



Publicado
Análise GGE biplot na avaliação de características de cachos em Açaizeiros da região Amazônica
GGE Biplot Analysis in Evaluations in Bunches Characteristics from Assai Palm Trees of the Amazon Region
GGE biplot análisis en la evaluación de las características de los racimos en los açaizeiros de la región amazónica
DOI:
https://doi.org/10.15446/ma.v8n1.63456Palavras-chave:
Euterpe oleracea, interação genótipos x ambientes, estabilidade, adaptabilidade, GGE Biplot (pt)Euterpe oleracea, interaction genotypes x environments, stability, adaptability, GGE biplot (en)
Euterpe oleracea, la interacción genotipo x ambiente, estabilidad, adaptabilidad, GGE biplot. (es)
O interesse pelos frutos do açaizeiro em outras regiões do país torna necessárias pesquisas de melhoramento genético para a Euterpe oleracea. A pesquisa teve o objetivo de verificar, pelo GGE (efeitos principais dos genótipos e da interação genótipo x ambiente) biplot, aqueles promissores para seleção entre trinta progênies de diferentes plantas matrizes de uma população do município de Anajás e cultivadas em Tomé-Açu, Pará, e também identificar o ambiente que melhor permite a discriminação destes. Foi utilizado o delineamento experimental de blocos casualizados, contendo trinta tratamentos (progênies) e três repetições. Foram avaliadas as seguintes características: peso médio dos cachos (PMC), em gramas; peso total de cachos (PTC), em gramas e; número total de cachos (NTC). Concluiu-se que para fins de seleção baseado no ideótipo, os melhores genótipos em PMC foram G22 e G19; em PTC foram G10 e G14 e, em NTC foi o genótipo G10; os genótipos com valores médios superiores para as características mostram menor estabilidade, excetuando-se apenas G10 para PTC; os GGE biplots permitem visualizar a disposição, o inter-relacionamento com efeitos ambientais e escolher genótipos superiores; na análise gráfica pelo método GGE biplot os efeitos dos ambientes (anos) permitem a diferenciação dos genótipos, com destaque ao ambiente A3 para PMC e PTC e, A1 para NTC; para a representatividade de todos os ambientes é o A1 nas características PMC e PTC; enquanto que é o A2 para NTC.
The interest by assai palms fruits in other regions of the country makes necessary genetic improvement for the Euterpe oleracea. The research aimed to verify, by gge (Genotype main effects + Genotype-by-environment interaction) biplot, those promising for selection among 30 families from different mother plants of a population of the municipality of Anajás and cultivated in Tomé-Açu, Pará, and also identify the environment that best allows the discrimination of these. The experimental design was in randomized blocks, with 30 treatments (progenies) and three repetitions, were evaluated the characteristics: average weight of the bunches (pmc), in grams; total number of clusters (ntc) and; total weight of the bunches (ptc). It was concluded that for purposes of selection based on the ideotype, the best genotypes in pmc were G22 and G19; in ptc are G10 and G14, and ntc was the genotype G10; the genotypes with average values for the evaluated characteristics showed less stability, except G10 for ptc; The gge biplots allow you to view the layout, the inter-relationship with environmental effects and choose superior genotypes; in the graphical analysis by gge biplot method the environments effects (years) allow differentiation of genotypes, with emphasis to the environment A3 for pmc and ptc, and A1 For ntc; for the representativity of all environments is the A1 on the characteristics pmc and ptc; while that is the A2 for ntc.
Análise GGE biplot na avaliação de características de cachos em Açaizeiros da região Amazônica
Yokomizo, G. Ken-Iti, J.T. de Farias Neto, M. do S. Padilha de Oliveira & Kuang Hongyu. (2017). Análise GGE biplot na avaliação de características de cachos em açaizeiros da região amazônica. Mundo Amazónico, 8(1): e63456. Mundo Amazónico 8(1): e63456. https://doi.org/10.15446/ma.v8n1.63456
Artigo de pesquisa. Editor: Juan A. Echeverri
Recebido: 2017-03-22. Devolvido para revisões: 2017-07-24 Aceito: 2017-07-28
Gilberto Ken-Iti Yokomizo1, João Tomé de Farias Neto2, Maria do Socorro Padilha de Oliveira3, Kuang Hongyu41Gilberto Ken-Iti Yokomizo. Engenheiro Agrônomo pela Universidade de São Paulo (1991), mestrado e doutorado em Agronomia (Genética e Melhoramento de Plantas) pela Universidade de São Paulo (1994 e 1999). Pesquisador da Embrapa Amapá. Pesquisador associado do doutorado do Bionorte do MCT. Tem experiência na área de genética vegetal e quantitativa, melhoramento genético, atuando nos temas: soja, citros, palmito de pupunheira, açaizeiro e mangabeiras. Embrapa Amapá, Rod. Juscelino Kubitschek, 2600, Universidade, CEP 68903-419 Macapá-AP. gilberto.yokomizo@embrapa.br
2João Tomé de Farias Neto. Engenheiro Agrônomo pela UFRA (1981), mestrado e doutorado em Agronomia (Genética e Melhoramento de Plantas) pela Universidade de São Paulo (1986 e 1995). Pesquisador da Embrapa Amazonia Oriental. Experiência na área de Agronomia, com ênfase em Melhoramento Vegetal, atuando nos temas: Amazônia, espécie frutífera, euterpe oleracea e genética quantitativa. Embrapa Amazônia Oriental, Travessa Dr. Enéas Pinheiro, s/no, CEP 66095-100 Belém-PA. joao.farias@embrapa.br
3Maria do Socorro Padilha de Oliveira. Engenheira Agrônoma e Florestal pela UFRA (1983 e 1987); mestrado em Botânica pela UFRPE (1995) e doutorado em Genética e Melhoramento de Plantas pela UFLA (2005). Pesquisadora da Embrapa Amazônia Oriental. Professor permanente da Pós-Graduação em Biotecnologia Aplicada à Agropecuária da UFRA. Experiência em recursos genéticos e melhoramento de palmeiras nativas da Amazônia. Embrapa Amazônia Oriental, Travessa Dr. Enéas Pinheiro, s/no, CEP 66095-100 Belém-PA. socorro-padilha.oliveira@embrapa.br
4Kuang Hongyu. Matemático pela UFU, Mestre e Doutor em Estatística e Experimentação Agronômica pela Universidade de São Paulo. Professor da Universidade Federal de Mato Grosso. Tem experiência na área de Estatística Experimental, com ênfase em Análise Multivariada, atuando principalmente nos seguintes temas: Modelos AMMI, GGE biplot, Modelos AMMI-ponderado, GGE biplot-ponderado, Distribuição dos Autovalores, Simulação Multivariada, Biplot e Interação Genótipos x Ambientes. Universidade Federal de Mato Grosso, Av. Fernando Corrêa da Costa, 2367, Boa Esperança, CEP 78060-900 Cuiabá-MT. kuang_hongyu@hotmail.com
Resumo: O interesse pelos frutos do açaizeiro em outras regiões do país torna necessárias pesquisas de melhoramento genético para a Euterpe oleracea. A pesquisa teve o objetivo de verificar, pelo GGE (efeitos principais dos genótipos e da interação genótipo x ambiente) biplot, aqueles promissores para seleção entre trinta progênies de diferentes plantas matrizes de uma população do município de Anajás e cultivadas em Tomé-Açu, Pará, e também identificar o ambiente que melhor permite a discriminação destes. Foi utilizado o delineamento experimental de blocos casualizados, contendo trinta tratamentos (progênies) e três repetições. Foram avaliadas as seguintes características: peso médio dos cachos (PMC), em gramas; peso total de cachos (PTC), em gramas e; número total de cachos (NTC). Concluiu-se que para fins de seleção baseado no ideótipo, os melhores genótipos em PMC foram G22 e G19; em PTC foram G10 e G14 e, em NTC foi o genótipo G10; os genótipos com valores médios superiores para as características mostram menor estabilidade, excetuando-se apenas G10 para PTC; os GGE biplots permitem visualizar a disposição, o inter-relacionamento com efeitos ambientais e escolher genótipos superiores; na análise gráfica pelo método GGE biplot os efeitos dos ambientes (anos) permitem a diferenciação dos genótipos, com destaque ao ambiente A3 para PMC e PTC e, A1 para NTC; para a representatividade de todos os ambientes é o A1 nas características PMC e PTC; enquanto que é o A2 para NTC.
Palavras-chave: Euterpe oleracea; interação genótipos x ambientes; estabilidade; adaptabilidade; GGE biplot.
GGE biplot analysis in evaluations in bunches characteristics from assai palm trees of the amazon region
Abstract:The interest by assai palms fruits in other regions of the country makes necessary genetic improvement for the Euterpe oleracea. The research aimed to verify, by GGE (Genotype main effects + Genotype-by-environment interaction) biplot, those promising for selection among 30 families from different mother plants of a population of the municipality of Anajás and cultivated in Tomé-Açu, Pará, and also identify the environment that best allows the discrimination of these. The experimental design was in randomized blocks, with 30 treatments (progenies) and three repetitions, were evaluated the characteristics: average weight of the bunches (PMC), in grams; total number of clusters (NTC) and; total weight of the bunches (PTC). It was concluded that for purposes of selection based on the ideotype, the best genotypes in PMC were G22 and G19; in PTC are G10 and G14, and NTC was the genotype G10; the genotypes with average values for the evaluated characteristics showed less stability, except G10 for PTC; The GGE biplots allow you to view the layout, the inter-relationship with environmental effects and choose superior genotypes; in the graphical analysis by GGE biplot method the environments effects (years) allow differentiation of genotypes, with emphasis to the environment A3 for PMC and PTC, and A1 For NTC; for the representativity of all environments is the A1 on the characteristics PMC and PTC; while that is the A2 for NTC. Keywords: Euterpe oleracea; interaction genotypes x environments; stability; adaptability; GGE biplot. .
Keywords: Euterpe oleracea; interaction genotypes x environments; stability; adaptability; GGE biplot.
GGE biplot análisis en la evaluación de las características de los racimos en los açaizeiros de la región amazónica
Resumen:El interés por los frutos de la palma Açaí en otras regiones del país hace necesario para el mejoramiento genético de la Euterpe oleracea. Las investigaciones destinadas a comprobar, por GGE (principales efectos del genotipo y la interacción genotipo x ambiente) biplot, esas prometedoras para la selección entre 30 familias de diferentes plantas madre de una población del municipio de Anajás y cultivadas en Tomé-Açu, Pará, y también identificar el médio ambiente que mejor permite la discriminación de éstas. El diseño experimental fue en bloques al azar, con 30 tratamientos (progenies) y tres repeticiones. Se evaluaron las siguientes características: peso promedio de los racimos (PMC), peso total en gramos de racimos (PTC), en gramos y; el número total de clusters (NTC). Se llegó a la conclusión de que, para los fines de la selección basada en el ideotype, los mejores genotipos en PMC fueron G22 y G19; en el PTC son G10 y G14, y el NTC fue el genotipo G10; los genotipos con valores de promedio más altos para las características mostraron una menor estabilidad, excepto G10 para PTC; excepto G10 para PTC; el GGE Biplots permiten ver el diseño, la inter-relación con los efectos ambientales y seleccionar genotipos superiores; en la análisis gráfica por método GGE biplot los efectos de entornos (años) permitir la diferenciación de genotipos, con énfasis en el medio ambiente A3 para PMC y PTC y A1 Para NTC; a la representatividad de todos los ambientes es el A1 sobre las características PMC y PTC; mientras que el A2 para el NTC.
Palabras clave: Euterpe oleracea; la interacción genotipo x ambiente; estabilidad; adaptabilidad; GGE biplot.
O açaizeiro, (Euterpe oleracea) além de produzir frutos, também é importante fornecedor de palmitos, presente em quase toda a região amazônica, onde a exploração é praticamente baseada na forma extrativa (Homma 2012). O Estado do Pará representa o principal produtor e consumidor de seus produtos, podendo-se identificar as mais densas e diferenciadas populações naturais em áreas de várzeas. Na última estimativa publicada pelo IBGE (2013), a açaizeiro teve uma produção em termos nacionais de 202 mil toneladas de frutos, atingindo uma movimentação comercial de R$ 410 milhões. Convém destacar que 93,7% da produção brasileira de açaí foram oriundos da região Norte, sendo que o Estado do Pará contribuiu com 54,9% dessa produção.
Apesar do aumento anual da produção extrativa, a quantidade ainda é insuficiente para atender a demanda crescente de consumo, estimulando o desenvolvimento do uso de sistemas de irrigação artificial para permitir o cultivo do açaizeiro em terra firme (Farias Neto et al. 2011). A adoção desse sistema de plantio baseia-se na possibilidade de se introduzir o cultivo mecanizado, desde o manejo de insetos e doenças até o momento da colheita, aumentando a produtividade. Conforme Dimenstein e Farias Neto (2003), também há outra vantagem: a resolução do problema em termos da sazonalidade na região da safra, não havendo frutos entre janeiro e junho, permitindo com isso que os trabalhadores tenham opção de trabalho na cadeia produtiva do açaizeiro. No extrativismo estariam desempregados ou sem opção de renda.
Trabalhos publicados por Farias Neto et al. (2003) e Ohashi e Kageyama (2004) demonstram que há diferenciação em termos genéticos entre populações e/ou progênies de açaizeiros. Esse aspecto fornece a possibilidade de seleção de materiais superiores para as características de interesse. Contudo as informações são poucas em relação ao comportamento de indivíduos de diferentes populações cultivados na mesma localidade, eliminando-se as diferenças de ambiente que podem gerar uma resposta diferencial e frequente. Denomina-se de interação genótipo por ambiente (GxE) a esse comportamento, causa problemas e torna mais difícil a recomendação de materiais genéticos mais adaptados e com maior estabilidade por gerar uma diminuição da previsibilidade do comportamento perante as novas condições (Camargo-Buitrago et al. 2011; Gauch 2013).
No melhoramento de plantas, os efeitos ambientais geram a necessidade de avaliar a adaptabilidade e estabilidade genotípica e a predição em gerações posteriores. A presença da GxE complica esse processo e normalmente é expresso como respostas inconsistentes de alguns genótipos em relação a outros, devido à mudança de classificação genotípica, ou como mudanças nas diferenças absolutas entre genótipos, mas com a manutenção da sequência dos mesmos (Araújo et al. 2012; Gauch 2013).
Para as pesquisas de adaptabilidade e também de estabilidade, dentre as metodologias mais recentes, destaca-se o modelo GGE biplot, proposto por Yan et al. (2000). Nesse modelo, o cálculo pondera conjuntamente o efeito principal de genótipo com a da interação genótipo e ambiente. Recomenda-se o seu uso para se discernir os distintos mega-ambientes, selecionando os ambientes que mais perfeitamente representam e discriminam os demais ambientes e, adicionalmente, indicam quais os genótipos mais adaptados e estáveis a ambientes intrínsecos (Gauch et al. 2008; Yan 2011). A metodologia permite a realização de inferências quanto a performance dos materiais genéticos e ambientes, com clareza na discriminação do que está sendo investigado (Yan et al. 2000; Yan 2002; Hassanpanah 2010).
Com base nestas informações, o objetivo deste trabalho foi verificar se há presença de materiais genéticos promissores para seleção entre as trinta progênies originadas de diferentes plantas matrizes de uma população do município de Anajás e cultivadas em Tomé-Açu, Pará, e também identificar o(s) ambiente(s) que melhor permite(m) a discriminação destes, utilizando a metodologia do GGE (Genotype main effects + Genotype environment interaction) biplot.
O estudo foi realizado na área experimental da Embrapa Amazônia Oriental em Tomé-Açu, no Nordeste do Estado do Pará (Latitudes de 01° 24 46,14 S e 01° 28 4,11 S; e longitudes 48° 20 4,60 O e 48° 20 31,84 O). Possuindo clima com temperaturas médias altas e com muita umidade, estando inserido no tipo climático intitulado Ami, da classificação de Köppen, existindo na maior parte do ano chuvas intensas e pouco tempo com déficit hídrico. As precipitações situam-se em dois períodos distintos: no primeiro, do mês de dezembro a maio, com índices pluviométricos acima de 150 mm mensais; e o segundo, envolvendo os meses de junho a novembro, com valores abaixo de 100 mm mensais. Anualmente tem-se uma média de aproximadamente 2.300 mm de precipitação pluviométrica. Nessa localidade a umidade relativa do ar possui pequenas variações dentro do ano, situando-se entre 81% e 89% (valores médios anuais). Para a temperatura do ar há também pouca variação dentro do ano, com valores médios de 26 °C, e máximas na faixa de 32 °C a 34 °C, e mínimas dentro de 21 °C e 23 °C (Valente et al. 2014).
As trinta progênies avaliadas são oriundas de sementes de trinta diferentes plantas matrizes, sem qualquer controle de polinização, pré-selecionadas no momento de coleta pelo tamanho do cacho, todas pertencentes a uma população existente no município de Anajás-PA, com produção de açaí dentro do período considerado como entressafra da espécie. Isso quer dizer que sua produção está concentrada no primeiro semestre, enquanto que as consideradas populações normais produzem no segundo semestre do ano. O teste das suas progênies foi instalado na área experimental em março de 2005. O delineamento experimental adotado foi o de blocos casualizados, contendo trinta tratamentos (progênies), repetidos três vezes e dentro de cada repetição houve a presença de cinco plantas. O espaçamento adotado entre as diferentes plantas foi de 6 m x 4 m. Externamente a área experimental foi inserida uma bordadura.
As características mensuradas entre os anos de 2009 a 2012 foram: peso médio dos cachos (pmc), em gramas; peso total de cachos (PTC), em gramas e; número total de cachos (NTC). Para a definição dos diferentes ambientes foram considerados os anos em que os materiais foram avaliados.
A análise de variância foi realizada com base nas médias das parcelas para verificar a presença de diferenças estatísticas significativas entre as progênies para as diferentes características. O procedimento estatístico adotado foi o proposto por Cruz et al. (2012).
O modelo estatístico empregado foi: Yij = m + Gi + Rj + eij; onde se tem que: Yij representa a média do valor fenotípico para a característica Y avaliado no genótipo identificado como i, no corte j; m representa um média geral de todos os valores em avaliação; Gi representa o efeito do i-ésimo materiais genético, considerado como de origem aleatória; Rj representa o efeito da repetição de número j-ésimo; eij representa o valor residual médio associado à observação identificado como Yij, tendo origem aleatória.
Foi empregado a metodologia que é denominada de GGE biplot, que discorre
conjuntamente sobre o efeito do genótipo e dos efeitos da interação entre
genótipos e ambientes (Yan et al. 2000). No arcabouço estatístico, tem-se
como fundamentais apenas o efeito principal de genótipo e da GxE e
considerados sincronicamente. O GGE biplot não desagrega o efeito de
genótipos (G) da interação genótipos com ambientes (GxE), conservando-os
juntos em dois termos multiplicativos, cuja cenário pode ser observado na
equação:

O ambiente foi definido pelos diferentes anos de avaliação, desejando-se verificar se os materiais genéticos em suas interações com os anos apresentaram comportamento diferenciado.
Foi calculada a relação de informação (ir) proposta por Yan e Tinker (2006) para avaliar a adequação do biplot em exibir os padrões de uma tabela de dupla entrada. A interpretação é: um pc (eixo da interação da análise de componentes principais) com ir = 1 contém padrões (associações entre ambientes), um pc 1 não contém padrões, mas pode conter alguma informação independente e, um pc com ir < 1 não contém qualquer padrão ou informação. O biplot de dimensão 2 representa adequadamente os padrões nos dados, se apenas os dois primeiros pc têm um ir =1.
Todos os procedimentos foram realizados com software estatístico Genes (Cruz 2013) e a análise GGE biplot foi realizada com o pacote estatístico GGE biplot gui do programa R (R Core Team, 2016).
Na tabela 1 encontra-se o resumo da análise de variância contendo os quadrados médios e a significância para o teste F para as características avaliadas. Houve a observação de diferenças significativas entre as progênies avaliadas (g) somente para a característica NTC, particularizando a presença de variabilidade genotípica e com isso a viabilidade da aparição de indivíduos superiores. Galate et al. (2014) avaliando 129 matrizes de açaizeiro do Nordeste Paraense não observaram diferenças para esta característica. A ausência de variabilidade pode ter sido ocasionada pela pré-seleção realizada no momento de coleta das sementes que originaram as plantas instaladas na área experimental, uniformizando-as de forma subjetiva, semelhante para PTC ao citado por Galate et al. (2014). Oliveira e Fernandes (2001) observaram resultados contrastantes ao avaliar em açaizeiros pertencentes ao Banco Ativo de Germoplasma da Embrapa Amazônia Oriental em Belém, Estado do Pará, permitindo inferir que diferentes materiais genéticos e locais da mesma espécie respondem de forma distinta.
Tabela 1. Resumo da análise de variância para três características a em trinta progênies de açaizeiros em quatro anos. Anajás-PA.
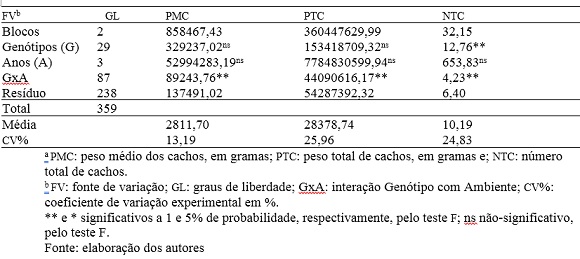
Na fonte de variação anos (a), não foram verificadas diferenças significativas. Pode-se afirmar que na condição experimental oferecida, os fatores não controláveis isoladamente foram incapazes de gerar possíveis diferenças entre as progênies para os valores nas características avaliadas, entre as diferentes safras. Esse comportamento demonstra a presença de estabilidade ambiental, o que é difícil de ocorrer de acordo com Cruz et al. (2012).
Apesar de haver efeito genotípico apenas para a característica NTC e não haver para anos, pode-se observar na tabela 1 que a interação GxA foi significativa em todas as características avaliadas, indicando que os genótipos interagiram de forma diferenciada com as condições ambientais de cada ano. Permitindo a realização da análise GGE biplot.
O coeficiente de variação experimental (cv) pode ser conceituado como de valor médio para pmc e alto para NTC e PTC; neste último grupo os valores demonstram haver elevada variabilidade entre as diferentes progênies, caracterizada pela alta interação com os fatores ambientais e desta forma indicando menor precisão experimental. Essa grande variabilidade verificada pode ser reflexo do sistema de domesticação do açaizeiro encontrar-se apenas em seu prelúdio, sendo a espécie ainda praticamente primitiva, com os programas de melhoramento genético ainda coletando informações morfogenéticas e havendo ainda pouca experimentação com seleção efetiva.
Baseado na relação de informação (ir) dos quatro componentes (tabela 2), para as características pmc e PTC, apenas o primeiro eixo pc conteve padrão (IR1 acima de 1 e IR2 menor que 1). Já para ptf o primeiro eixo conteve padrão (ir acima de 1) e o segundo não (ir próximo de 1), contudo mesmo assim pode conter alguma informação independente. Portanto, nestas características, o biplot foi considerado suficiente para representar os padrões dos dados.
Tabela 2. Valor singular, proporção explicada e relação da informação (ir) dos quatros componentes principais (PC1, PC2, PC3 e PC4) paras as características: peso médio dos cachos (pmc), peso total de cachos (PTC) e número total de cachos (NTC) em progênies de açaizeiro. Anajás-PA.
Os GGE biplot com base nesse conjunto de dados são apresentados nas figuras 1a, 1b e 1c, onde na abscissa do biplot apresentam se as médias para as características e no eixo da ordenada os escores de PC1 e nas figuras 2a, 2b e 2c, na abscissa do biplot são plotados os escores de PC1 e no eixo da ordenada os escores do PC2, dos genótipos e ambientes. O percentual acumulado próximo a 80% nas características estudadas, nos dois primeiros eixos, inferiu elevada confiabilidade na elucidação da variação total do desempenho do genótipo, conjuntamente com a interação com o ambiente (G+GxA), sendo ambiente nestas condições definidas pelos diferentes anos de avaliação. Contudo este valor foi inferior, possivelmente devido ao sistema de cruzamentos eminentemente alógama que permite a ampliação da variabilidade e por não ter havido processo de seleção para a estabilidade e adaptabilidade, diferente em relação as espécies que já passaram por processos de seleção como em pimenteiras, conforme cita Abu et al. (2011) na característica para produção de frutos frescos; em melão por Dehghani et al. (2012) para peso médio por fruto e também com número de frutos por planta; em cajueiros por Aliyu et al. (2014) para o número de amêndoas, produtividade e peso de amêndoas e; em genótipos de mandioca por Peprah et al. (2016) para produtividade de raiz, cujos resultados apresentados no percentual acumulado dos dois primeiros eixos pc valores superiores a 89%.
Figura 1. O GGE biplot (Which-won-where) para os dados das características peso médio dos cachos (a), peso total de cachos (b) e número total de cachos (c) em progênies de açaizeiro, quais dos genótipos apresentaram melhor desempenho em que ambientes. As linhas vermelhas contínuas delimitam setores que definem os mega-ambientes.
Figura 2. O GGE biplot (Média versus Estabilidade) com eixo do ambiente-média (EAM) para mostrar o desempenho médio e estabilidade dos genótipos para as características peso médio dos cachos (a), peso total de cachos (b) e número total de cachos (c) em progênies de açaizeiro, quais dos genótipos apresentaram melhor desempenho em que ambientes.
A figura 1 (which-won-where) dos dados multi ambientes permite o agrupamento dos materiais genéticos e visualizar os superiores e em quais ambientes, tendo importância para averiguar a presumível existência de mega-ambientes diferentes em uma localidade. Os trinta genótipos foram identificados de G1 a G30 e os quatro ambientes, A1, A2, A3 e A4 que seriam os anos 2009, 2010, 2011 e 2012, respectivamente.
Os vértices do polígono para pmc foram os genótipos: G26, G12, G11, G24, G16, G22 e G18. A divisão sugere a formação de dois mega-ambientes formados por (i) contendo os ambientes A1, A3 e A4; (ii) contendo A2. Enquanto que para PTC os vértices do polígono são compostos pelos genótipos: G5, G17, G12, G15, G10, G18 e G26, havendo a formação de dois mega-ambientes, com (i) contendo A1, A2 e A3; (ii) com A4.
Já para NTC os vértices foram formados por: G5, G16, G27, G18, G10, G15 e G17. Os quatros ambientes compuseram dois grupos formados por (i) contendo A1 e A4; (ii) contendo A2 e A3; contudo o ambiente A4 ficou exatamente sobre uma das linhas, podendo-se neste caso distinguir três grupos, se for considerado que A4 pertence ao vértice do genótipo G15, então os grupos seriam (i) contendo A1; (ii) com A2 e A3 e; (iii) com A4, com esta segunda classificação a ser adotada. Havendo maior coincidência entre os genótipos dos vértices entre PTC e NTC.
A divisão em dois mega-ambientes foi semelhante à observada em pimenteiras por três anos por Abu et al. (2011); para produção de frutos de pêssego também em três anos (Citadin et al. 2014); em número de amêndoas por planta, produtividade e peso em cajueiros, observando-se distribuição equitativa de genótipos nestes mega-ambientes, com os vértices (genótipos) do polígono próximos dos ambientes. Com a existência de genótipos que não se enquadraram em nenhum mega-ambiente (Aliyu et al. 2014). Em morango para produtividade já houve um comportamento distinto em que cada local compôs um mega-ambiente (Costa et al. 2016), assim como em mandioca (Peprah et al. 2016).
O genótipo G22 está no vértice do setor em que os ambientes A1, A3 e A4 estão situados, portanto, é o genótipo que teve melhor desempenho nestes ambientes; para o ambiente A2 não houve genótipo que tenha se destacado na característica pmc (figura 1a). Já os genótipos G10 e G18, em PTC, são os vértices do setor em que os ambientes A1, A2 e A3 estão situados, portanto, foram os genótipos que tiveram melhor desempenho nestes ambientes, sendo G10 mais próximo de A1 e A2 e, G18 de A3, para o ambiente A4 o genótipo G15 foi o que teve melhor desempenho (figura 1b). Para NTC o genótipo G18 é o vértice do setor em que os ambientes A2 e A3 estão situados, portanto, é o genótipo que teve melhor desempenho nestes ambientes; para o ambiente A1 foi o genótipo G10 e, para o ambiente A4 foi o genótipo G15 (figura 1c). Os genótipos superiores nos ambientes conforme constatado foram semelhantes em PTC e NTC com destaque para G10 e G18.
Não há proximidade de ambientes nos setores definidos genótipos G26, G12, G11, G24, G16 e G18, significando que estes não foram superiores na característica pmc em nenhum ambiente (figura 1a), já para PTC (figura 1b), os materiais genéticos G5, G17, G12 e G26 representaram baixo desempenho, enquanto que para NTC (figura 1c) foram G5, G16, G27 e G17, significando que estes tiveram desempenho abaixo em relação aos demais materiais genéticos avaliados. Podendo-se observar que G5, G12, G16, G17 e G26 foram inferiores em pelo menos duas das características avaliadas.
Houve a presença de poucos genótipos estáveis, assim como aqueles adaptados especificamente a determinados ambientes ou épocas e, também dos que não possuem estabilidade e nem adaptabilidade. Essas variações são comuns quando se estuda um grupo de genótipos distintos e, também ocorreram em outras espécies como em pimenteira (Abu et al. 2011), cajueiro (Aliyu et al. 2014), pêssego (Citadin et al. 2014), morangueiro (Costa et al. 2016) e mandioca (Peprah et al. 2016).
Denomina-se eixo do ambiente-média ou eam a linha reta contendo uma seta, que seria o ambiente-média (figuras 2a, 2b e 2c). Ao ultrapassar a ponta da seta tem-se um desempenho médio superior entre os genótipos avaliados. A linha perpendicular ao eam, indica que quanto maior a distância da origem maior a variabilidade de desempenho, com presença de menor estabilidade, em ambas as direções (Yan 2002; 2011).
Baseado nos valores médios de PMC os genótipos foram classificados desta forma: G22 > G19 > G18 > G1 > G10 > > G4 > média geral > G25 > G8 > G14> > G12 > G5 > G11. Entre os materiais que se destacaram com desempenho acima da média geral, estão G19 e G22, porém não foram estáveis, mesmo assim tiveram para a característica pmc desempenho de valores médios interessantes em todos os quatro ambientes em relação aos demais genótipos. Os materiais genéticos G6 e G23 tiveram pmc pouco acima da média e foram os mais estáveis; G26 foi o genótipo mais instável de todos os avaliados e pmc semelhante à média geral (figura 2a). Para a característica PTC (figura 2b) a classificação foi: G18 > G10 > G22 > G14 > G15 > G19>... > média geral > > G17 > G28 > G24 > G16 > G25 > G5. Para NTC (figura 2c) seguiu a sequência: G18 > G10 > G11 > G2 > G6 > > média geral > > G26 > G17 > G16 > G25 > G5, sendo que G10 e G18 foram os que mais se destacaram para PTC e NTC, apesar da ausência de estabilidade do segundo genótipo; G2 foi o mais estável para PTC e NTC, contudo teve valor médio semelhante à média geral, seguido por G14 (figura 2b e 2 c). Especificamente pmc e PTC o genótipo G26 e para NTC o genótipo G27 foram os mais instáveis de todos os avaliados (figura 2c).
Um ideótipo é uma forma ideal de planta ou genótipo para determinado ambiente, representando o objetivo almejado, pois apresenta alto desempenho médio e alta estabilidade nos mega-ambientes. A visualização do GGE biplot Média versus Estabilidade é uma ferramenta eficaz para avaliar genótipos em ambos os aspectos (Yan et al. 2007; Yan 2011) e identificar aqueles que possam se aproximar do ideótipo. Baseado nestas informações, nenhum genótipo de açaizeiro superou o ideótipo, pois todos os materiais mais produtivos foram também os mais instáveis, similar ao que foi obtido por Aliyu et al. (2014) quando avaliou cajueiros para número, peso e produtividade de amêndoas. Quanto à dispersão dos materiais genéticos para pmc e PTC, ocorreu a mesma diversidade que aquela apresentada em morangueiro (Costa et al. 2016) e mandioca (Peprah et al. 2016), cujas médias dos genótipos não se concentraram próximo da média dos mega-ambientes, enquanto que em NTC a dispersão foi relativamente menor.
As figuras 3a, 3b e 3c ilustram um conceito importante em relação à estabilidade, o termo estabilidade elevada só tem sentido quando associado ao desempenho médio. Pois um genótipo estável é desejado apenas quando apresenta associado também uma alta performance média para a característica em quaestão (Yan & Tinker 2006; Yan 2011). Nas figuras o ideótipo é representado pelos genótipos que estão sobre o centro dos círculos concêntrico ou próximos no eixo eam, no sentido positivo e tem um comprimento do vetor igual aos vetores mais longos dos genótipos sobre o lado positivo do eam, isto é, mais alto desempenho. Portanto, os genótipos localizados mais próximos do centro dos círculos concêntricos são mais desejáveis do que aqueles que estão mais distantes.
Figura 3. O GGE biplot (com eixo do ambiente-média (EAM) para classificar os genótipos em relação ao ideótipo (no centro dos círculos concêntricos) para as características peso médio dos cachos (a), peso total de cachos (b) e número total de cachos (c) em progênies de açaizeiro, quais dos genótipos apresentaram melhor desempenho em que ambientes.
Assim, G22, para a característica pmc, foi o genótipo mais próximo do ideal neste conjunto de dados, em seguida, os melhores foram os genótipos G19, G1 e G10 (figura 3a), contudo G22 e G19 foram mais desejáveis devido a suas melhores médias para a característica, em relação à G1 e G10, apesar destes apresentarem alta estabilidade. Para PTC foi o genótipo G10, seguido por G14, G18 e G22 (figura 3b), onde G14 e G22 foram mais desejáveis e mais estáveis que o G18 e; para NTC foram os genótipos G2, G10 e G11, seguido pelos genótipos G6 e G18 (figura 3c), sendo mais desejáveis os genótipos G2 e G11, que apesar de apresentarem menor média que G10 e G18, apresentaram maior estabilidade.
Os genótipos com desempenhos inferiores em relação ao grupo avaliado na característica pmc foram G5, G11, G12, G24, G28 e G1; para PTC foram G17, G28, G24, G16, G25 e G5 e; para NTC foram G17, G16, G25 e G5, pode-se notar que existiram materiais genéticos que foram aquém dos demais em mais de uma característica.
Os genótipos que foram altamente estáveis, mas não significando que estes genótipos tiveram valores de pmc superiores, mas apenas que o desempenho relativo destes foram consistentes em termos de estabilidade, mas estão longe de ser um dos genótipos ideais, foram G6 e G23; enquanto que para PTC foram os materiais G2, G11, G13, G22 e G23 e; para NTC foi aquele identificado como G14 (figuras 2 e 3).
O objetivo da avaliação do ambiente de teste é identificar aqueles que podem ser utilizados para selecionar genótipos superiores de forma eficaz num mega-ambiente ou neste caso se algum ambiente (ano) foi capaz de diferenciar de forma mais ampla os materiais genéticos avaliados e permitir a identificação daqueles de melhor desempenho. A visualização no GGE biplot (figuras 4a, 4b e 4c) foi projetada com esta finalidade.
Figura 4. O GGE biplot discriminação e representatividade para mostrar a capacidade de discriminação e representatividade dos ambientes de teste nas características peso médio dos cachos (a), peso total de cachos (b) e número total de cachos (c) em progênies de açaizeiro, quais dos genótipos apresentaram melhor desempenho em que ambientes. Anajás, PA.
Os ambientes de teste que possuem vetores mais longos são mais discriminantes em relação aos genótipos. Aqueles ambientes com vetores curtos são menos discriminantes, o que significa que todos os genótipos se tornam semelhantes, portanto, não devem ser utilizados como ambientes de teste (Hongyu et al. 2015).
Na característica pmc, os ambientes A2, A3 e A4, excetuando-se A1, apresentaram vetores longos, significando que são mais discriminantes em relação aos genótipos (figura 4a), permitindo a manifestação das diferenças entre os materiais genéticos. Já para PTC, foram os ambientes A2 e A3 (figura 4b) em relação a todos os genótipos, enquanto que A1 e A4 foram menos discriminantes apenas em relação ao genótipo G18 e; os ambientes A1, A2 e A3 tiveram maior poder de discriminação, excetuando-se A4, para NTC (figura 4c). Permitindo diferenciar o comportamento dos diferentes materiais genéticos. Semelhantemente, Peprah et al. (2016) observou o mesmo comportamento em produtividade de mandioca, com ambientes possuindo vetores mais longos que a maioria dos genótipos e apenas um material genético com vetor superior ao dos ambientes. Já comparativamente em relação a avaliação para número, peso e produtividade de amêndoas por planta em cajueiro, houve divergências, pois somente os ambientes, com vetores maiores, foram discriminantes, não existindo qualquer genótipo que fosse mais discriminante (Aliyu et al. 2014).
Uma segunda utilidade da figura 4 é indicar ambientes de representatividade do ambiente teste. Esses ambientes seriam os que contém valores dos fatores controláveis e não controláveis na média entre todos os demais ambientes. Os ambientes de teste A1 e A3, na figura 4a, correspondente a característica pmc, que tiveram os menores ângulos com eam (eixo do ambiente-média) são os ambientes mais representativos em relação aos demais. Para PTC (figura 4b) e NTC (figura 4c) foram os ambientes de teste A1 e A2, representando, desta forma, os ambientes de teste que são mais discriminantes e representativos. Esses ambientes podem ser considerados como os mais adequados de teste para a seleção de genótipos adaptados. Os demais ambientes podem ser considerados discriminantes, contudo não são representativos, tendo utilidade para a triagem de genótipos favoráveis especificamente em mega-ambientes; ou para seleção de genótipos instáveis se o ambiente teste é um único mega-ambiente (figura 4a, 4b e 4c). Apoiado neste resultado, pode-se verificar que os principais fatores que compõem os distintos ambientes devem ser também conhecidos, pois compõem condições intrínsecas que servem para a seleção de genótipos. Considerando que os ambientes são compostos por fatores não controláveis e controláveis, ocorrendo responsividade diferenciada pelos genótipos, gerando adaptabilidade e especificidade específicas, então o observado nas figuras 4a, 4b e 4c é perfeitamente normal, semelhante ao que citam Aliyu et al. (2014) trabalhando com diferentes ambientes em cajueiros.
Para cada característica, genótipos distintos se destacaram pela estabilidade ou adaptabilidade associado ao valor médio para a característica avaliada. Contudo os que mais se aproximaram do ideótipo desejado foram para pmc os genótipos G22 e G19; para PTC foram G10 e G14 e, para NTC foi o genótipo G10. Existindo também os que se mostram inferiores em qualquer dos ambientes específicos;
Os genótipos com maiores valores médios para as características mostram em geral menor estabilidade, excetuando-se apenas G10 para PTC, onde o desejável é a combinação de ambos os fatores, qual sejam, alta estabilidade com maiores valores nas características;
Os GGE biplot dispõem os genótipos de forma que é possível se realizar a escolha dos superiores e mostram o inter-relacionamento destes com os efeitos ambientais, devendo buscar a combinação daqueles com melhores valores médios e com maior estabilidade possível;
A análise gráfica do método GGE biplot permite verificar que existe diferenciação entre os ambientes, cujas condições interagem com os materiais genéticos, definindo o ambiente A3 como aquele que melhor oportuniza a distinção de desempenho dos genótipos para pmc e PTC, enquanto que A1 foi para NTC. Para a representatividade de todos os ambientes, é o A1 nas características pmc e PTC; enquanto que é o A2 para NTC.
Abu, N.E., Uguru, M. I. & Obi, I.U. (2011). Genotype by trait relations of yield and yield components in aromatic peppers (Capsicum annuum ) based on gt biplot. Journal of Plant Breeding and Crop Science 3(14): 382-390.
Aliyu, O.M., Adeigbe, O.O. & Lawal, O.O. (2014). Phenotypic stability analysis of yield components in Cashew (Anacardium occidentale L.) using additive main effect and multiplicative interaction (ammi) and gge biplot analyses. Plant Breeding Biotechnology 2(4): 354-369. https://doi.org/10.9787/PBB.2014.2.4.354
Araújo, M.F.C., Araújo, L.B., Faria, P.N. & Dias, C.T.S. (2012). Statistical test for genotype and environment contribution in the genotYpe and environment contribution in the genotypes × environments interaction matrix. Interciencia, 37(7): 515-519.
Camargo-Buitrago, I., Intire, E.Q.M. & Gordón-Mendoza, R. (2011). Identificación de mega-ambientes para potenciar el uso de genótipos superiores de arroz em Panamá. Pesquisa Agropecuária Brasileira 46(9): 1001-1069. https://doi.org/10.1590/S0100-204X2011000900013
Citadin, I., Scariotto, S., Sachet, M.R., Rosa, F.J., Raseira, M.C.B. & Wagner Júnior, A. (2014). Adaptability and stability of fruit set and production of peach trees in a subtropical climate. Scientia Agricola 71(2): 133-138. https://doi.org/10.1590/S0103-90162014000200007
Costa, A.F., Teodoro, P.E., Bhering, L.L., Leal, N.R., Tardin, F.D. & Daher, R.F. (2016). biplot analysis of strawberry genotypes recommended for the State of Espírito Santo. Genetics and Molecular Research 15(3): 1-9. https://doi.org/10.4238/gmr.15038919
Cruz, C.D., Regazzi, A.J. & Carneiro, P.C.S. (2012). Modelos biométricos aplicados ao melhoramento genético. Viçosa: ufv.
Cruz, C.D. (2013). GENES - a software package for analysis in experimental statistics and quantitative genetics. Acta Scientiarum 35(3): 271-276.
Dehghani, H., Feyzian, E., Jalali, M., Rezai, A. & Dane, F. (2012). Use of gge biplot methodology for genetic analysis of yield and related traits in melon (Cucumis melo L.). Canadian Journal of Plant Science 92(1): 77-85. https://doi.org/10.4141/cjps2010-046
Dimenstein, L. & Farias Neto, J.T. (2008). Dados preliminares para a produção de frutos em açaizeiros sob irrigação em terra firme no Estado do Pará. Em: L. Dimestein e J.T. Farias Neto. Irrigação e fertirrigação em fruteiras (pp. 139-144). Fortaleza: Instituto Frutal.
Farias Neto, J.T., Resende, M.D.V. & Oliveira, M.S.P. (2011). Seleção simultânea em progênies de açaizeiro irrigado para produção e peso do fruto. Revista Brasileira de Fruticultura 33(2): 532-539. https://doi.org/10.1590/S0100-29452011000200025
Farias Neto, J.T., Müller, A.A., Oliveira, M.S.P., Espírito Santo, D.E. & Silva, M.A. (2003). Variabilidade genética entre duas procedências de açaizeiro (Euterpe oleracea Martus). Boletim de Pesquisa Florestal 46: 97-104.
Galate, R.S., Mota, M.G.C., Gaia, J.M.D. & Costa, M. Do S.S. (2014). Distância fenotípica entre matrizes de açaizeiro (Euterpe oleracea Mart.) procedentes do nordeste do Pará. Semina: Ciências Agrárias 35(4): 1667-1682. https://doi.org/10.5433/1679-0359.2014v35n4p1667
Gauch, H.G. (2013). A Simple Protocol for ammi Analysis of Yield Trials. Crop Science 53(5): 1860-1869. https://doi.org/10.2135/cropsci2013.04.0241
Gauch, H.G., Piepho, H.P. & Annicchiarico, P. (2008). Statistical analysis of yield trials by ammi and GGE: Further considerations. Crop Science 48(3): 866-889. https://doi.org/10.2135/cropsci2007.09.0513
Hassanpanah, D. (2010). Analysis of GxE interaction by using the additive main effects and multiplicative interaction in potato cultivars. International Journal of Plant Breeding and Genetics 4: 23-29. https://doi.org/10.3923/ijpbg.2010.23.29
Homma, A.K.O. (2012). Extrativismo vegetal ou plantio: qual a opção para a Amazônia?. Estudos Avançados 26(74): 167-186. https://doi.org/10.1590/S0103-40142012000100012
Hongyu, K., Silva, F.L., Oliveira, A.C.S., Sarti, D.A., Araujo, L.B. & Dias, C.T.S. (2015). Comparação entre os modelos ammi e gge biplot para os dados de ensaios multi-ambientais. Revista Brasileira de Biomassa 33(2): 139-155.
IBGE Instituto Brasileiro de Geografia e Estatísticas (2013). Produção da Extração Vegetal e da Silvicultura 2013. Rio de Janeiro: IBGE.
Ohashi, S.T. & Kageyama, P.Y. (2004). Variabilidade genética entre populações de açaizeiro (Euterpe oleracea Mart.) do estuário amazônico. Em: L. Mourão, M.A. Jardim MA e M. Grossmann M (Eds.). Açaí: possibilidade e limites em processos de desenvolvimento sustentável no estuário amazônico (pp. 11-26). Belém: Cejup.
Oliveira, M.S.P. & Fernandes, G.L.C. (2001). Repetibilidade de caracteres do cacho de açaizeiro nas condições de Belém, PA. Revista Brasileira de Fruticultura 23: 613-616. https://doi.org/10.1590/S0100-29452001000300034
Peprah, B.B., Agyeman, A., Parkes, E., Kwadwo, O., Issac, A.K., Emmanuel, O. & Labuschagne, M.T. (2016). Stability, agronomic performance and genetic variability of 10 cassava genotypes in Ghana. Journal of Plant Breeding and Crop Science 8(9): 157-167. https://doi.org/10.5897/JPBCS2016.0581
R Core Team. (2016). R: A language and environment for statistical computing. Viena: R Foundation for Statistical Computing. http://www.R-project.org/.
Valente, M.A., Watrin, O. dos S. & Castro, A.R. da C. (2014). Mapeamento Detalhado dos Solos da Fazenda Experimental da Embrapa Amazônia Oriental em Tomé-Açu, PA . Belém: Embrapa Amazônia Oriental. Embrapa Amazônia Oriental. Documentos, 405.
Yan, W. (2002). Singular-value partition for biplot analysis of multi-environment trial data. Agronomy Journal 94(5): 990-996. https://doi.org/10.2134/agronj2002.0990
Yan, W. & Kang, M.S. (2003). gge biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists . Florida: crc Press.
Yan, W. (2011). gge biplot vs. ammi Graphs for Genotype-by-Environment Data Analysis. Journal of the India Society of Agricultural Statistics 65(2): 181-193.
Yan, W., Hunt, L.A., Sheng, Q. & Szlavnics, Z. (2000). Cultivar evaluation and megaenvironment investigation based on gge biplot. Crop Science 40(3): 597-605. https://doi.org/10.2135/cropsci2000.403597x
Yan, W., Kang, M.S., MA, B., Woods, S. & Cornelius, P.L. (2007). gge biplot vs. ammi analysis of genotype-by-environment data. Crop Science 47(2): 643-653. https://doi.org/10.2135/cropsci2006.06.0374
Yan, W. & Tinker, A. (2006). biplot analysis of multi environment trial data: principles and applications. Canadian Journal of Plant Science 86(3): 623-645. https://doi.org/10.4141/P05-169
Referências
Abu, N.E., Uguru, M. I. & Obi, I.U. (2011). Genotype by trait relations of yield and yield components in aromatic peppers (Capsicum annuum) based on GT biplot. Journal of Plant Breeding and Crop Science 3(14): 382-390.
Aliyu, O.M., Adeigbe, O.O. & Lawal, O.O. (2014). Phenotypic stability analysis of yield components in Cashew (Anacardium occidentale L.) using additive main effect and multiplicative interaction (AMMI) and GGE biplot analyses. Plant Breeding Biotechnology 2(4): 354-369. https://doi.org/10.9787/PBB.2014.2.4.354
Araújo, M.F.C., Araújo, L.B., Faria, P.N. & Dias, C.T.S. (2012). Statistical test for genotype and environment contribution in the genotYpe and environment contribution in the genotypes × environments interaction matrix. Interciencia, 37(7): 515-519.
Camargo-Buitrago, I., Intire, E.Q.M. & Gordón-Mendoza, R. (2011). Identificación de mega-ambientes para potenciar el uso de genótipos superiores de arroz em Panamá. Pesquisa Agropecuária Brasileira 46(9): 1001-1069. https://doi.org/10.1590/S0100-204X2011000900013
Citadin, I., Scariotto, S., Sachet, M.R., Rosa, F.J., Raseira, M.C.B. & Wagner Júnior, A. (2014). Adaptability and stability of fruit set and production of peach trees in a subtropical climate. Scientia Agricola 71(2): 133-138. https://doi.org/10.1590/S0103-90162014000200007
Costa, A.F., Teodoro, P.E., Bhering, L.L., Leal, N.R., Tardin, F.D. & Daher, R.F. (2016). biplot analysis of strawberry genotypes recommended for the State of Espírito Santo. Genetics and Molecular Research 15(3): 1-9. https://doi.org/10.4238/gmr.15038919
Cruz, C.D., Regazzi, A.J. & Carneiro, P.C.S. (2012). Modelos biométricos aplicados ao melhoramento genético. Viçosa: UFV.
Cruz, C.D. (2013). GENES - a software package for analysis in experimental statistics and quantitative genetics. Acta Scientiarum 35(3): 271-276.
Dehghani, H., Feyzian, E., Jalali, M., Rezai, A. & Dane, F. (2012). Use of GGE biplot methodology for genetic analysis of yield and related traits in melon (Cucumis melo L.). Canadian Journal of Plant Science 92(1): 77-85. https://doi.org/10.4141/cjps2010-046
Dimenstein, L. & Farias Neto, J.T. (2008). Dados preliminares para a produção de frutos em açaizeiros sob irrigação em terra firme no Estado do Pará. Em: L. Dimestein e J.T. Farias Neto. Irrigação e fertirrigação em fruteiras (pp. 139-144). Fortaleza: Instituto Frutal.
Farias Neto, J.T., Resende, M.D.V. & Oliveira, M.S.P. (2011). Seleção simultânea em progênies de açaizeiro irrigado para produção e peso do fruto. Revista Brasileira de Fruticultura 33(2): 532-539. https://doi.org/10.1590/S0100-29452011000200025
Farias Neto, J.T., Müller, A.A., Oliveira, M.S.P., Espírito Santo, D.E. & Silva, M.A. (2003). Variabilidade genética entre duas procedências de açaizeiro (Euterpe oleracea Martus). Boletim de Pesquisa Florestal 46: 97-104.
Galate, R.S., Mota, M.G.C., Gaia, J.M.D. & Costa, M. Do S.S. (2014). Distância fenotípica entre matrizes de açaizeiro (Euterpe oleracea Mart.) procedentes do nordeste do Pará. Semina: Ciências Agrárias 35(4): 1667-1682. https://doi.org/10.5433/1679-0359.2014v35n4p1667
Gauch, H.G. (2013). A Simple Protocol for AMMI Analysis of Yield Trials. Crop Science 53(5): 1860-1869. https://doi.org/10.2135/cropsci2013.04.0241
Gauch, H.G., Piepho, H.P. & Annicchiarico, P. (2008). Statistical analysis of yield trials by AMMI and GGE: Further considerations. Crop Science 48(3): 866-889. https://doi.org/10.2135/cropsci2007.09.0513
Hassanpanah, D. (2010). Analysis of GxE interaction by using the additive main effects and multiplicative interaction in potato cultivars. International Journal of Plant Breeding and Genetics 4: 23-29. https://doi.org/10.3923/ijpbg.2010.23.29
Homma, A.K.O. (2012). Extrativismo vegetal ou plantio: qual a opção para a Amazônia?. Estudos Avançados 26(74): 167-186. https://doi.org/10.1590/S0103-40142012000100012
Hongyu, K., Silva, F.L., Oliveira, A.C.S., Sarti, D.A., Araujo, L.B. & Dias, C.T.S. (2015). Comparação entre os modelos AMMI e GGE biplot para os dados de ensaios multi-ambientais. Revista Brasileira de Biomassa 33(2): 139-155.
IBGE – Instituto Brasileiro de Geografia e Estatísticas (2013). Produção da Extração Vegetal e da Silvicultura 2013. Rio de Janeiro: IBGE.
Ohashi, S.T. & Kageyama, P.Y. (2004). Variabilidade genética entre populações de açaizeiro (Euterpe oleracea Mart.) do estuário amazônico. Em: L. Mourão, M.A. Jardim MA e M. Grossmann M (Eds.). Açaí: possibilidade e limites em processos de desenvolvimento sustentável no estuário amazônico (pp. 11-26). Belém: Cejup.
Oliveira, M.S.P. & Fernandes, G.L.C. (2001). Repetibilidade de caracteres do cacho de açaizeiro nas condições de Belém, PA. Revista Brasileira de Fruticultura 23: 613-616. https://doi.org/10.1590/S0100-29452001000300034
Peprah, B.B., Agyeman, A., Parkes, E., Kwadwo, O., Issac, A.K., Emmanuel, O. & Labuschagne, M.T. (2016). Stability, agronomic performance and genetic variability of 10 cassava genotypes in Ghana. Journal of Plant Breeding and Crop Science 8(9): 157-167. https://doi.org/10.5897/JPBCS2016.0581
R Core Team. (2016). R: A language and environment for statistical computing. Viena: R Foundation for Statistical Computing. http://www.R-project.org/.
Valente, M.A., Watrin, O. dos S. & Castro, A.R. da C. (2014). Mapeamento Detalhado dos Solos da Fazenda Experimental da Embrapa Amazônia Oriental em Tomé-Açu, PA. Belém: Embrapa Amazônia Oriental. Embrapa Amazônia Oriental. Documentos, 405.
Yan, W. (2002). Singular-value partition for biplot analysis of multi-environment trial data. Agronomy Journal 94(5): 990-996. https://doi.org/10.2134/agronj2002.0990
Yan, W. & Kang, M.S. (2003). GGE biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists. Florida: CRC Press.
Yan, W. (2011). GGE biplot vs. AMMI Graphs for Genotype-by-Environment Data Analysis. Journal of the India Society of Agricultural Statistics 65(2): 181-193.
Yan, W., Hunt, L.A., Sheng, Q. & Szlavnics, Z. (2000). Cultivar evaluation and megaenvironment investigation based on GGE biplot. Crop Science 40(3): 597-605. https://doi.org/10.2135/cropsci2000.403597x
Yan, W., Kang, M.S., MA, B., Woods, S. & Cornelius, P.L. (2007). GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Science 47(2): 643-653. https://doi.org/10.2135/cropsci2006.06.0374
Yan, W. & Tinker, A. (2006). biplot analysis of multi environment trial data: principles and applications. Canadian Journal of Plant Science 86(3): 623-645. https://doi.org/10.4141/P05-169
Como Citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Baixar Citação
CrossRef Cited-by
1. Jocarla Ambrosim Crevelari, Yure Pequeno de Souza, Juliana Saltires Santos, Moisés Ambrósio, Vivane Mirian Lanhellas Gonçalves, Messias Gonzaga Pereira. (2023). Adaptability and stability of corn hybrids for silage via genotype and genotype × environment interaction biplot. Agronomy Journal, 115(2), p.687. https://doi.org/10.1002/agj2.21240.
Dimensions
PlumX
Acessos à página de resumo
Downloads
Licença
Copyright (c) 2017 Mundo Amazónico

Este trabalho está licenciado sob uma licença Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.
Os autores são responsáveis por todas as autorizações que a publicação das contribuições podem exigir. Quando o manuscrito é aceito para publicação, os autores devem enviar uma declaração formal sobre a autenticidade do trabalho, assumindo a responsabilidade pessoal por tudo o que o artigo contém e expressando seu direito de editá-lo. A publicação de um artigo no Mundo Amazónico não implica a transferência de direitos pelos seus autores. No entanto, a apresentação da contribuição representa a autorização dos autores para a Mundo Amazónico para publicação. No caso de uma reimpressão total ou parcial de um artigo publicado no Mundo Amazónico, em seu idioma original ou em uma versão traduzida, a fonte original deve ser citada. Os artigos publicados na revista são cobertos por um Creative Commons 4.0.Os autores que publicam nesta revista concordam com os seguintes termos:
- Os autores mantêm os direitos autorais e concedem à revista o direito da primeira publicação, com o trabalho licenciado simultaneamente sob uma licença Creative Commons Attribution License que permite que outros compartilhem o trabalho com o reconhecimento da autoria e a publicação inicial nesta revista.
- Os autores podem fazer arranjos contratuais adicionais para a distribuição não exclusiva da versão publicada na revista (por exemplo, colocá-lo em um repositório institucional ou publicá-lo em um livro), com reconhecimento de sua publicação inicial nesta revista.













