


Publicado
Tamaño del fragmento de bosque y aislamiento: potenciales efectos sobre ensambles de escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae)
Forest fragment size effects and isolation: potential effects on ensembles of dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae)
DOI:
https://doi.org/10.15446/caldasia.v45n3.99369Palabras clave:
aislamiento, biodiversidad, conectividad, conservación, deforestación (es)conservation biology, conectivity, deforestation, biodiversity, isolation (en)
Aunque existe abundante información sobre los efectos nocivos de la fragmentación, muchos interrogantes persisten sobre el efecto del tamaño de los relictos de bosque sobre la fauna. Con el objetivo de evaluar el efecto de este factor sobre la diversidad, riqueza, biomasa y composición del ensamble de escarabajos coprófagos, se realizaron muestreos con trampas de caída cebadas con excremento humano en seis fragmentos de bosque con tamaños entre 13 y 231 ha en un paisaje andino. El relicto de bosque más grande mostró mayor biomasa y diversidad de escarabajos, mientras la abundancia fue mayor en fragmentos de tamaño intermedio. La abundancia de diez especies de las 39 observadas se correlacionó con el tamaño del fragmento y los grupos más sensibles según la biomasa son los escarabajos más pesados, mayores a 0,5 gramos de peso seco, y los de peso intermedio, entre 0,031 a 0,12 gr. Los índices de similitud muestran que todos los fragmentos contribuyen a la biodiversidad regional y cada uno alberga un ensamble distinto, lo que demuestra que la matriz de pasturas y rastrojos es una barrera difícil para la mayoría de las especies. Las dos especies de escarabajos más pesadas Oxysternon conspicillatum y Deltochilum gibossum aportan el 31% de la biomasa de los escarabajos y son afectadas por la reducción del fragmento de bosque lo que podría tener consecuencias negativas en las funciones ecológicas que desempeñan este grupo de insectos.
Although abundant information exists on the harmful effects of fragmentation, many questions remain about the effect of the size of forest remnants on wildlife. To evaluate this effect on the diversity, richness, biomass, and composition of the coprophagous beetle assemblage, six forest remnants were sampled, ranging in size from 13 to 231 ha, in the northeast of the department of Antioquia in Colombia with pitfall traps baited with human dung. The largest forest fragment has the highest values of beetle biomass and diversity, while abundance was highest in intermediate-sized fragments. The abundance of ten species of the 39 observed correlates with the size of the fragment and the groups most sensitive according to biomass are the heaviest beetles, greater than 0.5 grams of dry weight, and those of intermediate weight between 0.031 to 0.12. Similarity indices show that all fragments, contribute to regional biodiversity, and each one has a different assembly, showing that the matrix of pastures and regeneration of different ages is a difficult barrier for most beetle species. The two heaviest species of beetles, Oxysternon conspicillatum and Deltochilum gibossum, contribute 31% of the beetle biomass and are affected by the reduction of the forest fragment, which could have negative consequences on the ecological functions performed by this group of insects.
Recibido: 13 de noviembre de 2021; Aceptado: 7 de febrero de 2023
RESUMEN
Aunque existe abundante información sobre los efectos nocivos de la fragmentación, muchos interrogantes persisten sobre el efecto del tamaño de los relictos de bosque sobre la fauna. Con el objetivo de evaluar el efecto de este factor sobre la diversidad, riqueza, biomasa y composición del ensamble de escarabajos coprófagos, se realizaron muestreos con trampas de caída cebadas con excremento humano en seis fragmentos de bosque con tamaños entre 13 y 231 ha en un paisaje andino. El relicto de bosque más grande mostró mayor biomasa y diversidad de escarabajos, mientras la abundancia fue mayor en fragmentos de tamaño intermedio. La abundancia de diez especies de las 39 observadas se correlacionó con el tamaño del fragmento y los grupos más sensibles según la biomasa son los escarabajos más pesados, mayores a 0,5 gramos de peso seco, y los de peso intermedio, entre 0,031 a 0,12 gr. Los índices de similitud muestran que todos los fragmentos contribuyen a la biodiversidad regional y cada uno alberga un ensamble distinto, lo que demuestra que la matriz de pasturas y rastrojos es una barrera difícil para la mayoría de las especies. Las dos especies de escarabajos más pesadas Oxysternon conspicillatum y Deltochilum gibossum aportan el 31% de la biomasa de los escarabajos y son afectadas por la reducción del fragmento de bosque lo que podría tener consecuencias negativas en las funciones ecológicas que desempeñan este grupo de insectos.
Palabras clave
aislamiento, biodiversidad, conectividad, conservación, deforestación.ABSTRACT
Although abundant information exists on the harmful effects of fragmentation, many questions remain about the effect of the size of forest remnants on wildlife. To evaluate this effect on the diversity, richness, biomass, and composition of the coprophagous beetle assemblage, six forest remnants were sampled, ranging in size from 13 to 231 ha, in the northeast of the department of Antioquia in Colombia with pitfall traps baited with human dung. The largest forest fragment has the highest values of beetle biomass and diversity, while abundance was highest in intermediate-sized fragments. The abundance of ten species of the 39 observed correlates with the size of the fragment and the groups most sensitive according to biomass are the heaviest beetles, greater than 0.5 grams of dry weight, and those of intermediate weight between 0.031 to 0.12. Similarity indices show that all fragments, contribute to regional biodiversity, and each one has a different assembly, showing that the matrix of pastures and regeneration of different ages is a difficult barrier for most beetle species. The two heaviest species of beetles, Oxysternon conspicillatum and Deltochilum gibossum, contribute 31% of the beetle biomass and are affected by the reduction of the forest fragment, which could have negative consequences on the ecological functions performed by this group of insects.
Keywords
conservation biology, conectivity, deforestation, biodiversity, isolation.INTRODUCCIÓN
Los Andes tropicales son considerados un área prioritaria para la conservación de la biodiversidad, debido a su alto endemismo de especies (Myers et al. 2000). A pesar de su importancia, la destrucción del área de bosque es notable, para los años 2016 y 2017 se registraron 45 606 y 36 745 ha deforestadas respectivamente (MinAmbiente c2017), esta situación es especialmente crítica en los bosques andinos y subandinos que conservan solo una pequeña porción (< 10 %) de los bosques originales (Armenteras et al. 2003).
La destrucción y modificación de los bosques naturales y su fragmentación cambian la composición y estructura de la biota (Santos y Telleria 2006). Estos cambios tienden a ser más drásticos en ciertos grupos biológicos como los escarabajos coprófagos (Coleoptera: Scarabaeinae) que muestran una alta especificidad de hábitat (Spector y Ayzama 2003, Nichols et al. 2007); y en los cuales la mayoría de las especies están adaptadas a la estabilidad micro climática del interior de los bosques neotropicales (Gill 1991). En ensambles de escarabajos coprófagos es notable la disminución de la riqueza y diversidad con la destrucción del bosque (Halffter y Arellano 2002, Andresen 2003, Spector 2006), la alteración de las funciones ecológicas que realizan estos animales, como la descomposición del excremento para reincorporarlo al ciclo de nutrientes (Horgan 2005, Noriega et al. 2021) y la dispersión secundaria de semillas (Andresen 2003, Nichols et al. 2008).
Además de la pérdida de hábitat, otros procesos menos evidentes, pero igualmente importantes, son responsables de las causas negativas de la fragmentación sobre la biodiversidad, como la reducción del tamaño promedio de los fragmentos (Santos y Telleria 2006). Casi todos los grupos de artrópodos sufren cambios en la composición y estructura de sus comunidades cuando se reduce el tamaño de los fragmentos de bosque que habitan (Vasconcelos y Bruna 2012). Las especies sensibles a estos cambios tienden a reducir el tamaño de las poblaciones hasta el punto de poner en riesgo su viabilidad (Santos y Telleria 2006). En términos generales se espera que fragmentos de bosque de mayor tamaño alberguen un número mayor de especies (Primack et al. 2001, Santos y Telleria 2006). Por ejemplo, en la Amazonia, Klein (1989) y Andresen (2003) y en la Orinoquía Rodríguez-García et al. (2021) encontraron que el número de especies y abundancia de escarabajos coprófagos fue menor en remanentes de bosque que en bosques grandes y continuos. Amézquita et al. (1999), también reportaron un aumento en los valores de abundancia y riqueza a medida que el tamaño del fragmento es mayor.
En mamíferos se ha encontrado que las reservas naturales pequeñas (<100 ha), aunque dan refugio a especies grandes y medianas conservan principalmente mamíferos pequeños (López-Arévalo et al. 2021). Por lo tanto, la dependencia de los escarabajos coprófagos grandes por porciones de excremento que solo pueden proporcionar mamíferos grandes (Peck y Howden 1984), podría explicar la pérdida de grandes telecopridos y paracopridos en fragmentos pequeños (Rodríguez-García et al. 2021), y la reducción del tamaño promedio de escarabajos al disminuir el tamaño del relicto (Klein 1989; Andresen 2003).
Sin embargo, el papel de las reservas pequeñas y dispersas en la conservación de biodiversidad regional de insectos no se debe subestimar, debido a que estos organismos tienen menor requerimiento de área que los vertebrados, y además poseen una alta diversidad beta (Shafer 1995). Pequeñas reservas pueden sostener una cantidad importante de las especies de artrópodos de un paisaje (Fahrig 2020); especialmente cuando hay una matriz favorable que aumente la conectividad entre fragmentos (Barnes et al. 2014).
En Colombia el área de influencia de la hidroeléctrica Porce II, en el nororiente del departamento de Antioquia, es ideal para este tipo de estudios. Se pueden encontrar fragmentos de bosque de diferente tamaño y forma, su cercanía, fácil acceso y similar altitud permiten hacer comparaciones significativas. El objetivo de este trabajo fue identificar la fauna de escarabajos coprófagos en fragmentos de bosque de la región de Porce II y evaluar el efecto del tamaño de los fragmentos sobre la abundancia, biomasa, estructura, riqueza, diversidad y composición de los ensambles de este grupo de insectos.
MATERIALES Y MÉTODOS
Área de estudio
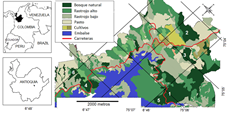
El área de influencia de la central hidroeléctrica Porce II (Fig. 1), se ubica al noreste del departamento de Antioquia, Colombia, en jurisdicción de los municipios de Amalfi, Yolombó y Gómez Plata (EPM 1995). El principal vertedor del embalse es el río Porce, que en sus primeros 60 kilómetros es llamado el río Medellín (EPM 1995). La temperatura promedio anual de la zona es de 23 °C, una humedad relativa del 81,9 % y una precipitación promedio anual de 1971 mm (Bernal y Sandoval 2017). De acuerdo con la clasificación de zonas de vida propuesta por Holdridge (1982), en la región de Porce se encuentran tres zonas: Bosque húmedo tropical (Bh-T), Bosque muy húmedo premontano (Bmh-P) y bosque húmedo montano bajo (Bh-Mb). La cobertura vegetal está compuesta en su mayoría por pastizales para ganadería, rastrojos bajos, rastrojos altos y fragmentos de bosques con diferentes grados de intervención (Cano 1995).

Figura 1: Mapa de la zona de influencia de la central hidroeléctrica Porce II en el departamento de Antioquia. Modificado del mapa de cobertura vegetal (Universidad Nacional de Colombia 2006). Fragmentos de bosque: 1. Normandia. 231 ha; 2. El Pital 69,7 ha; 3. San Ignacio 29,5 ha; 4. La frijolera 32,5 ha; 5. Acacias 29,5 ha; 6. La Picardia 13,4 ha.
Tabla 1: Caracterización de los fragmentos de bosque en el área de influencia de la Central Hidroeléctrica Porce II donde se hizo el muestreo.

Selección de los sitios de muestreo. Con base en el mapa de coberturas vegetales de la zona (Universidad Nacional de Colombia 2006), se seleccionaron seis fragmentos de bosque con tamaños entre los 13,4 y 231 ha, en un intervalo altitudinal entre los 948 y 1 137 m (Fig. 1). Para cada fragmento el tamaño fue estimado usando el programa ArcGIS 9.0 (ESRI c2005) (Tabla 1). La separación entre los fragmentos fue de 30 metros de matriz de pasturas para los fragmentos de 32,5 y 13,4 ha, o en el caso de los pares de fragmentos de 231 ha - 29 ha y 69,7 ha - 29,5 ha la separación se debió a la carretera principal. Tanto las pasturas como las carreteras se consideran una barrera que evita el paso de la mayoría de las especies presentes en el bosque quienes son usualmente incapaces de moverse en zonas abiertas (Montes-Rodríguez et al. 2019).
Muestreo e identificación de los escarabajos coprófagos. Dentro de cada uno de los fragmentos se trazó un transecto de 150 m, en orientación perpendicular al borde del bosque y se instalaron cinco trampas separadas 30 m del borde y entre sí, de este modo se espera que las trampas no estén influenciadas por el efecto de borde el cual fue determinado previamente en menos de 30 m (Montes-Rodríguez et al. 2019). Se realizaron tres salidas de campo, en los meses de noviembre del 2008 (época lluviosa), febrero (época seca) y mayo (época de transición climática) de 2009, cubriendo la variación del régimen de lluvias de la zona. Cada trampa consistió en un vaso de doce onzas enterrado al nivel del suelo y lleno hasta un tercio con agua jabonosa, el excremento humano utilizado como cebo se colocó en una copa plástica de una onza sostenida en el centro de la trampa con un alambre. La captura de insectos se realizó 48 horas después de su instalación. Los especímenes recolectados se guardaron en frascos plásticos con alcohol al 70 % y se transportaron al laboratorio de entomología de la Universidad Nacional sede Medellín, donde se realizaron las labores de curaduría.

Figura 2: Análisis de cobertura de muestreo entre fragmentos de bosque de diferente tamaño, en el área de influencia de la Hidroeléctrica Porce II. Las áreas sombreadas indican intervalos de confianza del 95 %.
La identificación taxonómica se realizó utilizando claves para géneros y la literatura taxonómica de escarabajos americanos referenciada por Vaz-de-Mello et al. (2011). Las especies se identificaron utilizando claves taxonómicas, diagnosis, descripciones y redescripciones para los géneros Deltochilum Eschscholtz, 1822 (González-A et al. 2009, González-Alvarado y Vaz-de-Mello 2014), Dichotomius Hope, 1838 (Sarmiento-G y Amat-G 2014, Montoya-Molina y Vaz-de-Mello 2021), Eurysternus Dalman, 1824 (Génier 2009), Ontherus Erichson, 1847 (Génier 1996), Oxysternon Castelnau, 1840 (Edmonds y Zidek 2004), Phanaeus MacLeay, 1819 (Edmonds y Zidek 2012), Sulcophanaeus Olsoufieff, 1924 (Edmonds 2000) y Sylvicanthon Halffter & Martinez, 1977 (Cupello y Vaz-de-Mello 2018). En el caso de los géneros Ateuchus Weber, 1801, Canthon Hoffmannsegg, 1817, Canthidium Erichson, 1847, Onthophagus Latreille, 1802 y Uroxys Westwood, 1842, la identificación de especies se realizó mediante la comparación de material con la Colección Entomológica del Instituto Humboldt (IAvH-E) y cuando no fue posible, se separaron morfoespecies con base en la morfología externa y del órgano genital masculino (González-A y Medina 2014, González 2021). Los ejemplares se depositaron en el Museo Entomológico Francisco Luís Gallego (MEFLG), de la Universidad Nacional de Colombia sede Medellín.
Análisis de datos. Para evaluar la eficiencia del muestreo para cada fragmento de bosque se realizó un análisis de la cobertura (Cm) y se obtuvo el déficit de muestreo (Cm - 1). El déficit indica la probabilidad de que una especie previamente no detectada, sea encontrada en la muestra cuando esta aumenta en un individuo (Chao y Jost 2012). Los cálculos se realizaron con el paquete “iNEXT” (Chao et al. 2014, Hsieh et al. c2020), en RStudio versión 1.4.1103 (RStudio Team c2020). La diversidad alfa fue estimada en términos de especies efectivas (qD), basados en los números de la serie de Hill, teniendo en cuenta las siguientes consideraciones: El exponente q determina la sensibilidad del índice D a la abundancia relativa de las especies, cuando q=0 es insensible a la abundancia de las especies y equivale a la riqueza observada; en q=1 todas las especies son incluidas con un peso exactamente proporcional a su abundancia y equivale al exponente del índice de equidad de Shannon–Wiener Exp (H’) y cuando q=2 toma en cuenta a las especies más comunes y equivale al índice de dominancia de Simpson (1/Simpson). La comparación de la riqueza alfa y diversidad se realizó con base en la menor cobertura de muestreo, indicando diferencias significativas cuando los intervalos de confianza no se solapan (Moreno et al. 2011, Chao et al. 2014). Los análisis se realizaron con el paquete “iNEXT”, en RStudio versión 1.4.1103 (RStudio Team c2020).

Figura 3: Abundancia y biomasa de Scarabaeinae entre fragmentos de bosque de diferente tamaño, en el área de influencia de la hidroeléctrica Porce II. Letras diferentes indican diferencias significativas entre fragmentos de bosque (Wilcoxon P < 0,05).

Figura 4: Curvas logarítmicas de abundancia en seis fragmentos de bosque en el área de influencia de la hidroeléctrica Porce II. O2= Onthophagus sp2. O3= Onthophagus sp3. C5= Canthidium sp5. C6= Canthidium sp6. C7= Canthidium sp7. AC= Ateuchus candezei. SA= Sylvicanthon aequinoctialis. CS= Canthon subhyalinus. UP= Uroxys aff. pauliani. DA= Dichotomius andresi.
La abundancia se consideró como el número de individuos total y promedio por fragmento. La biomasa se obtuvo pesando de dos a 30 ejemplares de cada especie según su abundancia, ellos se secaron en un horno a 60 °C hasta que alcanzaron un valor constante (Horgan 2005). En el caso de las pocas especies con uno o dos individuos, se decidió no exponerlos y tomar un valor de otra especie con tamaño similar. Posteriormente cada individuo fue pesado en una balanza digital Ohaus modelo AX124 con precisión de 0,0001 g y se obtuvo un valor promedio por especie, este valor se multiplicó por la abundancia para obtener la biomasa total por cada fragmento y por cada muestreo. Se probaron los supuestos de normalidad para la abundancia y biomasa utilizando las pruebas de Shapiro-Wilk y Kolmogorov-Smirnov (con la corrección Lilliefors) del paquete nortest (Gross y Ligges c2015) y la homogeneidad de varianzas con la prueba de Levene del paquete car (Fox y Weisberg c2019) ambos en RStudio versión 1.4.1103 (RStudio Team c2020). Los valores de abundancia y biomasa se compararon con una prueba de Kruskal Wallis y posteriormente una prueba pareada de Wilcoxon para identificar entre cuáles fragmentos se encuentran las diferencias utilizando el paquete Stats (Bolar c2019), en RStudio versión 1.4.1103 (RStudio Team c2020).
Para determinar las especies más afectadas o el aporte individual en abundancia y biomasa a los cambios de todo el ensamble, relacionados con el tamaño del fragmento, se dividieron las especies en cuatro grupos según su biomasa: Livianos, escarabajos con igual a menos de 0,03 gr de peso seco, Intermedios con valores entre 0,031 y 0,12, Pesados con valores entre 0,13 y 0,5 y muy pesados mayores a 0,5. Se determinó si existen diferencias en el valor de biomasa de estos grupos entre fragmentos con el Anova no paramétrico de Kruskal Wallis. También se correlacionaron las abundancias de cada especie con el tamaño del fragmento mediante el coeficiente no paramétrico de Spearman, en este caso se tomó cada muestra como una repetición. Adicionalmente, se elaboraron curvas logarítmicas de abundancia para determinar las especies más abundantes y cambios en la estructura del ensamble entre fragmentos de bosque (Magurran 2004).
La composición de los ensambles entre fragmentos se comparó mediante un análisis de escalamiento multidimensional no métrico (nMDS), con base en el índice de disimilitud de Bray Curtis (Clarke 1993). Teniendo en cuenta la posible dependencia espacial por la distancia entre trampas por fragmento, las salidas de campo se tomaron como réplicas para el ordenamiento. Se calculó el grado de estrés para el nMDS como indicador de confiabilidad del resultado: Valores mayores que 0,05 ofrecen una excelente representación, valores mayores a 0,1 corresponden a una buena ordenación sin riesgo de obtener inferencias falsas y valores mayores a 0,2 pueden conducir a un ordenamiento útil, aunque existe la posibilidad de inducir a error (Clarke 1993). El análisis de similitud (ANOSIM) complementa la información del nMDS con un valor de prueba estadístico R, que define si las diferencias entre las agrupaciones son significativas. Los análisis de nMDS y ANOSIM se realizaron con el paquete vegan (Oksanen et al. c2020) en RStudio versión 1.4.1103 (RStudio Team c2020).
RESULTADOS
Composición de la muestra y cobertura de muestreo: Se recolectaron en total 7123 individuos pertenecientes a quince géneros y 39 especies. Las cinco más abundantes fueron: Onthophagus sp. 2 (N= 1571, 22 %), Onthophagus sp. 3 (N= 1230, 17,2 %), Canthidium sp. 6 (N= 786, 11 %), Canthidium sp. 7 (N= 719, 10 %) y Sylvicanthon aequinoctialis Harold, 1868 (N= 488, 6,8 %) para un 67 % de la abundancia total, mientras que quince especies tuvieron igual o menos de diez individuos (Tabla 2). El estimador de cobertura indicó una alta eficiencia en el muestreo, sus valores superaron el 99 % en todos los fragmentos, siendo más alto en los fragmentos de 13,4 y 69,7 ha (99,8 %) y menor en el fragmento de 29,5 ha (99,2 %). El déficit de muestreo presentó valores entre 0,2 y 0,8 %. Estos resultados implican que las muestras son adecuadas para ser utilizadas con fines comparativos (Fig. 2).

Figura 5: Biomasa de escarabajos en seis fragmentos de bosque de diferente tamaño en la zona de influencia de la hidroeléctrica Porce II. LIV= Livianos, escarabajos con igual a menos de 0,03 gr peso seco. INT= Intermedios, entre 0,031 y 0,12 gr. PE= Pesados, entre 0,13 y 0,5 gr y MPE = muy pesados, mayores a 0,5 gr. letras diferentes indican diferencia significativa (P < 0,05).
Abundancia y biomasa de Scarabaeinae entre los fragmentos de bosque: Se observaron diferencias significativas en el promedio de la abundancia; los fragmentos de 32,5 y 69,7 ha tienen el 22 % del total de individuos cada uno (Fig. 3), seguido por el fragmento de 231 ha con el 16 %, y por último los fragmentos de tamaño igual o inferior a 29,5 ha, tienen menos del 14 % cada uno. La biomasa está relacionada con el tamaño del fragmento. Su mayor valor se obtuvo en el fragmento de 231 ha con el 22 % de la biomasa total, seguido por los fragmentos de 32,5 ha con el 20 % y 69,7 ha con el 18 %, mientras los fragmentos más pequeños, con 29 y 13,4 ha, tuvieron menor biomasa (Fig. 3). Las curvas logarítmicas de abundancia nos muestran que las especies dominantes en cada fragmento son diferentes y pertenecen principalmente a los géneros de escarabajos pequeños como Canthidium y Onthophagus (Fig. 4). Se encontraron diez especies cuya abundancia se correlaciona con el tamaño del fragmento, Canthidium sp7 (ρ=0,29; P=0,005); Canthon angustatus Harold, 1867 (ρ=0,24; P=0,019), Canthon subhyalinus Harold, 1867 (ρ=0,25; P=0,015), Onthophagus acuminatus Harold, 1880 (ρ=0,29; P=0,005), Uroxys micros Bates, 1887 (ρ=0,22; P=0,03) y Uroxys sp2 (ρ=0,23; P=0,025), dos de tamaño intermedio como Sylvicanthon aequinoctialis (ρ=0,25; P=0,017) y Eurysternus caribaeus (Herbst, 1789) (ρ=0,30; P=0,0031) y las dos especies con mayor biomasa Oxysternon conspicillatum (Weber, 1880) (ρ=0,26; P=0,012) y Deltochilum panamensis Howden, 1966 (ρ=0,35; P=0,0007). Los escarabajos medianos y los muy pesados tienen mayor biomasa en el fragmento de mayor tamaño (Fig. 5).

Figura 6: Riqueza y diversidad de Scarabaeinae entre fragmentos de bosque de diferente tamaño en el área de influencia de la Hidroeléctrica Porce II. Las barras indican intervalos de confianza del 95 %.
Tabla 2: Abundancia por especie de los escarabajos coprófagos Scarabaeinae recolectados en seis fragmentos de bosque de la zona de influencia de la hidroeléctrica Porce II.


Diversidad en orden qD entre fragmentos: La mayor riqueza se obtuvo en los fragmentos de 231 y 29,5 ha con 28 y 27 especies, seguidos por el fragmento de 29 ha con 23 especies y los demás fragmentos con menos de 20 especies (Fig. 6). En un mismo nivel de cobertura (99,2 %), no existen diferencias significativas entre la riqueza de los fragmentos de 29,5 y 231 ha, pero sí de estos fragmentos con los demás (Fig. 6).
La diversidad en orden q1 obtuvo su valor más alto en el fragmento de 231 ha con once especies efectivas y es significativamente mayor que en los fragmentos de 29 ha con nueve especies y los fragmentos 13,4, 32,5 y 29,5 ha con ocho especies efectivas cada uno. El fragmento de 69,7 ha fue significativamente menos diverso que los demás (Fig. 6). La diversidad en orden q2 presentó un patrón similar a q1, el fragmento de 231 ha es el más diverso y significativamente diferente a los fragmentos de 13,4, 29 y 32,5 ha. Los fragmentos de 69,7 y 29,5 ha fueron significativamente menos diversos que los demás, sin diferencias entre sí (Fig. 6).
Composición de los ensambles entre fragmentos: El análisis NMDS reveló diferencias en la composición de los ensambles de Scarabaeinae entre los fragmentos de bosque, que se evidenció en la ausencia de superposición entre los polígonos. Excepto en los fragmentos de 13,4 y 32,5 ha, indicando que fueron más cercanos entre sí (Fig. 7). Las agrupaciones fueron diferentes (ANOSIM global R= 0,78, P < 0,0001).
DISCUSIÓN
La deforestación y posterior actividad agropecuaria trae como consecuencia la transformación de un bosque grande original en numerosos fragmentos aislados de diferente tamaño (Santos y Telleria 2006). Se espera que especies sensibles de escarabajos coprófagos disminuyan sus poblaciones en fragmentos lo suficientemente reducidos y por otra parte aumenten las especies con mayor capacidad de adaptación, cambiando la composición y estructura del ensamble (Nichols et al. 2007). En la región de influencia de la hidroeléctrica Porce II se encontró que las diferencias en la riqueza de escarabajos coprófagos no tienen relación con el tamaño del fragmento de bosque, este resultado es similar a lo recopilado por Radtke et al. (2008) quienes revisaron trabajos relacionados en la Amazonía. Aunque no hay diferencia en la riqueza, si se encontró que el fragmento más grande tiene mayor diversidad, lo cual coincide con varios trabajos que han abordado el tema (Klein 1989, Nichols et al. 2007, Rodríguez-García et al. 2021).
Un factor que afecta significativamente la diversidad del ensamble de escarabajos coprófagos en bosques es la disponibilidad de recurso alimenticio el cual está constituido principalmente por excremento de mamíferos (Peck y Forsyth 1982, Estrada et al. 1999). A su vez el tamaño del fragmento afecta directamente la abundancia y riqueza de mamíferos (Vieira et al. 2009, Lawrence et al. 2018); y las probabilidades de supervivencia de sus poblaciones en el largo plazo (Schwarzkopf y Rylands 1989, Mandujano y Escobedo-Morales 2008). De esta manera, se espera que, al disminuir el tamaño del fragmento, disminuya la disponibilidad de excremento en cantidad y calidad para los escarabajos, lo cual pone en riesgo la viabilidad de algunas poblaciones de escarabajos (Estrada et al. 1999, Larsen et al. 2006). Aunque son pocos los trabajos que han evaluado simultáneamente las comunidades de mamíferos y escarabajos coprófagos, en términos generales, las poblaciones de escarabajos tienden a decrecer cuando disminuyen las de mamíferos (Raine y Slade 2019, Rodríguez-García et al. 2021).

Figura 7: Escalamiento multidimensional no métrico de la disimilitud en la composición de los ensambles de Scarabaeinae entre fragmentos de bosque de diferente tamaño, en el área de influencia de la Hidroeléctrica Porce II.
Las dos especies con mayor biomasa Oxysternon conspicillatum (Weber, 1880) y Deltochilum gibossum Howden, 1966, aportan el 31,8 % de la biomasa total con lo cual tienen una influencia grande en los cambios en biomasa de todo el ensamble y son sensibles a la reducción del fragmento de bosque. De manera similar, trabajos donde se ha medido el tamaño en lugar de la biomasa, han reportado escarabajos más pequeños en fragmentos en comparación al bosque grande o continuo (Klein 1989, Nichols et al. 2007, Radtke et al. 2008). Probablemente solo los mamíferos medianos y grandes con capacidad de producir mayores cantidades de excremento pueden sostener a los escarabajos de mayor tamaño que son atraídos principalmente por porciones grandes de excremento, mayores a 200 g (Peck y Howden 1984). Son precisamente estos mamíferos los más sensibles a los efectos nocivos de la fragmentación incluyendo la reducción del hábitat (Estrada y Coates-Estrada 2002, Lawrence et al. 2018, López-Arévalo et al. 2021).
Cuanto mayor es el tamaño del fragmento, mayor es la biomasa total de escarabajos, mientras que la abundancia fue mayor en los fragmentos de tamaño medio; resultados similares a los reportados por Radtke et al. (2008) y Andresen (2003), respectivamente. Desde el punto de vista funcional esto es relevante debido a que la biomasa se correlaciona mejor que la riqueza de especies y la abundancia, con las funciones ecológicas que realizan los escarabajos, como la dispersión secundaria de semillas (Vulinec 2002), el procesamiento de excremento (Klein 1989, Horgan 2005) y el control de las poblaciones de moscas hematófagas y parásitos vectores (Klein 1989, Andresen 2003, Nichols et al. 2008).
Por otra parte, Amézquita y Favila (2010) no encontraron relación entre el tamaño del fragmento y la cantidad de excremento removido por escarabajos, ellos plantean que hay una aparente compensación de un mayor número de individuos de escarabajos de menor tamaño con respecto a un menor número de los de mayor tamaño. Aunque el total de excremento procesado por escarabajos pequeños y grandes fuera igual, funcionalmente los escarabajos grandes son mejores dispersores secundarios de las semillas grandes y su ausencia en fragmentos pequeños puede afectar la capacidad de regeneración del bosque (Andresen 2003).
Un factor determinante en los resultados en este tipo de trabajos es la escala de tamaño de los fragmentos a la cual se evalúan los cambios en la composición y estructura de escarabajos coprófagos. En este estudio, en fragmentos entre 13,4 y 69,7 ha, no aumentan progresivamente la diversidad y biomasa de escarabajos con el tamaño, mientras que el fragmento más grande (231 ha) si se diferencia claramente de los demás, lo cual concuerda con estudios que muestran que solo cuando fragmentos de tamaños mayores de 100 ha son tenidos en cuenta, los cambios en la diversidad, como en el número de especies comunes y exclusivas, son notablemente detectadas (Klein 1989). Igualmente, Estrada y Coates-Estrada (2002) encontraron diferencias en la diversidad de escarabajos coprófagos entre el bosque grande o continuo (700 ha) y fragmentos de bosque (entre 1- 80 ha). Esta tendencia también ha sido evidente en mamíferos. Por ejemplo, Vieira et al. (2009) encontraron efecto del tamaño del fragmento en la riqueza de pequeños mamíferos, solo cuando se tuvieron en cuenta fragmentos mayores a 10 000 ha. Aunque la importancia de los fragmentos o reservas naturales grandes es alta, el papel de las reservas pequeñas no se debe subestimar (Shafer 1995, López-Arévalo et al. 2021, Rodríguez-García et al. 2021). En este estudio, todos los fragmentos de bosque aportaron a la riqueza regional de especies de escarabajos y todos deberían ser considerados importantes en la conservación de la biodiversidad regional. Desde este punto de vista, la conectividad entre fragmentos es una prioridad para disminuir el aislamiento que genera la pérdida de especies en el mediano y largo plazo (Escobar et al. 2008).
A pesar de su relativa cercanía, los fragmentos muestran diferente composición de escarabajos, lo que demuestra que las matrices de pasturas y regeneraciones tempranas son una barrera fuerte para el movimiento y conectividad de la mayoría de las especies de escarabajos (Montes-Rodríguez et al. 2019). La adquisición de las tierras como parte de la compensación ambiental de empresas públicas de Medellín EEPPM por la construcción de la hidroeléctrica Porce II ha dejado expuestas una parte del territorio a un proceso de restauración ecológica natural y progresivo aumento de la sucesión vegetal (Marín et al. 2008). Se espera que a medida que aumente la regeneración natural, cada vez más especies de escarabajos puedan moverse por la matriz (Montes-Rodríguez et al. 2019) y se aumente la conectividad y el flujo genético entre fragmentos y las probabilidades de conservación a largo plazo de especies restringidas al bosque (Escobar et al. 2008).
Además de la calidad de la matriz, otro factor que aparentemente influye en la conectividad es la distancia entre fragmentos; fragmentos separados por cerca de 50 m en su punto más cercano muestran más similitud en la composición de escarabajos que los fragmentos aislados por una distancia cercana a 200 m (Fig. 5). Al evaluar el efecto de borde en la región de Porce II, Montes-Rodríguez et al. (2019) encontraron que algunas especies principalmente adaptadas al bosque pueden desplazarse al menos 30 m en la matriz de pasturas y su abundancia disminuye al aumentar la distancia al borde del bosque, por lo cual, si los fragmentos están lo suficientemente cercanos, estos escarabajos podrían pasar de un fragmento a otro a pesar de las dificultades que implica moverse por la matriz.
Por último, es importante tener en cuenta que la distancia entre trampas fue de 30 metros, mientras la distancia mínima establecida para que las muestras sean independientes es de 50 m (Larsen y Forsyth 2005). La falta de independencia entre trampas podría afectar los resultados y análisis de este trabajo, por lo que las conclusiones de este trabajo deben ser tomadas con precaución.
AGRADECIMIENTOS
Los autores expresan su agradecimiento a la Universidad Nacional sede Medellín por la financiación de este proyecto. A las Empresas Públicas de Medellín por su apoyo logístico en las salidas de campo. A los evaluadores por sus recomendaciones y sugerencias.
Literatura citada
Referencias
Amézquita SJ, Forsyth A, Lopera A, Camacho A. 1999. Comparación de la composición y riqueza de especies de escarabajos coprófagos (Coleoptera: Scarabaeidae) en remanentes de bosque de la Orinoquía Colombiana. Acta Zool. Mex. (n.s.) 76:113-126. doi: https://doi.org/10.21829/azm.1999.76761703
Amézquita S, Favila ME. 2010. Removal Rates of native and exotic dung by dung beetles (Scarabaeidae: Scarabaeinae) in a fragmented tropical rain forest. Environ. Entomol. 39(2):328-336. doi: https://doi.org/10.1603/EN09182
Andresen E. 2003. Effect of forest fragmentation on dung beetle communities and functional consequences for plant regeneration. Ecography 26(1):87–97. doi: https://doi.org/10.1034/j.1600-0587.2003.03362.x
Armenteras D, Gast F, Villareal H. 2003. Andean forest fragmentation and the representativeness of protected natural areas in the eastern Andes, Colombia. Biol. Conserv. 113(2):245–256. doi: https://doi.org/10.1016/S0006-3207(02)00359-2
Barnes AD, Emberson RM, Chapman HM, Krell FT, Didham RK. 2014. Matrix habitat restoration alters dung beetle species responses across tropical forest edges. Biol. Conserv. 170:28-37. doi: https://doi.org/10.1016/j.biocon.2013.12.006
Bernal C, Sandoval DC. 2017. Evaluación de la precipitación como indicador de la alteración del mesoclima en las zonas de influencia de los embalses Miel I y Porce II. [Tesis]. [Bogotá]: Universidad de la Salle.
Bolar K. c2019. The STATS package. Interactive Document for Working with Basic Statistical Analysis. [Revisada en: 1 Ago 2020]. https://cran.r-project.org/web/packages/STAT/STAT.pdf
Cano JA. 1995. Utilización de un sistema de información geográfico (SIG) en la elaboración de un plan de uso y manejo de coberturas vegetales. [Tesis]. [Medellín]: Universidad Nacional de Colombia.
Chao A, Gotelli NJ, Hsieh TC, Sander EL, Ma KH, Colwell RK, Ellison AM. 2014. Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol. Monogr. 84(1):45–67. doi: https://doi.org/10.1890/13-0133.1
Chao A, Jost L. 2012. Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology 93(12):2533–2547. doi: https://doi.org/10.1890/11-1952.1
Clarke KR. 1993. Non-parametric multivariate analysis of changes in community structure. Austral. J. Ecol. 18(1):117-143. doi: https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Cupello M, Vaz-De-Mello FZ. 2018. A monographic revision of the Neotropical dung beetle genus of Sylvicanthon Halffter & Martínez (Coloptera: Scarabaeidae: Scarabaeinae: Deltochilini), including a reappraisal of the taxonomic history of ‘Canthon sensu lato’. Eur. J. Taxon. 467:1-205. doi: https://doi.org/10.5852/ejt.2018.467.
Edmonds WD. 2000. Revision of the Neotropical dung beetle genus Sulcophanaeus. Folia Heyrovskyana Supplementum 6:1–60.
Edmonds WD, Zídek J. 2004. Revision of the neotropical dung beetle genus Oxysternon (Coleoptera: Scarabaeidae: Scarabaeinae). Folia Heyrovskyana Supplementum 11:1–58.
Edmonds WD, Zidek J. 2012. Taxonomy of Phanaeus revisited: Revised keys to and comments on species of the New World dung beetle genus Phanaeus MacLeay, 1819 (Coleoptera: Scarabaeidae: Scarabaeinae: Phanaeini). Insecta Mundi 0274:1–108.
[EPM] Empresas Públicas de Medellín. 1995. Proyecto hidroeléctrico Porce II. Medellín: Oficina de comunicaciones empresas públicas de Medellín.
Escobar F, Halffter G, Solís A, Halffter V, Navarrete D. 2008. Temporal shifts in dung beetle community structure within a protected area of tropical wet forest: a 35-year study and its implications for long-term conservation. J. Appl. Ecol. 45(6):1584–1592. doi: https://doi.org/10.1111/j.1365-2664.2008.01551.x
ESRI. c2005. Arc Gis versión 9.0. GIS and Mapping Software. [Revisada en: 6 Feb 2009]. http://www.esri.com/software/arcgis/
Estrada A, Anzures D, Coates-Estrada R. 1999. Tropical rain forest fragmentation, howler monkeys (Alouatta palliata), and dung beetles at Los Tuxtlas, Mexico. Am. J. Primatol. 48(4):253–262. doi: https://doi.org/10.1002/(SICI)1098-2345(1999)48:4<253::AID-AJP1>3.0.CO;2-D
Estrada A, Coates-Estrada R. 2002. Dung beetles in continuous forest fragments and agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodivers. Conserv. 11:1903-1919. doi: https://doi.org/10.1023/A:1020896928578
Fahrig L. 2020. Why do several small patches hold more species than few large patches? Global Ecol. Biogeogr. 29(4):615-628. doi: https://doi.org/10.1111/geb.13059
Fox J, Weisberg S. c2019. An R Companion to Applied Regression, Third edition. Sage, Thousand Oaks CA. R package version 3.1-0. [Revisada en: 1 Ago 2020]. https://cran.r-project.org/web/packages/car/car.pdf
Génier F. 1996. A revision of the Neotropical genus Ontherus Erichson (Coleoptera: Scarabaeidae, Scarabaeinae). Mem. Ent. Soc. Can. 128(170):1–169. doi: https://doi.org/10.4039/entm128170fv
Génier F. 2009. Le genre Eurysternus Dalman, 1824 (Scarabaeidae: Scarabaeinae: Oniticellini), révision taxonomique et clés de détermination illustrées. Sofia, Bulgaria: Pensoft Series Faunistica.
Gill BD. 1991. Capitulo 12 Dung beetles in tropical American forests. En: Hanski I, Cambefort Y, editores. The Ecology of Dung Beetles. New Jersey: Princeton University Press. p. 211-229.
González-A FA, Molano F, Medina CA. 2009. Los subgéneros Calhyboma, Hybomidium y Telhyboma (Coleoptera: Scarabaeidae: Scarabaeinae: Deltochilum). Rev. Colomb. Entomol. 35(2):253–274. doi: https://doi.org/10.25100/socolen.v35i2.9228
González-A FA, Medina CA. 2014. Escarabajos coprófagos de la subfamilia Scarabaeinae. En: Pizano C, García H, editores. El bosque seco tropical en Colombia. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH). p. 195-213.
González-Alvarado A, Vaz-de-Mello FZ. 2014. Taxonomic review of the subgenus Hybomidium Shipp 1897 (Coleoptera: Scarabaeidae: Scarabaeinae: Deltochilum). Ann. Soc. Entomol. Fr. 50(3-4):431–476. doi: https://doi.org/10.1080/00379271.2014.989178
González A. 2021. Escarabajos coprófagos (Coleoptera: Scarabaeidae: Scarabaeinae) de bosques secos colombianos en la Colección Entomológica del Instituto Alexander von Humboldt. Biota Colomb. 16(1):88–95. doi: https://www.doi.org/10.15468/hdfuql
Gross J. Ligges U. c2015. Nortest: Tests for Normality. Five omnibus tests for testing the composite hypothesis of normality. R package version 1.0-4. [Revisada en: 1 Ago 2020] https://CRAN.R-project.org/package=nortest
Halffter G, Arellano L. 2002. Response of dung beetle diversity to human-induced changes in a tropical landscape. Biotropica 34(1):144–154. doi: https://doi.org/10.1111/j.1744-7429.2002.tb00250.x
Hsieh TC, Ma KH, Chao A. c2020. iNEXT: Interpolation and Extrapolation for Species Diversity. R package version 2.0.20. [Revisada en: 1 Ago 2020]. https://cran.r-project.org/web/packages/iNEXT/iNEXT.pdf
Holdridge LR. 1982. Ecología basada en zonas de vida. San José de Costa Rica: Instituto Interamericano de Cooperación de la Agricultura.
Horgan FG. 2005. Effects of deforestation on diversity, biomass, and function of dung beetles on the eastern slopes of peruvian Andes. Forest. Ecol. Manag. 216(1-3):117-133. doi: https://doi.org/10.1016/j.foreco.2005.05.049
Klein BC. 1989. Effects of forest fragmentation on dung and carrion beetle communities in central Amazonia. Ecology 70(6):1715-1725. doi: https://doi.org/10.2307/1938106
Lawrence A, O’Connor K, Haroutounian V, Swei A. 2018. Patterns of diversity along a habitat size gradient in a biodiversity hotspot. Ecosphere 9(4):e02183. doi: https://doi.org/10.1002/ecs2.2183
Larsen TH, Forsyth A. 2005. Trap Spacing and Transect Design for Dung Beetle Biodiversity Studies. Biotropica 37(2):322–325. doi: https://dx.doi.org/10.1111/j.1744-7429.2005.00042.x
Larsen TH, Lopera A, Forsyth A. 2006. Extreme trophic and habitat specialization by peruvian dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopts. Bull. 60(4):315-324. doi: https://doi.org/10.1649/0010-065X(2006)60[315:ETAHSB]2.0.CO;2
López-Arévalo HF, Liévano-Latorre LF, Montenegro OL. 2021. El papel de las pequeñas reservas en la conservación de mamíferos en Colombia. Caldasia 43(2):354–365. doi: https://doi.org/10.15446/caldasia.v43n1.85466
Magurran A. 2004. Measuring biological diversity. Malden: Blackwell Science Ltd. Chapter 2, The commonness, and rarity, of species; p.18−71.
Mandujano S, Escobedo-Morales LA. 2008. Population viability analysis of howler monkey (Alouatta palliata mexicana) in a highly fragmented landscape in Los Tuxtlas, Mexico. Trop. Conserv. Sci. 1(1):43-62. doi: https://doi.org/10.1177/194008290800100104
Marín AL, Álvarez CF, Uribe SI, Morales M. 2008. Dinámica temporal del patrón del paisaje en el área de la hidroeléctrica Porce II, (Antioquia, Colombia) de 1961 al 2001. Bol. Cienc. Tierra 23:33–42.
[MinAmbiente] Ministerio de Ambiente y Desarrollo Sostenible. c2017. Resultados monitoreo de la deforestación. [Revisada en: 1 Ago 2020] http://documentacion.ideam.gov.co/openbiblio/bvirtual/023835/Resultados_Monitoreo_Deforestacion_2017.pdf
Montes-Rodríguez JM, Ortega Molina OE, Espinosa Vélez Y. 2019. Efecto de borde en ensambles de escarabajos coprófagos (Coleoptera: Scarabaeidae) en fragmentos de bosque de un paisaje andino. Caldasia 41(2):380–391. doi: https://doi.org/10.15446/caldasia.v41n2.70845
Montoya-Molina S. Vaz-de-Mello FZ. 2021. Taxonomic review of the Dichotomius (Luederwaldtinia) agenor species group (Coleoptera: Scarabaeidae: Scarabaeinae). Eur. J. Taxon. 734(1):1-64. doi: https://doi.org/10.5852/ejt.2021.734.1233
Moreno CE, Barragán F, Pineda E, Pavón NP. 2011. Reanálisis de la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Rev. mex. Biodivers. 82(4):1249-1261. doi: https://doi.org/10.22201/ib.20078706e.2011.4.745
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature 403:1-72. doi: https://doi.org/10.1038/35002501
Nichols E, Larsen T, Spector S, Davis AL, Escobar F, Fávila M, Vulinec K.2007. Global dung beetle response to tropical forest modification and fragmentation: A quantitative literature review and meta-analysis. Biol. Conserv. 137(1):1–19. doi: https://doi.org/10.1016/j.biocon.2007.01.023
Nichols E, Spector S, Louzada J, Larsen T, Amézquita S, Fávila ME. 2008. Ecological functions and ecosystems services provided by Scarabaeinae dung beetles. Biol. Conserv. 141(6):1461–1474. doi: https://doi.org/10.1016/j.biocon.2008.04.011
Noriega JA, March-Salas M, Castillo S, García-Q H, Hortal J, Santos AMC. 2021. Human perturbations reduce dung beetle diversity and dung removal ecosystem function. Biotropica 53(3):753-766. doi: https://doi.org/10.1111/btp.12953
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H. c2020. vegan: Community Ecology Package. R package version 2.5-7. [Revisada en: 1 Ago 2020]. https://CRAN.R-project.org/package=vegan
Peck BS, Forsyth A. 1982. Composition, structure, and competitive behaviour in a guild of ecuadorian rain forest dung beetles (Coleoptera: Scarabaeidae). Can. J. Zool. 60(7):1624-1634. doi: https://doi.org/10.1139/z82-213
Peck SB, Howden HF. 1984. Response of a dung beetle guild to different size of dung bait in a panamian rainforest. Biotropica 16(3):235-238. doi: https://doi.org/10.2307/2388057
Primack R, Rozzi R, Feisinger P. 2001. Diseño de áreas protegidas. En: Primack R, Rozzi R, Feisenger P, Massardo F, Dirzo R. Fundamentos de Conservación Biológica. Perspectivas latinoamericanas. México D. F.: Fondo de Cultura Económica. p. 480-484.
Radtke MG, da Fonseca CRV, Williamson GB. 2008. Forest fragment size effects on dung beetle communities? Biol. Conserv. 141(3):613-614. doi: https://doi.org/10.1016/j.biocon.2007.11.010
Raine EH, Slade EM. 2019. Dung beetle–mammal associations: methods, research trends and future directions. Proc. R. Soc. B 286. doi: https://doi.org/10.1098/rspb.2018.2002
Rodríguez-García S, Carretero-Pinzón X, Monroy D, Medellín F, García M, Noriega JA. 2021. Influencia de la fragmentación de bosques de galería en la Orinoquia Colombiana sobre la diversidad y estructura funcional del ensamblaje de escarabajos coprófagos. Bol. Cient. Mus. Hist. Nat. 25(2):119-137. doi: https://doi.org/10.17151/bccm.2021.25.2.8
RStudio Team. c2020. RStudio: integrated development for R. Boston: RStudio, Inc. [Revisada en: 1 Ago 2020]. http://www.rstudio.com/
Santos T, Tellería JL. 2006. Pérdida y fragmentación del hábitat: efecto sobre la conservación de las especies. Ecosistemas 15(2):3-12.
Sarmiento-G R, Amat-G G. 2014. Escarabajos del género Dichotomius Hope 1838 (Scarabaeidae: Scarabaeinae) en Colombia. Fauna de Colombia. Monografia 4. Bogotá: Universidad Nacional de Colombia.
Shafer CL. 1995. Values and shortcomings of small reserves. BioScience 45(2):80-88. doi: https://doi.org/10.2307/1312609
Spector S, Ayzama S. 2003. Rapid turnover and edge effects in dung beetle assemblages (Scarabaeidae) at a Bolivian Neotropical Forest–Savanna Ecotone. Biotropica 35(3):394-404. doi: https://doi.org/10.1111/j.1744-7429.2003.tb00593.x
Spector S. 2006. Scarabaeinae dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) an invertebrate focal taxon for biodiversity research and conservation. Coleopts. Bull. 60(5):71–83. doi: http://dx.doi.org/10.1649/0010-065X(2006)60%5B71:SDBCSS%5D2.0.CO;2
Schwarzkopf L, Rylands AB. 1989. Primate species richness in relation to habitat structure in amazonian rainforest fragments. Biol. Conserv. 48:1-12. doi: https://doi.org/10.1016/0006-3207(89)90055-4
Universidad Nacional de Colombia. 2006. Mapa de Coberturas Vegetales. Escala 1: 25.000. Departamento de Ciencias Forestales.
Vasconcelos HL, Bruna EM. 2012. Arthropod responses to the experimental isolation of Amazonian forest fragments. Zoologia 29(6):515–530. doi: https://doi.org/10.1590/S1984-46702012000600003
Vaz-de-Mello FZ, Edmonds WD, Ocampo FC, Schoolmeesters P. 2011. A multilingual key to the genera and subgenera of the subfamily Scarabaeinae of the new world (Coleoptera: Scarabaeidae). Zootaxa 2854(1):1–73. doi: https://doi.org/10.11646/zootaxa.2854.1.1
Vieira MV, Olifiers N, Delciellos AC, Antunes VZ, Bernardo LR, Grelle CEV, Cerqueira R. 2009. Land use vs. fragment size and isolation as determinants of small mammal composition and richness in atlantic forest remnants. Biol. Conserv. 142(6):1191–1200. doi: https://doi.org/10.1016/j.biocon.2009.02.006
Vulinec K. 2002. Dung beetle communities and seed dispersal in primary forest and disturbed land in Amazonia. Biotropica 34(2):297–309. doi: https://doi.org/10.1111/j.1744-7429.2002.tb00541.x
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
Licencia

Esta obra está bajo una licencia internacional Creative Commons Atribución 4.0.
Aquellos autores/as que tengan publicaciones con esta revista, aceptan los términos siguientes:
- Los autores/as conservarán sus derechos de autor y garantizarán a la revista el derecho de primera publicación de su obra, el cual estará simultáneamente sujeto a la Licencia de reconocimiento de Creative Commons que permite a terceros compartir la obra siempre que se indique su autor y su primera publicación esta revista.
- Los autores/as podrán adoptar otros acuerdos de licencia no exclusiva de distribución de la versión de la obra publicada (p. ej.: depositarla en un archivo telemático institucional o publicarla en un volumen monográfico) siempre que se indique la publicación inicial en esta revista.
- Se permite y recomienda a los autores/as difundir su obra a través de Internet (p. ej.: en archivos telemáticos institucionales o en su página web) antes y durante el proceso de envío, lo cual puede producir intercambios interesantes y aumentar las citas de la obra publicada. (Véase El efecto del acceso abierto).















