropical 56: 1531-1542.
8. BARRETO, J.S. A.J. DUQUE, D. CÁRDENAS & F. MORENO. 2010. Variación florística de especies arbóreas a escala local en un bosque de tierra firme en la Amazonia colombiana. Acta Amazonica 40(1): 179-188.
9. BECK, H. 2005. Seed predation and dispersal by peccaries throughout the Neotropics and its consequences: a review and synthesis. En Forget, P.-M., Lambert, J. E., Hulme, P. E. & VanderWall, S.B. (eds.). Seed fate: predation dispersal and seedling establishment. CABI Publishing,Wallingford. Págs 77–115.
10. BECK, H. 2006. A review of peccary-palm interactions and their ecological ramifications across the Neotropics. Journal of Mammalogy 87:519–530.
11. BERNAL, R. 1992. Colombian palm products. En: M. Plotkin & L. Famolare (eds.). Sustainable harvest and marketing of rain forest products. Island, Washington, Columbia, EEUU. Págs. 158-172.
12. BERNAL, R. 1998. Demography of the vegetable ivory palm Phytelephas seemannii in Colombia, and the impact of seed harvesting. Journal of Applied Ecology 35: 64-74.
13. BERNAL, R., D. MARMOLEJO & M.F. MONTES. 2007. Eastern Tukanoan names of the palm Iriartea deltoidea: evidence of its possible preagricultural use as a starch source. Journal of Ethnobiology 27: 174-181.
14. BYG, A., J. VORMISTO & H. BALSLEV. 2007. Influence of diversity and road access on palm extraction at landscape scale in SE Ecuador. Biodiversity and Conservation 16: 631-642.
15. CABRERA, W.H. & R. WALLACE. 2007. Densidad y distribución espacial de palmeras arborescentes en un bosque preandino-amazónico de Bolivia. Ecología en Bolivia 42(2): 121-135.
16. CANTILLO, E. 2007. La Estructura de la Vegetación en Colombia: Una Síntesis Preliminar. Tesis Doctoral. Universidad Nacional de Colombia. Bogotá, D.C.
17. CARVAJAL, L. & J. MURILLO-A. 2007. Análisis Florístico y Fitogeográfico del sector nororiental de la sierra de la Macarena, Colombia. Universidad Distrital Francisco José de Caldas. Bogota, D.C. Pg. 214.
18. CASTAÑEDA, E. 2009. Parámetros demográficos de Dioon spinulosum Dyer & Eichler (Zamiaceae), en San Miguel Soyaltepec, Oaxaca. Instituto de Ecologia A.C. Veracruz. Pg. 129.
19. CONTE, R., R.O. NODARI, R. VENCOVSKY & M. SEDREZ DOS REIS. 2003. Genetic diversity and recruitment of the tropical palm, Euterpe edulis Mart. in a natural population from the Brazilian Atlantic Forest. Heredity 91: 401–406.
20. CORDERO, J., F. MESÉN, M. MONTERO, J. STEWART, D. BOSHIER, J. CHAMBERLAIN, T. PENNINGTON, M. HANDS, C. HUGHES & G. DETLEFSEN. 2003. Descripciones de especies de árboles nativos de América Central: Iriartea deltoidea (Ruiz & Pavón). Oxford Forestry Institute - Centro Agronómico Tropical de Investigación y Enseñanza. Turrialba. Págs. 617-620.
21. CORPOAMAZONIA. 2009. Plan de acción ajustado Corpoamazonia 2007-2011. Corporación para el desarrollo sostenible del sur de la Amazonia. Mocoa. Pg. 75.
22. DUIVENVOORDEN, J.F. 1995. Tree species composition and rain forest-environment relationships in the middle Caquetá area, Colombia, NW Amazonia. Plant Ecology 120: 91-113.
23. DUQUE A.J, D. CÁRDENAS & N. RODRÍGUEZ. 2003. Dominancia florística y variabilidad estructural en bosques de tierra firme en el noroccidente de la Amazonia Colombiana. Caldasia 25: 139-152.
24. DURÁN, R. & R. FRANCO. 1992. Estudio demográfico de Pseudophoenix sargentii. Bulletin de l'Institut Français d'Études Andines 21: 609-621.
25. IDEAM. 2009. Informe anual sobre el estado del medio ambiente y los recursos naturales renovables de Colombia: Bosques 2009. –Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia-. Bogotá, D.C. Pg. 179.
26. KAHN, F. & K. MEJÍA. 1991. Las comunidades de palmeras en los ecosistemas forestales inundables de la amazonia peruana. Folia Amazonica IIAP 3: 47-58.
27. GALEANO, G. & R. BERNAL. 2010. Palmas de Colombia, guía de campo. Universidad Nacional de Colombia, Facultad de Ciencias Naturales. Instituto de Ciencias Naturales. Bogotá, D.C. Págs. 162-166.
28. GÓMEZ, H. 1999. Algunos métodos estadísticos para el estudio de poblaciones de organismo. Universidad Nacional de Colombia, Falcultad de Ciencias Agropecuarias. Medellín. Pg. 216.
29. GONZÁLEZ, M.D., A. GONZÁLEZ & G. PÉREZ. 2007. Análisis Exploratorio de Datos. Universidad de Huelva, Facultad de Ciencias Empresariales. Huelva.
30. KAHN, F. 1991. Palms as key swamp forest resources in Amazonia. Forest Ecology and Management 38: 133-142.
31. LEDESMA, A. 2009. Una aplicación del proceso de Poisson en la enseñanza y aprendizaje de la estadística. Jornadas de Enseñanza e Investigación Educativa en el campo de las Ciencias Exactas y Naturales. Actas 2: 62-68.
32. LINK, A. & P. STEVENSON. 2004. Fruit dispersal syndromes in animal disseminated plants at Tinigua National Park, Colombia. Revista Chilena de Historia Natural 77: 319-334.
33. LÓPEZ, R., J. NAVARRO, M. MONTERO, K. AMAYA, M. RODRÍGUEZ & A. POLANIA. 2006. Manual de identificación de especies no maderables del corregimiento de Tarapacá. Instituto Amazónico de Investigaciones Científicas -Sinchi- (editor). Bogotá, D.C. Pg. 218.
34. MACIA, M.J. & J.C. SVENNING. 2005. Oligarchic dominance in western Amazonian plant communities. Journal of Tropical Ecology 21: 613-626.
35. MARTÍNEZ, L.J. 1998. Suelos de la Amazonía. Serie Escuela y Amazonía. Bogotá, D.C. Págs. 62-68.
36. MONTUFAR, R. & J. PINTAUD. 2006. Variation in species composition, abundance and micro habitat preferences among western Amazonian terra firme palm communities. Botanical Journal of the Linnean Society 151: 127–140.
37. MURIA, J. & R. GIL. 1998. Preparación, tabulación y análisis de encuentas para directivos. Esic Editorial. Madrid. Pg. 167.
38. NAVARRO, J.A., G. GALEANO & R. BERNAL. 2011. Impact of leaf harvest on populations of Lepidocaryum tenue, an Amazonian understory palm used for thatching. Tropical Conservation Science 4(1): 25-38
39. ONAINDIA, M., I. DOMINGUEZ, I. ALBIZU, C. GARBISU, & I. AMEZAGA. 2004. Vegetation diversity and vertical structure as indicators of forest disturbance. Forest Ecology and Management 195: 341–354.
40. PANIAGUA-ZAMBRANA, N.Y. 2005. Diversidad, densidad, distribución y uso de las palmas en la región del Madidi, noreste del departamento de La Paz (Bolivia). Ecologia en Bolivia 40: 265-288.
41. PERES, C. 1994. Composition, density, and fruiting phenology of arborescent palms in an Amazonian terra firme forest. Biotropica 26: 285-294.
42. PINARD, M. 1993. Impacts of stem harvesting on populations of Iriartea deltoidea (Palmae) in an extractive reserve in Acre, Brazil. Biotropica 25: 2-14.
43. PITMAN, N. 2000. A large-scale inventory of two Amazonian tree communities. PhD thesis. DukeUniversity, Department of Botany. Pg. 220.
44. QUINN, G.P. & M.J. KEOUGH. 2002. Experimental design and data analysis for biologists. Cambridge University Press. Cambridge. Pg. 509.
45. RICH, P.M., M. HOLBROOK & A.N. LUTTINGER. 1995. Leaf development and crown geometry of two iriarteoid palms. American Journal of Botany 82(3): 328-336.
46. RODRÍGUEZ-BURITICA, S., M. ORJUELA & G. GALEANO. 2005. Demography and life history of Geonoma orbignyana: An understory palm used as foliage in Colombia. Forest Ecology and Management 211: 329-340.
47. RUDAS, A. 1996. Estudio florístico y de la vegetación del Parque Nacional Natural Amacayacu. Tesis de Maestría. Universidad Nacional de Colombia. Bogotá, D.C.
48. RUDAS, A. & A. PRIETO. 2005. Flórula del Parque Nacional Natural Amacayacu. Monographs in Systematic Botany. Missouri Botanical Garden. St. Louis. Pg. 68.
49. RUIZ S. L., SÁNCHEZ E., TABARES E., PRIETO A., ARIAS J. C, GÓMEZ R., CASTELLANOS D., GARCÍA P. & L. RODRÍGUEZ. 2007. Diversidad biológica y cultural del sur de la Amazonia colombiana - Diagnóstico. Corpoamazonia, Instituto Humboldt, Instituto Sinchi, UAESPNN, Bogotá. D.C. pp 636.
50. SILMAN, M.R., A. ARAUJO-MURAKAMI, D.H. URREGO, M.B. BUSH & H. PARIAMO. 2005. Estructura de las comunidades de árboles en el límite sur de la Amazonía occidental: Manu y Madidi. Ecologia en Bolivia 40(3): 443-452.
51. SMYTHE, N. 1989. Seed Survival in the Palm Astrocaryum standleyanum: evidence for dependence upon its seed dispersers. Biotropica 21: 50-56.
52. SOKAL, R.R. & F.J. ROHLF. 1998. Biometry: The principles and practice of statistics in biological research. W.H. Freeman and Company. Pg. 776.
53. STEEL, R.G. & J.H. TORRIE. 1988. Bioestadistica: principios y procedimientos. Mc Graw-Hill. México D.F. Pg. 622
54. SUATUNCE, P., E. SOMARRIBA, C. HALEY & B. FINEGAN. 2003. Composición florística y estructura de bosques y cacaotales en los Territorios Indígenas de Talamanca, Costa Rica. Agroforesteria en las Américas 10(37-38): 31-35.
55. TERBORGH, J. & E. ANDRESEN. 1998. The Composition of Amazonian Forests: Patterns at Local and Regional Scales. Journal of Tropical Ecology 14: 645-664.
56. TERBORGH, J. & L. DAVENPORT. 2001. Endogenous and exogenous control of leaf morphology in Iriartea deltoidea (Palmae). Journal of Tropical Ecology 17: 695-703
57. TNC - THE NATURE CONSERVANCY. 2000. The Five-S Framework for Site Conservation: A Practitioner's Handbook for Site Conservation Planning and Measuring Success. Segunda edición. Arlington.
58. VALENCIA R., R.B. FOSTER, G. VILLA, R. CONDIT, J.C. SVENNING, C. HERNÁNDEZ, K. ROMOLEROUX, E. LOSOS, E. MAGARD & H. BALSLEV. 2004. Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador. Journal of Ecology 92: 214-229.
59. VALLEJO M., A. LONDOÑO, R. LÓPEZ, G. GALEANO, E. ÁLVAREZ & W. DEVIA. 2005. Establecimiento de parcelas permanentes en bosques de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá D.C. 309 pp.
60. VORMISTO, J., J. SVENNING, P. HALL & H. BALSLEV. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. Journal of Ecology 92: 577–588.
How to Cite
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Download Citation
Article abstract page views
Downloads
License
Copyright (c) 2012 Caldasia

This work is licensed under a Creative Commons Attribution 4.0 International License.
Authors who publish with this journal agree to the following terms:
- Authors retain copyright and grant the journal right of first publication with the work simultaneously licensed under a Creative Commons Attribution License that allows others to share the work with an acknowledgement of the work's authorship and initial publication in this journal.
- Authors are able to enter into separate, additional contractual arrangements for the non-exclusive distribution of the journal's published version of the work (e.g., post it to an institutional repository or publish it in a book), with an acknowledgement of its initial publication in this journal.
- Authors are permitted and encouraged to post their work online (e.g., in institutional repositories or on their website) prior to and during the submission process, as it can lead to productive exchanges, as well as earlier and greater citation of published work (See The Effect of Open Access).





















