



Publicado
Efecto de la micorrización sobre el crecimiento de plántulas de plátano en sustrato con y sin la presencia de nematodos fitoparásitos
Effect of mycorrhization on the growth of plantain seedlings in substrate with and without the presence of plant parasitic nematodes
Palabras clave:
Colonización micorrícica, crecimiento radical, Musa AA, Radopholus similis. (es)Musa AAB, mycorrhizal colonization, Radopholus similis, root growth. (en)
Descargas
1Investigador visitante, Patología de Yuca y Frutas Tropicales, Centro Internacional de Agricultura Tropical (CIAT), A.A. 6713,
Cali, Colombia.
2Corporación Colombiana de Investigación Agropecuaria (Corpoica), Palmira, Valle del Cauca, Colombia.
3Investigador Asociado, Centro Internacional de Agricultura Tropical (CIAT), A.A. 6713, Cali, Colombia.
*Autor para correspondencia: legabet@gmail.com; †mbmarthabolanos@gmail.com; ‡nasakawa@cgiar.org
Se comparó el efecto de la pre-inoculación de los hongos micorrícicos arbusculares (HMA) Glomus fistulosum, Glomus fasciculatum, micorriza comercial (G. aggregatum, G. manihotis, G. fistulosum, G. fasciculatum, Kuklosplora colombiana y Scutellospora spp.) y un control sin HMA, sobre el crecimiento de propágulos vegetativos de plátano Dominico Hartón (Musa AAB) en sustrato con la presencia de Radopholus similis y Helicotylenchus spp y sin ella. Adicionalmente se evaluó el efecto de los HMA sobre la cantidad final de los nematodos fitoparásitos en raíces y suelo rizosférico. Los resultados indicaron un incremento significativo en el porcentaje de colonización micorrícica en raíces inoculadas con HMA comercial (40% - 41%), en comparación con el control y los demás HMA (< 3%). Se presentaron incrementos superiores a 30% en las variables de peso fresco de raíces y peso seco (cormo y tejido aéreo) en las plantas inoculadas con HMA comercial, aun en la presencia de nematodos fitoparásitos. La cantidad final de nematodos fitoparásitos no fue afectada por la aplicación de HMA. Se concluye que la micorrización obtenida con las especies de HMA del inóculo comercial favoreció el crecimiento radical y la absorción de nutrientes, mejoró la producción de biomasa y contrarrestó los daños causados por nematodos fitoparásitos.
Palabra clave: Colonización micorrícica, crecimiento radical, Musa AAB, Radopholus similis.
We compared the effect of pre-inoculation with the following arbuscular mycorrhizal fungi (AMF), on the growth of vegetative propagules (suckers) of plantain (Musa AAB): 1) Glomus fistulosum, 2) Glomus fasciculatum, and 3) commercial inoculum (mixture of G. aggregatum, G. fistulosum, G.manihotis, G. fasciculatum, Kuklosplora colombiana and Scutellospora spp.), compared with a control without AMF, in substrate with and without the presence of Radopholus similis and Helicotylenchus spp.. Furthermore, we evaluated the effect of AMF on the final amount of plant parasitic nematodes in roots and rhizosphere soil. The results indicated a significant intensification in the mycorrhizal colonization in roots inoculated with commercial inoculum of AMF (40 - 41%) compared with control and other AMFs (<3%). There were over 30% of increase in fresh weight of roots and dry matter (corm and air tissue) variables, in plants inoculated with commercial AMF, even in the presence of plant parasitic nematodes. The final amount of plant parasitic nematodes was not affected by the inoculation of AMF. We conclude that mycorrhizal fungi species mixture, present within the AMF commercial inoculum, stimulated root growth and nutrient uptake, thereby improving biomass production and offsetting the damage by plant parasitic nematodes.
Key words: Musa AAB, mycorrhizal colonization, Radopholus similis, root growth.
Los bananos y plátanos son el cuarto cultivo alimenticio más importante del mundo, después del arroz, el trigo y el maíz (Arias et ál., 2004). Las estadísticas de la FAO (FAO, 2009) indican un área cultivada con plátano en Colombia de aproximadamente 379.863 ha y lo ubican como el principal exportador mundial de plátano, razón por la cual esta musácea es un producto de gran importancia económica y social para el país. Las principales dificultades en la producción de este fruto en Colombia se refieren a los bajos niveles de inversión en labores de cultivo como la adecuación de fincas, la renovación, la fertilización y el drenaje, sumado a la utilización de material de siembra no certificado, por lo cual se hace evidente la aparición de problemas fitosanitarios, como el moko (Ralstonia solanaceraum Smith), la sigatoka negra (Mycosphaerella fijiensis Morelet), el picudo negro (Cosmopolites sordidus Germar) y los nematodos fitoparásitos. De acuerdo con Araya (2003) después de la sigatoka negra, el daño causado por los nematodos fitoparásitos es de gran importancia ya que reducen el rendimiento del cultivo hasta en 80%, sin dejar de tener presente asimismo las condiciones agroecológicas y la susceptibilidad del cultivar. Las especies de nematodos fitoparásitos que afectan los cultivos de banano y plátano han sido, en orden de importancia, los endoparásitos migratorios Radopholus similis (Cobb) Thorne, y Pratylenchus goodey (Sher y Allen), el ecto-endoparásito Helicotylenchus multicinctus (Cobb) Golden y diferentes especies del endoparásito sedentario Meloidogyne spp., siendo Meloidogyne incognita (Kofoid y White) Chitwood la especie de mayor importancia (Gowen et ál., 2005; Araya, 2003; Brooks, 2004; Carlier et ál., 2003).
El daño al sistema radical ocasionado por estos nematodos se traduce en baja absorción de nutrientes y agua, lo cual resulta en una reducción del peso del racimo, prolongación del ciclo vegetativo y volcamiento de las plantas, especialmente en la etapa de llenado del racimo (Sarah et ál., 1996; Quénéhervé et al., 1991; Gowen et ál, 2005). En condiciones tropicales, R. similis es el más frecuente y más abundante en cualquier estado de la planta y su infección ocurre tanto por nematodos procedentes del cormo que pasan de la planta madre a las yemas, como por nematodos procedentes del suelo (Araya, 2003).
El manejo de estos fitoparásitos se basa en la aplicación de nematicidas granulares combinada con prácticas agronómicas (Marin et ál., 2000). No obstante, los productos químicos son altamente tóxicos y su uso inapropiado o indiscriminado puede tener efectos negativos en la salud animal y humana, así como en el medio ambiente. Adicionalmente, en Colombia no se dispone de cultivares comerciales de plátano con resistencia genética a nematodos y los agricultores de plátano a pequeña escala generalmente no implementan prácticas de manejo diferentes de la remoción ocasional del tejido afectado en el cormo. Las anteriores razones muestran la importancia de evaluar e implementar alternativas de manejo que puedan ser usadas fácilmente en un programa de manejo integrado.
El uso de hongos endomicorrízicos, los cuales establecen una asociación simbiótica mutualista con la mayoría de las plantas superiores, ha mostrado ser eficiente para incrementar el crecimiento de plántulas de plátano y banano (Jaizme-Vega et ál., 2002; Elsen et ál., 2003) y los mecanismos de tolerancia contra nematodos fitoparásitos, atribuidos a la mejor nutrición y la compensación de daños en raíces (Hol y Cook, 2005; Jaizme-Vega y Rodríguez-Romero, 2004; Jaizme-Vega et ál., 1997). Por lo anterior, el objetivo del presente estudio fue evaluar el efecto de la pre-inoculación con tres hongos formadores de micorriza arbuscular (HMA) sobre el crecimiento inicial de propágulos vegetativos de plátano clon Dominico Hartón en sustrato, con la presencia de nematodos fitoparásitos y sin ella, bajo condiciones de casa de malla. Adicionalmente, se evaluó el efecto de los tratamientos con HMA sobre la cantidad final de nematodos fitoparásitos en suelo rizosférico y en raíces.
Sustrato de siembra y localización del experimento
El sustrato de siembra consistió de una mezcla en relación 2:1 (suelo/arena de río). El suelo se recolectó en la estación experimental del Centro Internacional de Agricultura Tropical (CIAT) en Santander de Quilichao (Colombia), caracterizado por bajo contenido de fósforo, el cual se esterilizó con vapor de agua por dos horas diarias durante dos días. La composición química de este sustrato aparece en el Cuadro 1. Después de la esterilización, el sustrato fue empacado en bolsas de polietileno con capacidad para 5 kg que fueron colocadas en condiciones de casa de malla en el Centro de Investigación Palmira de Corpoica, localizado en Palmira (3° 31' N, 76° 19' O, a 1001 m.s.n.m y 24°C), Valle del Cauca, Colombia.

Inóculo de HMA
Las fuentes de inóculo de HMA fueron aplicadas en cada bolsa manualmente, de la forma siguiente: (1) Glomus fasciculatum (Thaxt.) Gerd. y Trappe emend. C. Walker y Koske; (2) Glomus fistulosum Skou y Jakobsen, ambas especies obtenidas del Banco de Aislamientos de HMA del CIAT; y (3) inóculo comercial consistente en la mezcla de las especies G. fasciculatum, G. fistulosum, G. aggregatum Schenck y G.S. Smith, G. manihotis Howeler, Sieverding y Schenk, Kuklosplora colombiana Oehl y Sieverding y Scutellospora spp. Walker y Sanders. El inóculo consistió de sustrato, fragmentos de raíces colonizadas, esporas, esporocarpos y fragmentos de hifas extrarradicales. La cantidad de inóculo aplicado se estableció con base en el número de esporas de HMA contenidas en 20 g del inóculo comercial dosis comercial recomendada para cultivo en vivero determinada mediante extracción y cuantificación con utilización del método de flotación en azúcar. En cada bolsa con sustrato se aplicó una dosis de inóculo con 283 ± 43 esporas de HMA. El tratamiento control consistió sólo en el sustrato estéril.
Después de la inoculación con HMA, en cada bolsa se sembró un propágulo vegetativo de plátano Dominico Hartón (Musa AAB Simmonds) de aproximadamente 10 cm de altura y 3 cm de ancho, obtenidos mediante inducción de la brotación de yemas laterales en cormos, en condiciones de alta temperatura (50 °C aprox.). La humedad del suelo con el propágulo fue mantenida cerca y siempre por debajo de capacidad de campo para evitar cualquier estrés hídrico.
Inóculo de nematodos fitoparásitos
Se recolectaron raíces de plátano (Musa AAB Simmonds) infectadas por nematodos en fincas en los municipios de Montenegro y Armenia, Quindío (Colombia). Los nematodos fitoparásitos fueron extraídos utilizando el método de macerado y decantación descrito por Niblack y Hussey (1987) e identificados con base en las características morfológicas y morfométricas (Mai y Mullin, 1996) como R. similis y Helicotylenchus spp., los cuales fueron concentrados, contados en estereoscopio utilizando alícuotas de 1 mL y ajustados en un volumen final a una concentración (individuos/ml de agua) de 167 R. similis y 24 Helicotylenchus spp. La inoculación de estos nematodos se hizo dos meses después de la siembra de los propágulos vegetativos, para lo cual se hicieron tres orificios en el suelo de 1 cm de diámetro, 2 cm de profundidad distantes 1.5 cm desde la base del pseudotallo (Dosselaere et ál., 2003; Zum Felde et ál., 2006). En cada orificio se aplicaron 2 ml de la suspensión con nematodos, inoculando, así, una cantidad aproximada de 1000 R. similis y 144 Helicotylenchus spp. por planta. En el tratamiento control, sin la presencia de nematodos, se añadió un volumen igual de agua estéril.
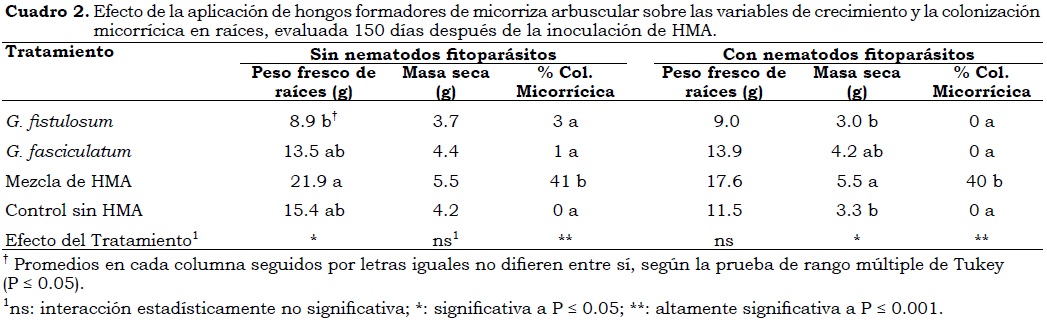
Los tratamientos en el experimento se dispusieron en un diseño completamente aleatorizado, en el cual se comparó el efecto de los HMA: G. fistulosum, G. fasciculatum, micorriza comercial (mezcla G. aggregatum, G. manihotis G. fistulosum, G. fasciculatum, Kuklosplora colombiana y Scutellospora spp.) y un control sin HMA sobre el crecimiento de propágulos vegetativos de plátano en sustrato con la presencia de nematodos y sin ella (Cuadro 2). Cada tratamiento fue replicado cuatro veces utilizando cuatro plantas como unidad experimental.
Las plantas fueron removidas noventa días después de la inoculación y los resultados de infestación sobre las variables de crecimiento de la planta fueron, peso fresco radical (g) y peso seco (cormo más tejido aéreo) (g).
La cantidad final de nematodos en suelo se determinó en una submuestra de 100 cm3 de suelo rizosférico en cada planta, utilizando el método de extracción mediante centrifugación y flotación en azúcar descrito por Niblack y Hussey (1987). Para determinar el número de nematodos en raíces, éstas fueron cortadas en fragmentos de 2 - 3 cm de largo y luego de homogenizadas se utilizó una submuestra de 10 g la cual fue macerada en una licuadora durante tiempos iguales de 10 segundos a baja y alta velocidad y, posteriormente, se utilizó el método de centrifugación y flotación en azúcar para extraer los nematodos (Niblack y Hussey, 1987). La cantidad de nematodos fue evaluada en tres alícuotas de 5 ml de una muestra de 25 ml, utilizando una cámara de conteo y estereoscopio. La identificación de R. similis y Helicotylenchus spp. se realizó de la forma anteriormente mencionada. Finalmente, la cantidad de nematodos fue expresada en 100 cm3 de suelo y 1 g de raíces.
La colonización micorrícica de HMA en raíces fue determinada en segmentos de raíces de 1cm, clareadas con KOH (2.5%) en baño María (90 °C) durante 1 h; posteriormente, se sumergieron en HCl (2%) durante 1 h a temperatura ambiente, teñidas con azul de tripano al baño María (90 °C) durante 1 h, montadas en placas porta-objetos y evaluadas en microscopio (objetivo de 40X) mediante la escala de cinco clases de Trouvelot (Trouvelot et ál., 1986) (Figura 1). Los resultados fueron analizados con el programa Mycocalc (Clapp y Zhao, 2001) para calcular el porcentaje de colonización micorrícica (CM).

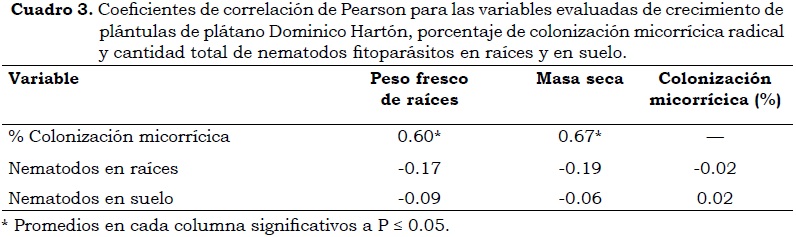
Antes de realizar el análisis, la cantidad de nematodos fitoparásitos fue transformada [log10 (x + 1)] para ajustar los datos a una distribución normal. Las diferencias entre los tratamientos fueron evaluadas con Anava, usando el procedimiento GLM (modelo lineal general) con los tratamientos de inóculos de HMA como fuente de variación. Las medias de los datos fueron separadas mediante la prueba de comparación de Tukey (P < 0.05), así mismo, se hicieron correlaciones múltiples de Pearson entre las variables evaluadas y la cantidad de nematodos fitoparásitos en suelo y raíz y la colonización micorrícica. Todos los análisis se hicieron con el paquete estadístico SAS versión 9.1 (SAS Institute Inc, 2004).
En ausencia de nematodos se observó un incremento significativo en la producción de raíces con el tratamiento de HMA comercial, el cual fue dos veces mayor que el obtenido con la cepa G. fistulosum (Cuadro 2). Las plantas con inóculo de HMA comercial incrementaron el peso fresco radical promedio aproximadamente en un 60% y 40% en comparación con las plantas que no recibieron HMA y las que recibieron G. fasciculatum, respectivamente; no obstante, estas diferencias no fueron significativas. El contenido de masa seca de cormo y parte aérea de las plántulas no fue diferente (P > 0.05) entre los tratamientos; sin embargo, se observa que las plantas inoculadas con la mezcla de HMA incrementaron su biomasa en un 30% en comparación con el control sin HMA.
El porcentaje de colonización micorrícica fue significativo (P < 0.05) entre tratamientos, el cual fue del 41% en las plantas inoculadas con HMA comercial, mientras que el control sin HMA y las inoculadas con G. fistulosum y G. fasciculatum presentaron menos del 3% de colonización (Cuadro 2). El incremento en raíces y biomasa estuvo correlacionado positivamente con el porcentaje de colonización micorrícica (Cuadro 3), lo cual demuestra el efecto benéfico de la micorriza en plántulas de plátano Dominico Hartón. Resultados similares se encontraron en las variables de crecimiento en plántulas cuando se inocularon nematodos fitoparásitos en el suelo rizosférico. En este caso, se presentó un incremento significativo en la masa seca de las plantas micorrizadas con el inóculo de HMA comercial, superior a 30%, en comparación con los demás tratamientos. El mayor peso fresco de raíces fue obtenido en las plántulas que recibieron el inóculo comercial, no obstante las diferencias no fueron significativas (P > 0.05). Los incrementos en peso fresco de raíz y masa seca de las plántulas inoculadas con HMA comercial y nematodos fitoparásitos fueron superiores al presentado en el testigo sin nematodos. La micorrización incrementa el crecimiento y favorece una serie de cambios a nivel morfológico en raíces, lo cual hace que las plantas sean más tolerantes al daño por nematodos gracias a que el incremento en la ramificación de las raíces en plantas micorrizadas contrarresta los impactos negativos de los nematodos (Hol y Cook, 2005).
La ausencia en este estudio de una buena asociación simbiótica entre G. fistulosum o G. fasciculatum con las plantas de plátano no es explicable, ya que en otros estudios (Reyes et al., 1995; Hurtado, 1996; Jaramillo y Rivillas, 2001) se encontraron colonizaciones superiores al 80% en raíces de plantas de plátano y banano con estas especies. Igualmente se citan efectos de bioprotección de G. fasciculatum contra R. similis en banano (Siddiqui y Mahmood, 1995; Umesh et ál., 1988). Posiblemente, las cepas de G. fistulosum y G. fasciculatum utilizadas en el presente estudio requieren una mayor cantidad de esporas/g de inóculo, o que hayan sido afectadas por condiciones físicas y químicas del suelo que no les permitieron alcanzar niveles adecuados de adaptación e inicio de la colonización, comparado con algún género o especie presente en el inóculo de HMA comercial. Esto sugiere que la inoculación de una única cepa no puede ser más efectiva que la inoculación de una mezcla de cepas. En la actualidad se acepta la especificidad entre huésped y cepa inoculada en la asociación micorrícica, incluso existen evidencias en la diversidad de respuesta entre esporas provenientes de la misma especie (Ian Sanders, University of Lausanne, Suiza, 2010 -información personal). Elsen et ál (2003) indican que la dependencia relativa de micorriza (DRM), definida como el grado de dependencia de la planta a la micorriza para producir un máximo crecimiento o mayor producción a un determinado nivel de fertilidad del suelo, es dependiente del cultivar y del HMA. De hecho, se ha observado una gran variación en DMR entre siete cultivares de Musa spp. (grupo AAA), inoculados con dos especies de HMA en condiciones de invernadero (Declerck et ál, 1995). El DRM también fue muy variable de acuerdo con la especie de Glomus spp. inoculada (Declerck et ál., 1995).
Los resultados obtenidos confirman el beneficio en el crecimiento de plántulas de Musa spp. (Declerck et ál., 1995; Rizzardi, 1990; Elsen et ál., 2002; Jaizme-Vega et ál., 2002), como respuesta a la micorrización presentada por las especies de HMA del inóculo comercial. En este caso, el incremento se asoció con una mayor absorción de nutrientes, ya que se encontró que las plantas micorrizadas presentaron mayores contenido de fósforo (P), potasio (K), calcio (Ca), magnesio (Mg), y cobre (Cu) que el mostrado por las plantas sin micorrización (datos no incluidos), lo cual coincide con los resultados en otras investigaciones (Allen, 1996; Smith y Read, 1997; Pinochet et ál, 1997; Clark y Zeto, 2000; Ramos et ál., 2009).
En este estudio, la colonización micorrícica no fue afectada por la ausencia o presencia de nematodos fitoparásitos, lo cual se demuestra por el bajo índice de correlación obtenido entre ambas variables (Cuadro 3) y por el porcentaje de colonización obtenido con HMA comercial (Cuadro 2) el cual fue similar en un 40 y un 41% respectivamente, en suelo con nematodos fitoparásitos y sin ellos. Elsen et ál (2002) igualmente encontraron que la intensidad de la asociación micorrícica no fue afectada en presencia de R. similis y P. coffeae. No obstante, los nematodos fitoparásitos pueden degradar las células que contribuyen a la nutrición de los HMA (Hol y Cook, 2005). Este efecto puede presentarse con un mayor tiempo de exposición de las raíces a este tipo de nematodos, lo cual es necesario para una mayor degradación de células radicales.
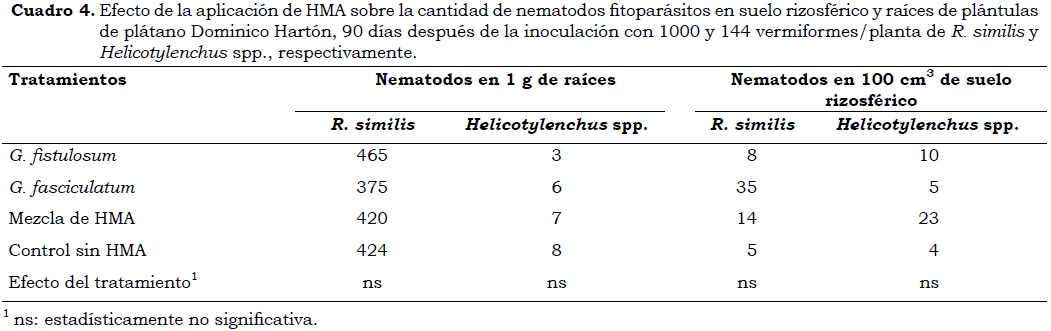
En cuanto a la cantidad de nematodos fitoparásitos en raíces y suelo, los resultados demuestran que el método de inoculación fue adecuado. En general, se obtuvieron, en promedio, 5473 vermiformes de R. similis en el promedio total del sistema radical (13 g/ planta), demostrando un incremento mayor a cinco veces de la cantidad inicial inoculada (1000 R. similis). De igual forma, se demostró la sanidad del material de siembra utilizado, ya que el sistema radical de los propágulos vegetativos sembrados en suelo sin nematodos permaneció sano sin la presencia de nematodos en raíces o en suelo rizosférico. Lo anterior demuestra la importancia de utilizar material de siembra limpio de patógenos, con lo cual se disminuye el inóculo inicial que puede ser un factor crítico en la plantación en campo.
La ventaja en el uso del sistema de propagación por inducción de rebrotes y termoterapia es la cantidad de material que se puede producir a partir de pequeñas cantidades de cormos sanos de plátano en un corto tiempo y lo asequible que resulta para pequeños agricultores por su bajo costo (menos de US$0.35/ planta) (González, 2011).
En los tratamientos que incluyeron inoculación de nematodos, la cantidad final de estos vermiformes tanto en suelo rizosférico como en las raíces, no evidenció diferencias (P < 0.05) entre los diversos tratamientos (Cuadro 4). No obstante, la cantidad total de nematodos fue mayor en las raíces de plántulas inoculadas con G. fistulosum (424 R. similis/g raíces) lo cual puede explicar porqué se obtuvieron los valores más bajos en las variables de crecimiento, incluso menores que las presentadas con el testigo sin HMA. En suelo rizosférico, la cantidad promedio de nematodos fitoparásitos fluctuó entre 3 y 24 vermiformes/100 cm3 de suelo (P < 0.05). Que la cantidad de nematodos presentes en la raíz fue mayor a la observada en el suelo está relacionado con el hábito endoparásito de R. similis y ecto-endoparásito de Helicotylenchus spp. (Gowen et ál, 2005).
Tanto en plátano como en banano se ha encontrado que los HMA pueden proveer protección contra nematodos fitoparásitos, a través de diferentes modos de acción, tales como reducción de los daños o incremento en la tolerancia de la planta a la enfermedad (Jaizme-Vega y Pinochet, 1997; Jaizme-Vega y Rodríguez-Romero, 2004), reducción en la población de nematodos (Bagyaraj et ál, 1979; Jaizme-Vega et ál., 1997; Elsen et ál., 2002; Elsen et ál., 2003; Elsen et ál., 2008; Oyekanmi et ál., 2008; Van der Veken et ál., 2010; Becerra-Encinales et ál., 2010), e inducción de resistencia sistémica (Elsen et ál, 2008; Zhang et ál., 2008).
En el presente estudio no se observó reducción en la población de nematodos en raíces, lo cual concuerda con los hallazgos de Hol y Cook (2005) quienes en su trabajo con nematodos migratorios, R. similis y P. coffeae, encontraron que estos son el único grupo cuyos números son altos en plantas colonizadas con HMA. En este caso, el efecto de protección se asocia a una alteración directa en el crecimiento de la planta, a su morfología y a su nutrición (Zhang et ál., 2008; Whipps, 2004) lo cual hace que las plantas sean más tolerantes al daño por nematodos (Hol y Cook, 2005).
- La inoculación de una única especie de HMA posiblemente no es más efectiva que cuando se inocula una mezcla de especies de HMA, lo que sucedió en este estudio con el inóculo comercial.
- Los resultados indican que el incremento en raíces y biomasa estuvo correlacionado positivamente con el porcentaje de colonización micorrícica obtenido con la preinoculación de esporas de las especies G. aggregatum, G. manihotis, G. fistulosum, G. fasciculatum, K. colombiana y Scutellospora spp., presentes en el inóculo comercial de HMA, aun en presencia de nematodos.
- Este trabajo permitió demostrar que no existe un efecto de la micorrización en la supresión del desarrollo y la reproducción de los nematodos fitoparásitos. El efecto de bioprotección se asoció más a una alteración directa en el crecimiento de la planta, a su morfología y a su nutrición, lo cual hace que las plantas sean más tolerantes al daño por nematodos. No obstante, el tipo y magnitud de la respuesta varían con el ambiente, el genotipo del hospedero, las especies y poblaciones de nematodos y los aislamientos del hongo (Hol y Cook, 2005).
- De esta manera, se plantea el uso de propágulos vegetativos sanos a partir de inducción de rebrotes y termoterapia y la adición de HMA, como prácticas para disminuir la dispersión de nematodos en el campo e incrementar el crecimiento vegetativo en almácigo, dentro de un plan de manejo integrado del cultivo del plátano.
Este trabajo hizo parte del proyecto Mejoramiento de la sanidad y competitividad del cultivo de plátano, mediante el desarrollo de nuevas estrategias de manejo del suelo y la nutrición, financiado por el Fondo Nacional de Fomento Hortofrutícola y el Ministerio de Agricultura y Desarrollo Rural. Se agradece a la fitopatóloga Francia Varón, a Silverio González, presidente de Fedeplátano, por sus contribuciones a este trabajo y a los jurados de revisión del presente artículo por sus valiosos aportes.
Allen, M. F. 1996. The ecology of arbuscular mycorrhizas: a look back into 20th century and a peak into the 21st century. Mycorr. Res. 100:769 - 782.
Araya, M. 2003. Situación actual del manejo de nematodos en banano (Musa AAA) y plátano (Musa AAB) en el trópico americano. Actas del taller manejo convencional y alternativo de la Sigatoka negra, nematodos y otras plagas asociadas al cultivo de Musáceas. Guayaquil, Ecuador. p. 79-102.
Arias, P.; Liu, P.; y Dankers, C. 2004. La economía mundial del banano 1985-2002. Roma: Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). p. 1.
Bagyaraj, D. J.; Manjunath, A.; y Reddy, D. D. R. 1979. Interaction of vesicular arbuscular mycorrhizas with root knot nematodes in tomato. Plant Soil 51:397 - 403.
Becerra-Encinales, J. F.; Castaño-Zapata, J.; y Villegas-Estrada, B. 2010. Efecto de la micorrización sobre el manejo de nematodos en plántulas de plátano híbrido FHIA-20AAAB. Agronomía 18(1):7 - 18.
Brooks, F. E. 2004. Plant-parasitic nematodes of banana in American Samoa. Nematropica 34:65 - 72.
Carlier, J.; De Waele, D.; y Escalant, J. V. 2003. Evaluación global de la resistencia de los bananos al marchitamiento por Fusarium, enfermedades de las manchas foliares causadas por Mycosphaerella y nematodos. Evaluación de comportamiento. Vézina, A.; y Picq, C. (eds.). Guías técnicas INIBAP 7. Red Internacional para el Mejoramiento del Banano y el Plátano, Montpellier, Francia.
Clapp, J.; y Zhao, B. 2001. Manual de micorrizas. Preparado para el taller: Hongos micorricicos Arbúsculares en los Sistemas de Producción Vegetal: detección, taxonomía, conservación y ecofisiología. Wuhan, PR China. Laboratorio de Microbiología Agrícola, Universidad de Huazhong, Wuhan, PR China. Disponible en http://www2.dijon.inra.fr/mychintec/Protocole/Workshop_ Procedures.html#1.5 [Fecha de revisión Marzo de 2011]
Clark, R .B.; y Zeto, S. K. 2000. Mineral acquisition by arbuscular mycorrhizal plants. J. Plant Nutr. 23:867 - 902.
Declerck, S.; Plenchette, C.; y Strullu, D. G. 1995. Mycorrhizal dependency of banana (Musa acuminata, AAA group) cultivars. Plant Soil 176:183 - 187.
Dosselaere, N.; Araya, M.; y De Waele, D. 2003. Efecto del pote sobre el crecimiento de las raíces, el potencial de reproducción de R. similis y los daños que ocasiona a los bananos. Infomusa 12(1):17 - 21.
Elsen, A.; Declerck, S.; y De Waele, D. 2002. Effect of three arbuscular mycorhizal fungi on root-knot nematode (Meloidogyne spp.) infection of Musa. Infomusa 11(1):21 - 23.
Elsen, A.; Baimey, H.; Swennen, R.; y De Waele, D. 2003. Relative mycorrhizal dependency and mycorrhiza- nematode interaction in banana cultivars (Musa spp.) differing in nematode susceptibility. Plant Soil 256:303 - 313.
Elsen, A.; Gervacio, D.; Swennen, R.; y De Waele, D. 2008. AMF-induced biocontrol against plant parasitic nematodes in Musa sp.: a systemic effect. Mycorrhiza 18:251 - 256.
FAO. 2009. Faostat. Disponible en: http://faostat.fao.org/site/339/default.aspx [Fecha de revisión Marzo de 2011].
González, S. 2011. Federación Nacional de Plataneros (Fedeplátano). Bol. Musalac 2(2):11 - 12. Disponible en: http://bananas.bioversityinternational.org/files/files/pdf/publications/BoletinMUSALACV2N2.pdf. [Fecha de revisión Octubre 15 de 2011].
Gowen, S. R.; Quénéhervé, P.; y Fogain, R. 2005. Nematode parasites of bananas and plantains. En: Luc, M.; Sikora, R. A.; Bridge, J. (eds.). Plant parasitic nematodes in subtropical and tropical agriculture. CABI, Wallingford. p 611 - 643.
Hol, G. W. H.; y Cook, R. 2005. An overview of arbuscular mycorrhizal fungi-nematode interactions. Basic. Appl. Ecol. 6:489 - 503.
Hurtado, J. 1996. Efecto de Glomus fistulosum y Scutellospora heterogama en el crecimiento y desarrollo del plátano Musa sp. Grupo AAB cv Dominico Hartón. Tesis (Adm. Agr.) Universidad Católica de Oriente, Rionegro (Colombia). 119 p.
Jaizme-Vega, M. C.; y Pinochet, J. 1997. Growth response of banana to three mycorrhizal fungi in Pratylenchus goodeyi infested soil. Nematropica 27(1):69 - 76.
Jaizme-Vega, M. C.; y Rodriguez-Romero, A. S. 2004. Uso de micorrizas en banano: logros y perspectivas. En: Memorias XVI Reunión Internacional Acorbat. Oaxaca, México. p 143 - 160.
Jaizme-Vega, M. C.; Tenoury, P.; Pinochet, J.; y Jaumot, M. 1997. Interaction between the rootknot nematode Meloidogyne incognita and the mycorrhizal association of Glomus mossae and Grande Naine banana. Plant and Soil 196:27 - 35.
Jaizme-Vega, M. C.; Esquivel, M.; Tenoury, P.; y Rodriguez, A. S. 2002. Effects of mycorrhization on the development of two cultivars of micropropagated banana. Infomusa 11(1):25 - 28.
Jaramillo, M. M.; y Rivillas, C. A. 2001. Efecto de Glomus manihotis y Glomus fistulosum en el manejo de nematodos fitopatógenos en plantas micropropagadas de plátano y banano en Colombia. En: Memorias 22 Congreso de la Asociación de Fitopatología y Ciencias Afines. Medellín, Colombia. 22:23.
Mai, W. F.; y Mullin, P. G. 1996. Plant-parasitic nematodes. A pictoria key to genera. 5th ed. Cornell University Press.
Marin, D. H.; Barker, K. R.; Kaplan, D. T.; Sutton, T. B.; y Opperman, C. H. 2000. Development and evaluation of a standard method for screening for resistance to Radopholus similis in bananas. Plant Dis. 84:689 - 693.
Niblack, T. L.; y Hussey, R. S. 1987. Extracción de nematodos del suelo y tejidos vegetales. Department of Plant Pathology. University of Georgia, Pp. 235-243. En: Fitonematología, Manual de Laboratorio. CATIE. Turrialba, Costa Rica. 248 p.
Oyekanmi, E. O.; Coyne, D. L.; y Fawole, B. 2008. Utilization of the potentials of selected microorganisms as biocontrol and biofertilizer for enhanced crop improvement. J. Biol. Sci. 8(4):746 - 752.
Pinochet, J.; Fernández, C.; Jaizme-Vega, M. C.; y Tenoury, P. 1997. Micropropagated banana infected with Meloidogyne javanica responds to Glomus intraradices and phosphorus. Hort. Sci. 32(1):101 - 103.
Quénéhervé, P.; Cadet, P.; Mateille, T.; y Topart, P. 1991. Population of nematodes in soils under banana cv. Poyo, in the Ivory Coast. 5. Screening of nematicides and horticultural results. Rev. Nematol. 14:231 - 249.
Ramos, A. C.; Martins, M. A.; Okorokova-Façanha, A. L.; Lopes, F.; Okorokov, L. A.; Sepúlveda, N.; Féijo, J. A.; y Façanha, A. R. 2009. Arbuscular mycorrhizal fungi induce differential activation of the plasma membrane and vacuolar H+ pumps in maize roots. Mycorrhiza 19:69 - 80.
Rizzardi, V. 1990. Effect of inoculation with vesiculararbuscular mycorrhizal fungi on the growth of micropropagated Musa acuminata clone Grand Nain. Rev. Agric. Subtrop. Trop. 84(3):473 - 484.
Sarah, J. L.; Pinochet, J.; y Stanton, J. 1996. Radopholus similis Cobb, el nematodo barrenador del banano. plagas de Musa. INIBAP, Montpellier, Francia. Hoja divulgativa No.1.
SAS Institute Inc. 2004. SAS/STAT 9.1 Users Guide, Version 8. SAS Institute, Cary, NC.
Siddiqui, Z. A.; y Mahmood, I. 1995. Biological control of Heterodera cajani and Fusarium udum by Bacillus subtilis, Bradyrhizobium japonicum and Glomus fasciculatum on pigeon pea. Fundam. Appl. Nematol. 18:559 - 566.
Smith, S. E.; y Read, D. J. 1997. Mycorrhizal Symbiosis, second ed., Academic Press, Londres.
Trouvelot, A.; Kough, J. L.; y Gianinazzi-Pearson, V. 1986. Mesure du taux de mycorrhization VA dun systeme radiculaire. Reserche de methodes destimation ayant une signification fonctionelle. En Physiological and genetical aspects of mycorrhizae. Gianinazzi-Pearson, V. and Gianinazzi, S (eds.). INRA, Paris. p: 101-109.
Umesh, K. C.; Krishnappa, K.; y Bagyaraj, D. J. 1988. Interaction of burrowing nematode, Radopholus similis (Cobb, 1893) Thorne 1949, and VA mycorrhiza, Glomus fasciculatum (Thaxt.) Gerd. and Trappe in Banana (Musa acuminata Colla.). Indian J. Nematol. 18(1):6 - 11.
Van der Veken, L.; PaWinb, P.; Elsen, A.; Swennen, R.; y DeWaele, D. 2010. Susceptibility of banana intercrops for rhizobacteria, arbuscular mycorrhizal fungi and the burrowing nematode Radopholus similis. App. Soil Ecol. 40:283 - 290.
Whipps, J. M. 2004. Prospects and limitations for mycorrhizals in biocontrol of root pathogens. Can. J. Bot. 82:1198 - 1227.
Zhang, L.; Zhang, J.; Christie, P.; y Li, X. 2008. Preinoculation with arbuscular mycorrhizal fungi suppresses root knot nematode (Meloidogyne incognita) on cucumber (Cucumis sativus). Biol. Fertil. Soils 45:205 - 211.
Zum Felde, A.; Pocasangre, L. E.; Carñizares-Monteros, C. A.; Sikora, R. A.; Rosales, F. E.; y Riveros, A. S. 2006. Efecto de inoculaciones combinadas de hongos endofíticos en el biocontrol de Radopholus similis. Infomusa 15(1-2):12 - 18.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Visitas a la página del resumen del artículo
Descargas
Licencia
Derechos de autor 2011 Acta Agronómica

Esta obra está bajo una licencia internacional Creative Commons Atribución-NoComercial-SinDerivadas 4.0.
Política sobre Derechos de autor:Los autores que publican en la revista se acogen al código de licencia creative commons 4.0 de atribución, no comercial, sin derivados.

Es decir, que aún siendo la Revista Acta Agronómica de acceso libre, los usuarios pueden descargar la información contenida en ella, pero deben darle atribución o reconocimiento de propiedad intelectual, deben usarlo tal como está, sin derivación alguna y no debe ser usado con fines comerciales.





















